

Evaluating the Bioremediation Capacity of the Polychaete Perinereis gualpensis (Jeldes, 1963) for Atlantic Salmon Aquaculture Sludge

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Perinereis gualpensis
2.2. Salmon Aquaculture Sludge
2.3. Experimental Design
2.4. Organic Compounds Analysis
2.5. Statistical Analysis
3. Results
3.1. Aquaculture Sludge
3.2. Growth and Survival of Perinereis gualpensis
3.3. Organic and Inorganic Components Analysis
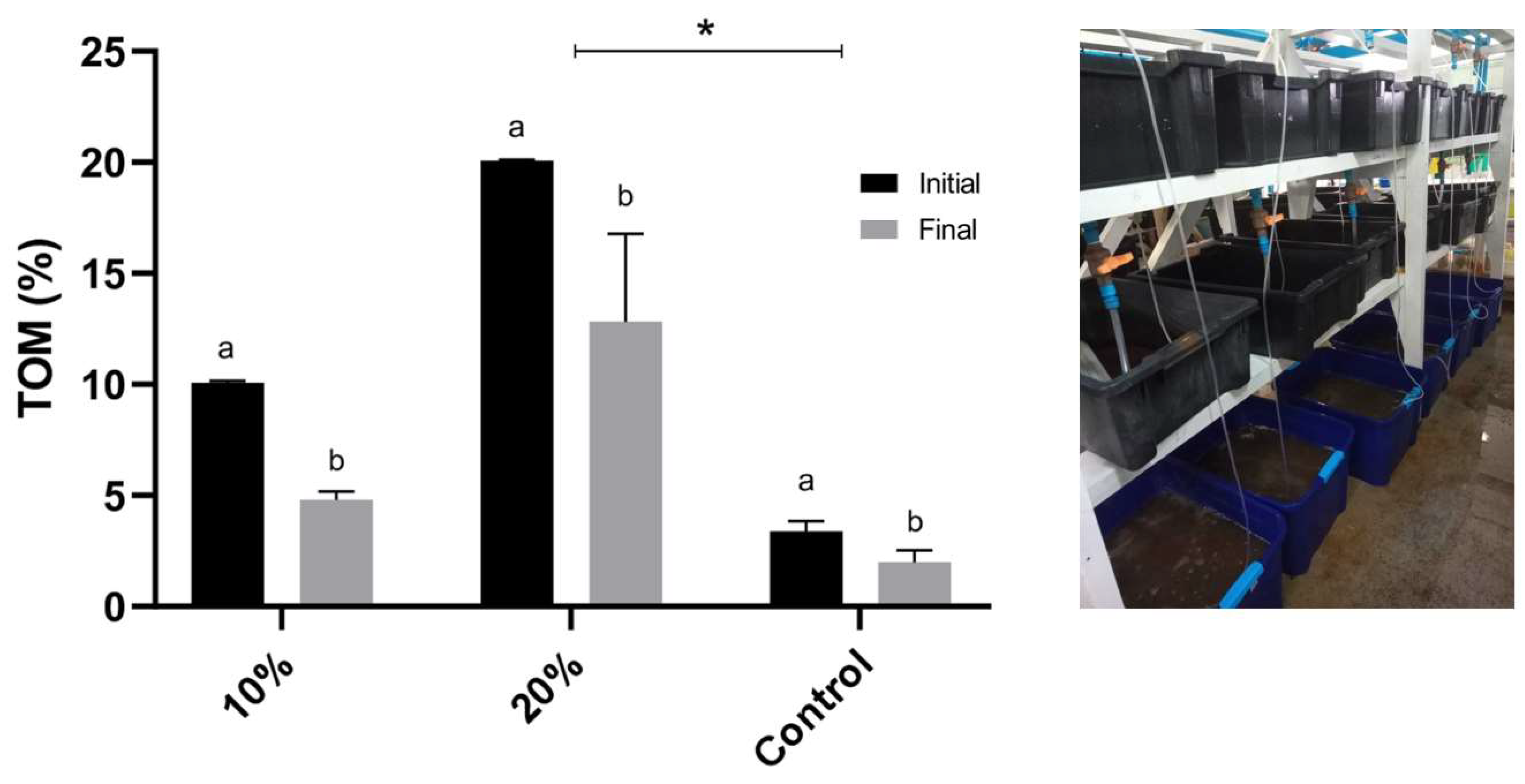
3.3.1. Total Organic Matter
3.3.2. Total Nitrogen (N) and Total Carbon (C)
3.3.3. Protein Content
3.3.4. Water Content
4. Discussion
4.1. TOM Removal Rates
4.2. Total N and Total C
4.3. Protein Content
4.4. Growth and Survival of P. gualpensis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dauda, A.B.; Ajadi, A.; Tola-Fabunmi, A.S.; Akinwole, A.O. Waste production in aquaculture: Sources, components and managements in different culture systems. Aquac. Fish. 2019, 4, 81–88. [Google Scholar] [CrossRef]
- Akizuki, S.; Toda, T. An anaerobic-aerobic sequential batch process with simultaneous methanogenesis and short-cut denitrification for the treatment of marine biofoulings. Waste Manag. 2018, 74, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Galasso, H.L.; Lefebvre, S.; Aliaume, C.; Sadoul, B.; Callier, M.D. Using the dynamic energy budget theory to evaluate the bioremediation potential of the polychaete Hediste diversicolor in an integrated multi-trophic aquaculture system. Ecol. Model. 2020, 437, 109296. [Google Scholar] [CrossRef]
- Lang, Z.; Zhou, M.; Zhang, Q.; Yin, X.; Li, Y. Comprehensive treatment of marine aquaculture wastewater by a cost-effective flow-through electro-oxidation process. Sci. Total Environ. 2020, 722, 137812. [Google Scholar] [CrossRef] [PubMed]
- Senff, P.; Elba, B.; Kunzmann, A.; Gillis, L.G.; Robinson, G. Carbon supplementation promotes assimilation of aquaculture waste by the sea cucumber Holothuria scabra: Evidence from stable isotope analysis. Aquaculture 2022, 547, 737295. [Google Scholar] [CrossRef]
- Maigual-Enriquez, Y.A.; Maia, A.A.D.; Guerrero-Romero, C.L.; Matsumoto, T.; Rangel, E.C.; de Morais, L.C. Comparison of sludges produced from two different recirculating aquaculture systems (RAS) for recycle and disposal. Aquaculture 2019, 502, 87–96. [Google Scholar] [CrossRef]
- Jasmin, M.Y.; Syukri, F.; Kamarudin, M.S.; Karim, M. Potential of bioremediation in treating aquaculture sludge. Aquaculture 2020, 519, 734905. [Google Scholar] [CrossRef]
- Schumann, M.; Brinker, A. Understanding and managing suspended solids in intensive salmonid aquaculture: A review. Rev. Aquacult. 2020, 12, 2109–2139. [Google Scholar] [CrossRef]
- Malzahn, A.M.; Villena-Rodríguez, A.; Monroig, Ó.; Johansen, Å.; Castro, L.F.C.; Navarro, J.C.; Hagemann, A. Diet rather than temperature determines the biochemical composition of the ragworm Hediste diversicolor (OF Müller, 1776) (Annelida: Nereidae). Aquaculture 2023, 569, 739368. [Google Scholar] [CrossRef]
- Jerónimo, D.; Lillebø, A.I.; Cremades, J.; Cartaxana, P.; Calado, R. Recovering wasted nutrients from shrimp farming through the combined culture of polychaetes and halophytes. Sci. Rep. 2021, 11, 6587. [Google Scholar] [CrossRef]
- Yearsley, R.D.; Jones, C.L.; Britz, P.J.; Vine, N.G. Integrated culture of silver kob Argyrosomus inodorus and bloodworm Arenicola loveni loveni in abalone farm effluent. Afr. J. Mar. Sci. 2011, 33, 223–228. [Google Scholar] [CrossRef]
- Marques, B.; Calado, R.; Lillebø, A.I. New species for the biomitigation of a super-intensive marine fish farm effluent: Combined use of polychaete-assisted sand filters and halophyte aquaponics. Sci. Total Environ. 2017, 599, 1922–1928. [Google Scholar] [CrossRef]
- Pajand, Z.O.; Soltani, M.; Bahmani, M.; Kamali, A. The role of polychaete Nereis diversicolor in bioremediation of wastewater and its growth performance and fatty acid composition in an integrated culture system with Huso huso (Linnaeus, 1758). Aquac. Res. 2017, 48, 5271–5279. [Google Scholar] [CrossRef]
- Pombo, A.; Baptista, T.; Granada, L.; Ferreira, S.M.; Gonçalves, S.C.; Anjos, C.; Sá, E.; Chainho, P.; da Fonseca, L.C.; Fidalgo e Costa, P.; et al. Insight into aquaculture’s potential of marine annelid worms and ecological concerns: A review. Rev. Aquacult. 2018, 12, 107–121. [Google Scholar] [CrossRef]
- Wang, H.; Hagemann, A.; Reitan, K.I.; Ejlertsson, J.; Wollan, H.; Handå, A.; Malzahn, A.M. Potential of the polychaete Hediste diversicolor fed on aquaculture and biogas side streams as an aquaculture food source. Aquac. Environ. Interact. 2019, 11, 551–562. [Google Scholar] [CrossRef]
- Dahl, T.H. Biochemical Composition of Hediste diversicolor Cultivated on Aquaculture Sludge and Utilization as a Potential Fish Feed Resource. Master’s Thesis, Norwegian University of Science and Technology, Trondheim, Norway, 2021. Available online: https://hdl.handle.net/11250/2787911 (accessed on 15 May 2023).
- Kristensen, B.S.B. Bioremediation of Aquaculture Sludge by Cultivation of Hediste diversicolor (OF Müller, 1776). Master’s Thesis, Norwegian University of Science and Technology, Trondheim, Norway, 2021. Available online: https://hdl.handle.net/11250/2787912 (accessed on 15 May 2023).
- Anglade, I.; Dahl, T.H.; Kristensen, B.S.; Hagemann, A.; Malzahn, A.M.; Reitan, K.I. Biochemical composition of Hediste diversicolor (OF Müller, 1776) (Annelida: Nereidae) reared on different types of aquaculture sludge. Front. Mar. Sci. 2023, 10, 1197052. [Google Scholar] [CrossRef]
- Wang, H.; Seekamp, I.; Malzahn, A.; Hagemann, A.; Carvajal, A.K.; Slizyte, R.; Standal, I.B.; Handå, A.; Reitan, K.I. Growth and nutritional composition of the polychaete Hediste diversicolor (OF Müller, 1776) cultivated on waste from land-based salmon smolt aquaculture. Aquaculture 2019, 502, 232–241. [Google Scholar] [CrossRef]
- Fang, J.; Zhang, J.; Jiang, Z.; Du, M.; Liu, Y.; Mao, Y.; Gao, Y.; Fang, J. Environmental remediation potential of Perinereis aibuhitensis (Polychaeta) based on the effects of temperature and feed types on its carbon and nitrogen budgets. Mar. Biol. Res. 2016, 12, 583–594. [Google Scholar] [CrossRef]
- Gómez, S.; Hurtado, C.F.; Orellana, J. Bioremediation of organic sludge from a marine recirculating aquaculture system using the polychaete Abarenicola pusilla (Quatrefages, 1866). Aquaculture 2019, 507, 377–384. [Google Scholar] [CrossRef]
- Hu, F.; Sun, M.; Fang, J.; Wang, G.; Li, L.; Gao, F.; Jian, Y.; Wang, X.; Liu, G.; Zou, Y.; et al. Carbon and nitrogen budget in fish-polychaete integrated aquaculture system. J. Oceanol. Limnol. 2021, 39, 1151–1159. [Google Scholar] [CrossRef]
- Palmer, P.J.; Wang, S.; Houlihan, A.; Brock, I. Nutritional status of a nereidid polychaete cultured in sand filters of mariculture wastewater. Aquac. Nutr. 2014, 20, 675–691. [Google Scholar] [CrossRef]
- Monroig, Ó. Biosynthesis of long-chain polyunsaturated fatty acids in aquatic invertebrates: Applications in aquaculture nutrition. In Revisions in Nutrition of Aquatic Organisms, 1st ed.; Cruz-Suárez, L.E., Ricque-Marie, D., Tapia-Salazar, M., Nieto-López, M.G., Villarreal-Cavazos, D.A., Gamboa-Delgado, J., Gaxiola Cortes, M.G., Olvera-Novoa, M., Eds.; Universidad Autónoma de Nuevo León: San Nicolás de los Garza, Nuevo León, Mexico, 2019; pp. 249–270. Available online: http://hdl.handle.net/10261/209414 (accessed on 23 May 2023)ISBN 978-607-27-1268-3.
- Yang, D.; Wang, C.; Kou, N.; Xing, J.; Li, X.; Zhao, H.; Luo, M. Gonadal maturation in Litopenaeus vannamei fed on four different polychaetes. Aquac. Rep. 2022, 22, 100920. [Google Scholar] [CrossRef]
- Estante-Superio, E.G.; Mandario, M.A.E.; Santander-Avanceña, S.S.; Geanga, T.M.M.; Parado-Estepa, F.D.; Mamauag, R.E.P. Inclusion of live mud polychaete (Marphysa iloiloensis) in the feeding regime improved the hatchery performance of domesticated Indian white shrimp (Penaeus indicus). Reg. Stud. Mar. Sci. 2023, 62, 102923. [Google Scholar] [CrossRef]
- Farías, A.; Valenzuela, G.; Hernández, J.; Uriarte, I.; Viana, M.T. Seasonal Variation in Fatty Acid and Amino Acid Composition of the Patagonian Marine Polychaete Abarenicola pusilla and Its By-Products. Aquac. Res. 2023, 2023, 719721. [Google Scholar] [CrossRef]
- Honda, H.; Kikuchi, K. Nitrogen budget of polychaete Perinereis nuntia vallata fed on the feces of Japanese flounder. Fish. Sci. 2002, 68, 1304–1308. [Google Scholar] [CrossRef]
- Bischoff, A.A.; Fink, P.; Waller, U. The fatty acid composition of Nereis diversicolor cultured in an integrated recirculated system: Possible implications for aquaculture. Aquaculture 2009, 296, 271–276. [Google Scholar] [CrossRef]
- Seekamp, I. Utilization of Excess Nutrients from Land-Based Aquaculture Facilities by Hediste diversicolor (OF Müller, 1776). Production of Polychaete Biomass and Its Potential Use in Fish Feed. Master’s Thesis, Norwegian University of Science and Technology, Trondheim, Norway, 2017. Available online: http://hdl.handle.net/11250/2451314 (accessed on 15 May 2023).
- Gómez, S.; Hurtado, C.F.; Orellana, J.; Valenzuela-Olea, G.; Turner, A. Abarenicola pusilla (Quatrefages, 1866): A novel species for fish waste bioremediation from marine recirculating aquaculture systems. Aquac. Res. 2018, 49, 1363–1367. [Google Scholar] [CrossRef]
- Marques, B.; Lillebø, A.I.; Ricardo, F.; Nunes, C.; Coimbra, M.A.; Calado, R. Adding value to ragworms (Hediste diversicolor) through the bioremediation of a super-intensive marine fish farm. Aquac. Environ. Int. 2018, 10, 79–88. [Google Scholar] [CrossRef]
- Jerónimo, D.; Lillebø, A.I.; Santos, A.; Cremades, J.; Calado, R. Performance of polychaete assisted sand filters under contrasting nutrient loads in an integrated multi-trophic aquaculture (IMTA) system. Sci. Rep. 2020, 10, 20871. [Google Scholar] [CrossRef]
- Nesto, N.; Simonini, R.; Prevedelli, D.; Da Ros, L. Effects of diet and density on growth, survival and gametogenesis of Hediste diversicolor (OF Müller, 1776) (Nereididae, Polychaeta). Aquaculture 2012, 362, 1–9. [Google Scholar] [CrossRef]
- Avendaño-Herrera, R. Proper antibiotics use in the Chilean salmon industry: Policy and technology bottlenecks. Aquaculture 2018, 495, 803–805. [Google Scholar] [CrossRef]
- Quiñones, R.A.; Fuentes, M.; Montes, R.M.; Soto, D.; León-Muñoz, J. Environmental issues in Chilean salmon farming: A review. Rev. Aquacult. 2019, 11, 375–402. [Google Scholar] [CrossRef]
- Quijón, P.; Jaramillo, E. Temporal variability in the intertidal macroinfauna in the Queule river estuary, south-central Chile. Estuar. Coast. Shelf Sci. 1993, 37, 655–667. [Google Scholar] [CrossRef]
- Sampértegui, S.; Rozbaczylo, N.; Canales-Aguirre, C.B.; Carrasco, F.; Hernández, C.E.; Rodríguez-Serrano, E. Morphological and molecular characterization of Perinereis gualpensis (Polychaeta: Nereididae) and its phylogenetic relationships with other species of the genus off the Chilean coast, Southeast Pacific. Cah. Biol. Mar. 2013, 54, 27–40. Available online: https://repositorio.uc.cl/xmlui/bitstream/handle/11534/12412/672.pdf (accessed on 17 May 2023).
- Bertrán, C. Zonación y Dinámica Temporal de la Macroinfauna Intermareal en el Estuario del río Lingue (Valdivia, Chile). Rev. Chil. Hist. Nat. 1989, 62, 19–32. Available online: http://rchn.biologiachile.cl/pdfs/1989/1/Beltran_1989.pdf (accessed on 17 May 2023).
- Díaz-Jaramillo, M.; Sandoval, N.; Barra, R.; Gillet, P.; Valdovinos, C. Spatio-temporal population and reproductive responses in Perinereis gualpensis (Polychaeta: Nereididae) from estuaries under different anthropogenic influences. Chem. Ecol. 2015, 31, 308–319. [Google Scholar] [CrossRef]
- Gaete, H.; Álvarez, M.; Lobos, G.; Soto, E.; Jara-Gutiérrez, C. Assessment of oxidative stress and bioaccumulation of the metals Cu, Fe, Zn, Pb, Cd in the polychaete Perinereis gualpensis from estuaries of central Chile. Ecotoxicol. Environ. Saf. 2017, 145, 653–658. [Google Scholar] [CrossRef]
- Jørgensen, E.H.; Christiansen, J.S.; Jobling, M. Effects of stocking density on food intake, growth performance and oxygen consumption in Arctic charr (Salvelinus alpinus). Aquaculture 1993, 110, 191–204. [Google Scholar] [CrossRef]
- Hopkins, K.D. Reporting fish growth: A review of the basics. J. World Aquacult. Soc. 1992, 23, 173–179. [Google Scholar] [CrossRef]
- Berntsen, F.H. Growth of the Polychaete Hediste diversicolor (OF Müller, 1776) Fed on Smolt Sludge and Biogas Residues-Evaluation of RNA/DNA Ratio as Growth Indicator. Master’s Thesis, Norwegian University of Science and Technology, Trondheim, Norway, 2018. Available online: http://hdl.handle.net/11250/2502353 (accessed on 15 June 2023).
- Nederlof, M.A.; Fang, J.; Dahlgren, T.G.; Rastrick, S.P.; Smaal, A.C.; Strand, Ø.; Sveier, H.; Verdegem, M.C.J.; Jansen, H.M. Application of polychaetes in (de) coupled integrated aquaculture: An approach for fish waste bioremediation. Aquac. Environ. Interact. 2020, 12, 385–399. [Google Scholar] [CrossRef]
- Nederlof, M.A.; Jansen, H.M.; Dahlgren, T.G.; Fang, J.; Meier, S.; Strand, Ã.; Sveier, H.; Verdegem, M.C.J.; Smaal, A.C. Application of polychaetes in (de) coupled integrated aquaculture: Production of a high-quality marine resource. Aquac. Environ. Interact. 2019, 11, 221–237. [Google Scholar] [CrossRef]
- Kurihara, Y. Study of domestic sewage waste treatment by the polychaetes, Neanthes japonica and Perinereis nuntia var. vallata, on an artificial tidal flat. Int. Rev. Gesamten. Hydrobiol. 1983, 68, 649–670. [Google Scholar] [CrossRef]
- Yousefi-Garakouei, M.; Kamali, A.; Soltani, M. Effects of rearing density on growth, fatty acid profile and bioremediation ability of polychaete Nereis diversicolor in an integrated aquaculture system with rainbow trout (Oncorhynchus mykiss). Aquac. Res. 2019, 50, 725–735. [Google Scholar] [CrossRef]
- Jerónimo, D.; Lillebø, A.I.; Rey, F.; Ii, H.K.; Domingues, M.R.M.; Calado, R. Optimizing the Timeframe to Produce Polychaetes (Hediste diversicolor) Enriched with Essential Fatty Acids Under Different Combinations of Temperature and Salinity. Front. Mar. Sci. 2021, 8, 671545. [Google Scholar] [CrossRef]
- Bischoff, A.A. Solid Waste Reduction of Closed Recirculated Aquaculture Systems by Secondary Culture of Detritivorous Organisms. Ph.D. Thesis, Leibniz-Institute of Marine Sciences, Kiel, Germany, 2007. Available online: https://macau.uni-kiel.de/servlets/MCRFileNodeServlet/dissertation_derivate_00002061/d2061.pdf (accessed on 15 May 2023).
- Mandario, M.A.E.; Alava, V.R.; Añasco, N.C. Evaluation of the bioremediation potential of mud polychaete Marphysa sp. in aquaculture pond sediments. Environ. Sci. Pollut. Res. 2019, 26, 29810–29821. [Google Scholar] [CrossRef]
- Bianchi, T.S. Biogeochemistry of Estuaries; Oxford University Press: New York, NY, USA, 2007. [Google Scholar]
- Andrade, C.; Ríos, C.; Gerdes, D.; Brey, T. Trophic structure of shallow-water benthic communities in the sub-Antarctic Strait of Magellan. Polar Biol. 2016, 39, 2281–2297. [Google Scholar] [CrossRef]
- Nelson, E.J.; MacDonald, B.A.; Robinson, S.M.C. The absorption efficiency of the suspension-feeding sea cucumber, Cucumaria frondosa, and its potential as an extractive integrated multi-trophic aquaculture (IMTA) species. Aquaculture 2012, 370, 19–25. [Google Scholar] [CrossRef]
- Mongirdas, V.; Žibienė, G.; Žibas, A. Waste and its characterization in closed recirculating aquaculture systems: A review. Water Secur. 2017, 3, 1–8. [Google Scholar] [CrossRef][Green Version]
- Palmer, P.J. Polychaete-assisted sand filters. Aquaculture 2010, 306, 369–377. [Google Scholar] [CrossRef]
- Brown, N.; Eddy, S.; Plaud, S. Utilization of waste from a marine recirculating fish culture system as a feed source for the polychaete worm, Nereis virens. Aquaculture 2011, 322, 177–183. [Google Scholar] [CrossRef]
- Magaña-Gallegos, E.; González-Zúñiga, R.; Arevalo, M.; Cuzon, G.; Chan-Vivas, E.; López-Aguiar, K.; Noreña-Barroso, E.; Pacheco, E.; Valenzuela, M.; Maldonado, C.; et al. Biofloc and food contribution to grow-out and broodstock of Farfantepenaeus brasiliensis (Latreille, 1817) determined by stable isotopes and fatty acids. Aquac. Res. 2018, 49, 1782–1794. [Google Scholar] [CrossRef]
- Ortiz, P.; Quiroga, E.; Montero, P.; Hamame, M.; Betti, F. Trophic structure of benthic communities in a Chilean fjord (45 °S) influenced by salmon aquaculture: Insights from stable isotopic signatures. Mar. Pollut. Bull. 2021, 173, 113149. [Google Scholar] [CrossRef] [PubMed]
- Seemann, J. The use of 13C and 15N isotope labeling techniques to assess heterotrophy of corals. J. Exp. Mar. Biol. Ecol. 2013, 442, 88–95. [Google Scholar] [CrossRef]
- Pairohakul, S.; Olive, P.J.; Bentley, M.G.; Caldwell, G.S. Trophic upgrading of long-chain polyunsaturated fatty acids by polychaetes: A stable isotope approach using Alitta virens. Mar. Biol. 2021, 168, 67. [Google Scholar] [CrossRef]
- Yokoyama, H. Growth and food source of the sea cucumber Apostichopus japonicus cultured below fish cages—Potential for integrated multi-trophic aquaculture. Aquaculture 2013, 372, 28–38. [Google Scholar] [CrossRef]
- Olive, P.J.W.; Craig, S.; Cowin, P.B.D. Aquaculture of Marine Worms. U.S. Patent 7004109 B2, 28 February 2006. Available online: https://patents.google.com/patent/US7004109B2/en (accessed on 6 June 2023).
- Meziane, T.; Retiere, C. Growth of Nereis diversicolor (L.) juveniles fed with detritus of halophytes. Oceanol. Acta 2002, 25, 119–124. [Google Scholar] [CrossRef]
- Suckow, B.; Bischoff, A.; Buck, B.H.; Simon, M. Growth and Biochemical Composition of Nereis virens (Sars, 1835) Applied as a Bioconverter of Solid Waste from Land-Based Fish Culture. Seafarming Tomorrow. In Proceedings of the Aquaculture Europe 2010—The Annual Meeting of the European Aquaculture Society, Porto, Portugal, 5–8 October 2010; Available online: https://epic.awi.de/id/eprint/33982/ (accessed on 23 May 2023).
- García-Alonso, J.; Müller, C.T.; Hardege, J.D. Influence of food regimes and seasonality on fatty acid composition in the ragworm. Aquat. Biol. 2008, 4, 7–13. [Google Scholar] [CrossRef]
- Yang, D.; Cao, C.; Wang, G.; Zhou, Y.; Xiu, Z. The Growth Study of in Airlift Recirculating Aquaculture System. Open Biotechnol. J. 2015, 9, 143–149. [Google Scholar] [CrossRef]
- Batista, F.M.; Fidalgo e Costa, P.; Matias, D.; Joaquim, S.; Massapina, C.; Passos, A.M.; Ferreira, P.P.; da Fonseca, L.C. Preliminary Results on the Growth and Survival of the Polychaete Nereis diversicolor (OF Muller, 1776), When Fed with Faeces from the Carpet Shell Clam Ruditapes decussatus (L., 1758). Bolet. Inst. Esp. Oceanogr. 2003, 19, 443–446. Available online: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.556.4108&rep=rep1&type=pdf (accessed on 23 May 2023).
- Serebiah, J.S. Culture of marine polychaetes. In Advances in Marine and Brackishwater Aquaculture; Perumal, S., Thirunavukkarasu, A.R., Pachiappan, P., Eds.; Springer: New Delhi, India, 2015; pp. 43–49. [Google Scholar] [CrossRef]
- Fauchald, K.; Jumars, P.A. The Diet of Worms: A Study of Polychaete Feeding Guilds. Oceanogr. Mar. Biol. Ann. Rev. 1979, 17, 193–284. Available online: https://repository.si.edu/bitstream/handle/10088/3422/OMBARFauchald1979.pdf (accessed on 6 June 2023).
- Galasso, H.L.; Richard, M.; Lefebvre, S.; Aliaume, C.; Callier, M.D. Body size and temperature effects on standard metabolic rate for determining metabolic scope for activity of the polychaete Hediste (Nereis) diversicolor. PeerJ 2018, 6, e5675. [Google Scholar] [CrossRef]
- Fang, J.; Jiang, Z.; Fang, J.; Kang, B.; Gao, Y.; Du, M. Selectivity of Perinereis aibuhitensis (Polychaeta, Nereididae) feeding on sediment. Mar. Biol. Res. 2018, 14, 478–483. [Google Scholar] [CrossRef]


{kind=link}
| Water content (%) | 95.67 ± 0.74 |
| Dry matter (mg/g WW) | 43.31 ± 7.39 |
| Total organic matter (mg/g DM) | 845.84 ± 14.73 |
| T (°C) | 16.77 ± 0.06 |
| Salinity (g L−1) | 0.00 ± 0.00 |
| pH | 5.98 ± 0.02 |
| Ash content (%) | 15.42 ± 1.47 |
| N total (%) | 3.20 ± 0.23 |
| C total (%) | 25.41 ± 1.71 |
| C/N ratio | 7.95 ± 0.05 |
| Protein content (g/100 g) | 17.06 ± 0.13 |
| 10% Sludge | 20% Sludge | Control | |
|---|---|---|---|
| Absolute growth rate (g m−2) | 0.03 ± 0.01 a | 0.11 ± 0.04 a | −0.42 ± 0.05 b |
| Specific growth rate (µ) (% day−1) | 0.08 ± 0.04 a | 0.28 ± 0.10 a | −1.50 ± 0.28 b |
| Survival (%) | 88.00 ± 7.21 a | 95.33 ± 1.15 a | 55.33 ± 8.08 b |
| Change in individual weight (g) | 0.02 ± 0.01 a | 0.05 ± 0.02 a | −0.21 ± 0.03 b |
| Weight gain (%) | 2.56 ± 1.23 a | 8.76 ± 3.32 a | −36.13 ± 5.24 b |
| 10% Sludge | 20% Sludge | Control | |
|---|---|---|---|
| Density (org m−2) | 300 | 300 | 300 |
| Biomass (g m−2) | 59.85 ± 2.71 a | 60.72 ± 5.30 a | 58.71 ± 4.28 a |
| TOM RR (g m−2 day−1) | 17.30 ± 1.20 ab | 23.95 ± 13.19 a | 4.69 ± 0.47 b |
| TOM removal (%) | 51.90 ± 3.61 | 35.92 ± 19.78 | 42.13 ± 8.09 |
| 10% Sludge | 20% Sludge | Control | |
|---|---|---|---|
| N total (%) | 9.29 ± 0.88 | 8.28 ± 1.38 | 5.51 ± 2.86 |
| C total (%) | 43.13 ± 1.20 | 40.52 ± 5.17 | 32.94 ± 9.49 |
| C/N ratio | 4.66 ± 0.30 | 4.92 ± 0.22 | 6.71 ± 2.17 |
| Protein content (g/100 g) | 58.03 ± 5.47 | 51.74 ± 8.62 | 34.45 ± 17.90 |
| Water content (%) | 82.05 ± 1.42 | 82.12 ± 1.70 | 81.59 ± 2.66 |
| Dry matter (mg/g WW) | 179.53 ± 14.22 | 178.75 ± 17.03 | 184.08 ± 26.60 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez, S.; Lara, G.; Hurtado, C.F.; Espinoza Alvarado, R.; Gutiérrez, J.; Huechucoy, J.C.; Valenzuela-Olea, G.; Turner, A. Evaluating the Bioremediation Capacity of the Polychaete Perinereis gualpensis (Jeldes, 1963) for Atlantic Salmon Aquaculture Sludge. Fishes 2023, 8, 417. https://doi.org/10.3390/fishes8080417
Gómez S, Lara G, Hurtado CF, Espinoza Alvarado R, Gutiérrez J, Huechucoy JC, Valenzuela-Olea G, Turner A. Evaluating the Bioremediation Capacity of the Polychaete Perinereis gualpensis (Jeldes, 1963) for Atlantic Salmon Aquaculture Sludge. Fishes. 2023; 8(8):417. https://doi.org/10.3390/fishes8080417
Chicago/Turabian StyleGómez, Silvia, Gabriele Lara, Carlos Felipe Hurtado, René Espinoza Alvarado, Jayro Gutiérrez, José Carlos Huechucoy, Guillermo Valenzuela-Olea, and Alice Turner. 2023. "Evaluating the Bioremediation Capacity of the Polychaete Perinereis gualpensis (Jeldes, 1963) for Atlantic Salmon Aquaculture Sludge" Fishes 8, no. 8: 417. https://doi.org/10.3390/fishes8080417
APA StyleGómez, S., Lara, G., Hurtado, C. F., Espinoza Alvarado, R., Gutiérrez, J., Huechucoy, J. C., Valenzuela-Olea, G., & Turner, A. (2023). Evaluating the Bioremediation Capacity of the Polychaete Perinereis gualpensis (Jeldes, 1963) for Atlantic Salmon Aquaculture Sludge. Fishes, 8(8), 417. https://doi.org/10.3390/fishes8080417