
Molecular Characterization and Expression Analysis of NLRC3-like, ASC, and Caspase1 in Spotted Sea Bass (Lateolabrax maculatus)

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish
2.2. RNA Extraction and cDNA Synthesis
2.3. Gene Cloning
2.4. LPS and Poly (I:C) Stimulation
2.5. E. tarda Culture and Infection
2.6. qPCR for Expression Analysis
2.7. Bioinformatics Analysis
2.8. Statistical Analysis of Data
3. Results
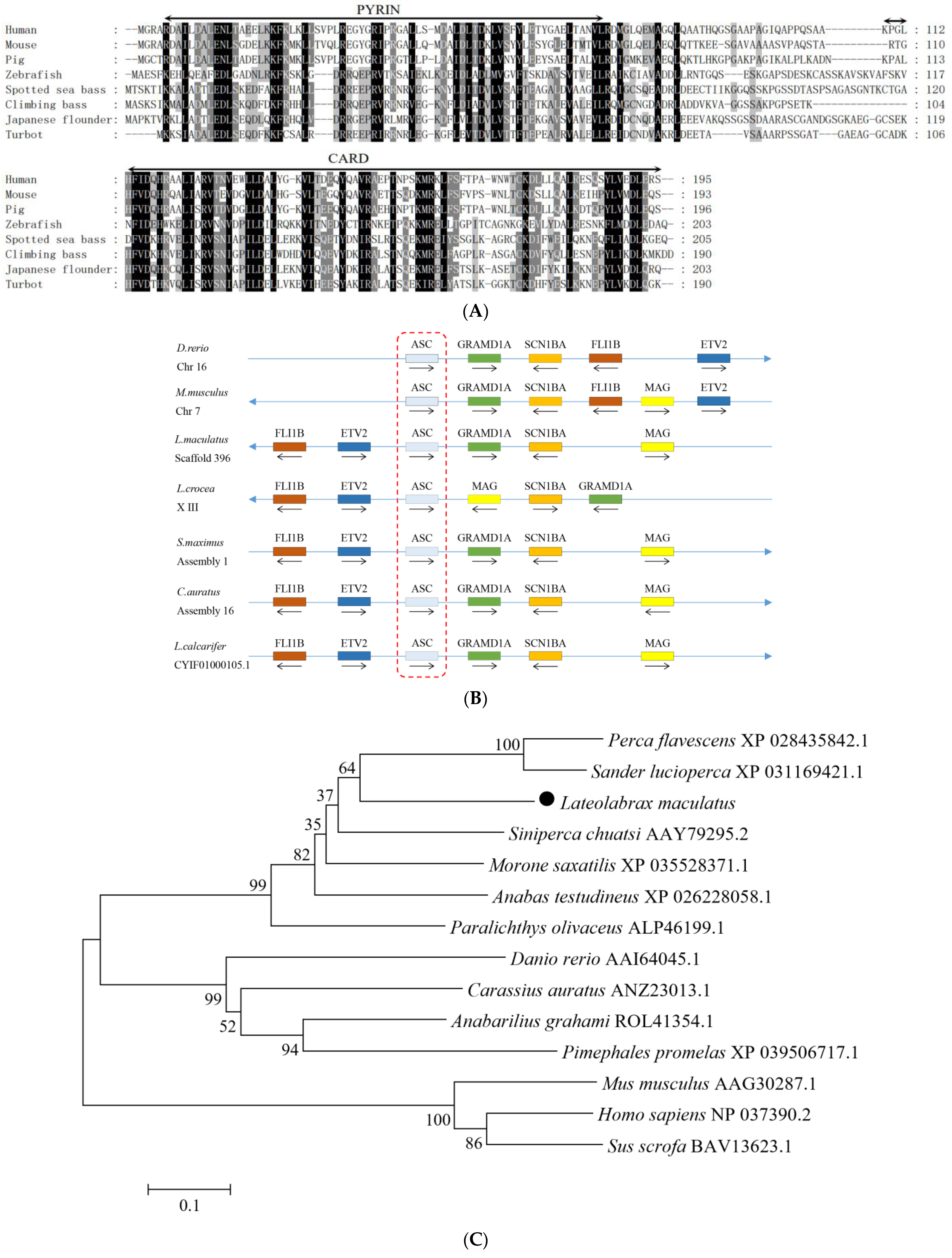
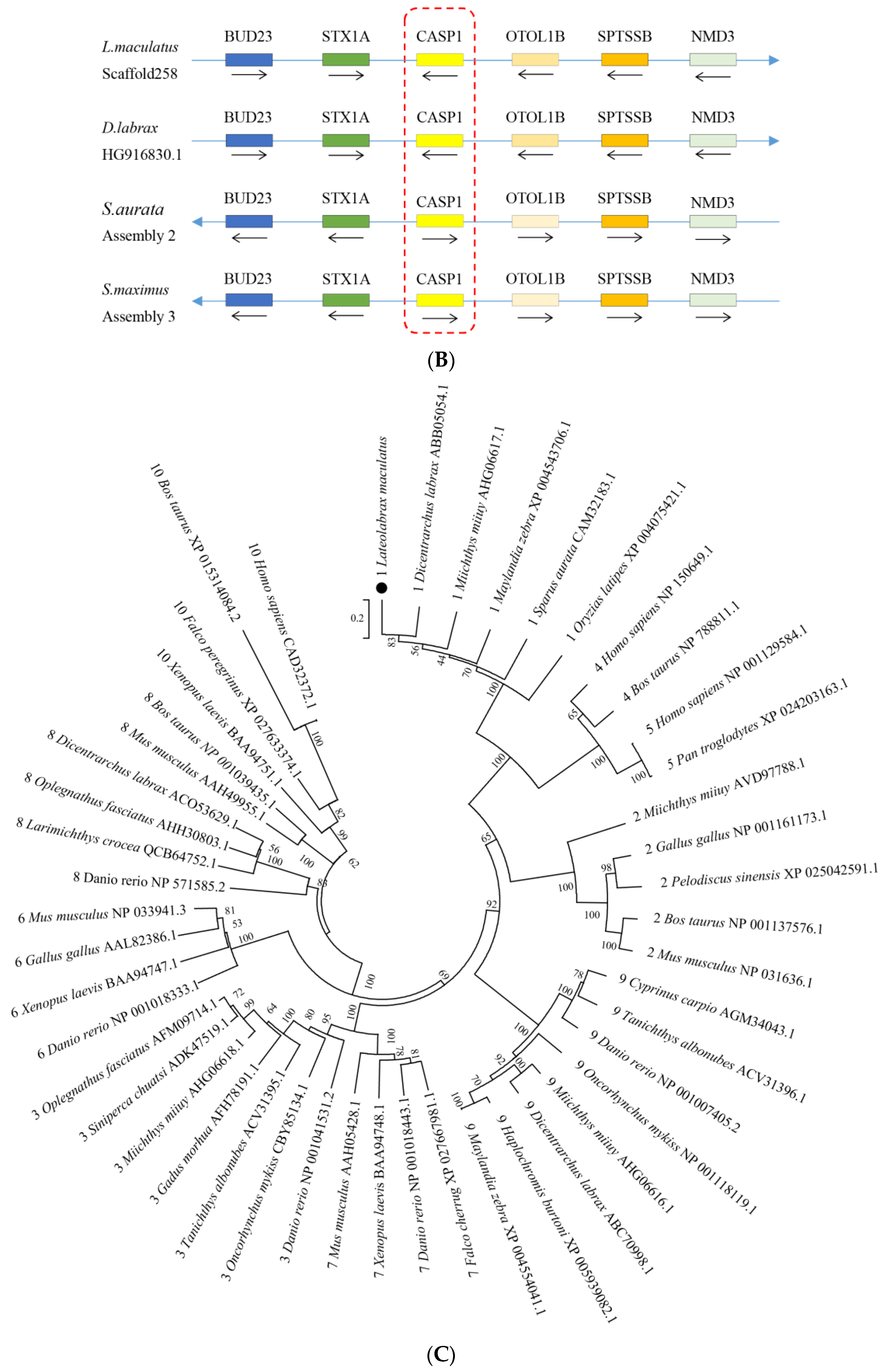
3.1. Sequence Features of NLRC3L, ASC, and Caspase1 of Spotted Sea Bass
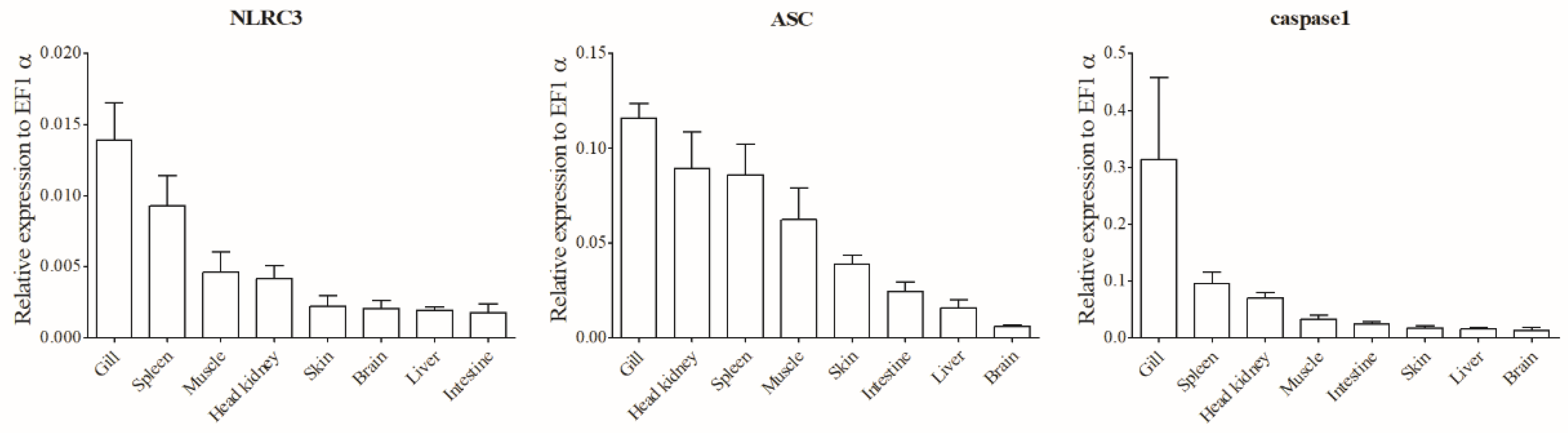
3.2. Tissue Distributions of NLRC3L, ASC, and Caspase1 of Spotted Sea Bass
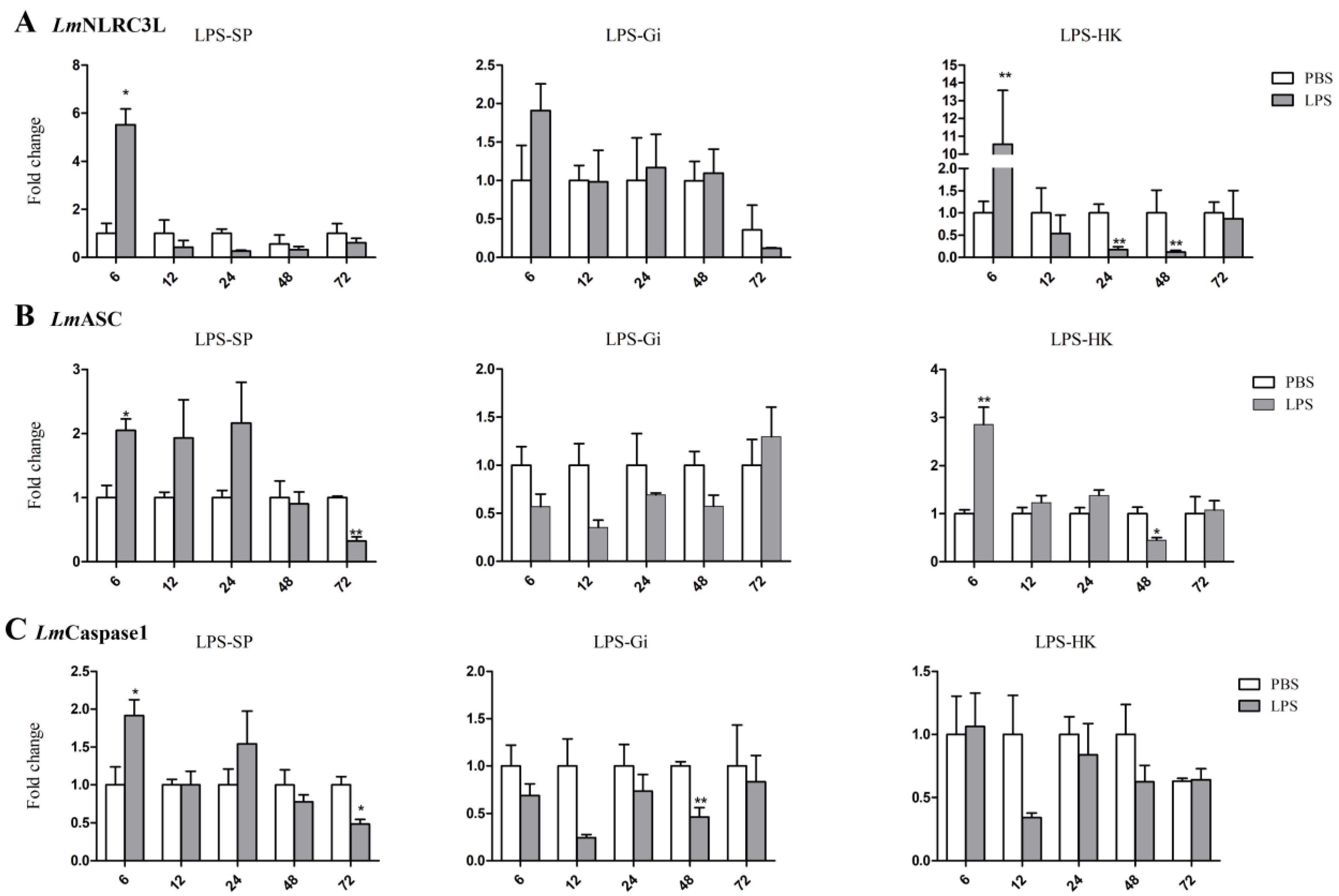
3.3. Expressions of NLRC3L, ASC, and Caspase1 of Spotted Sea Bass Following LPS Stimulation
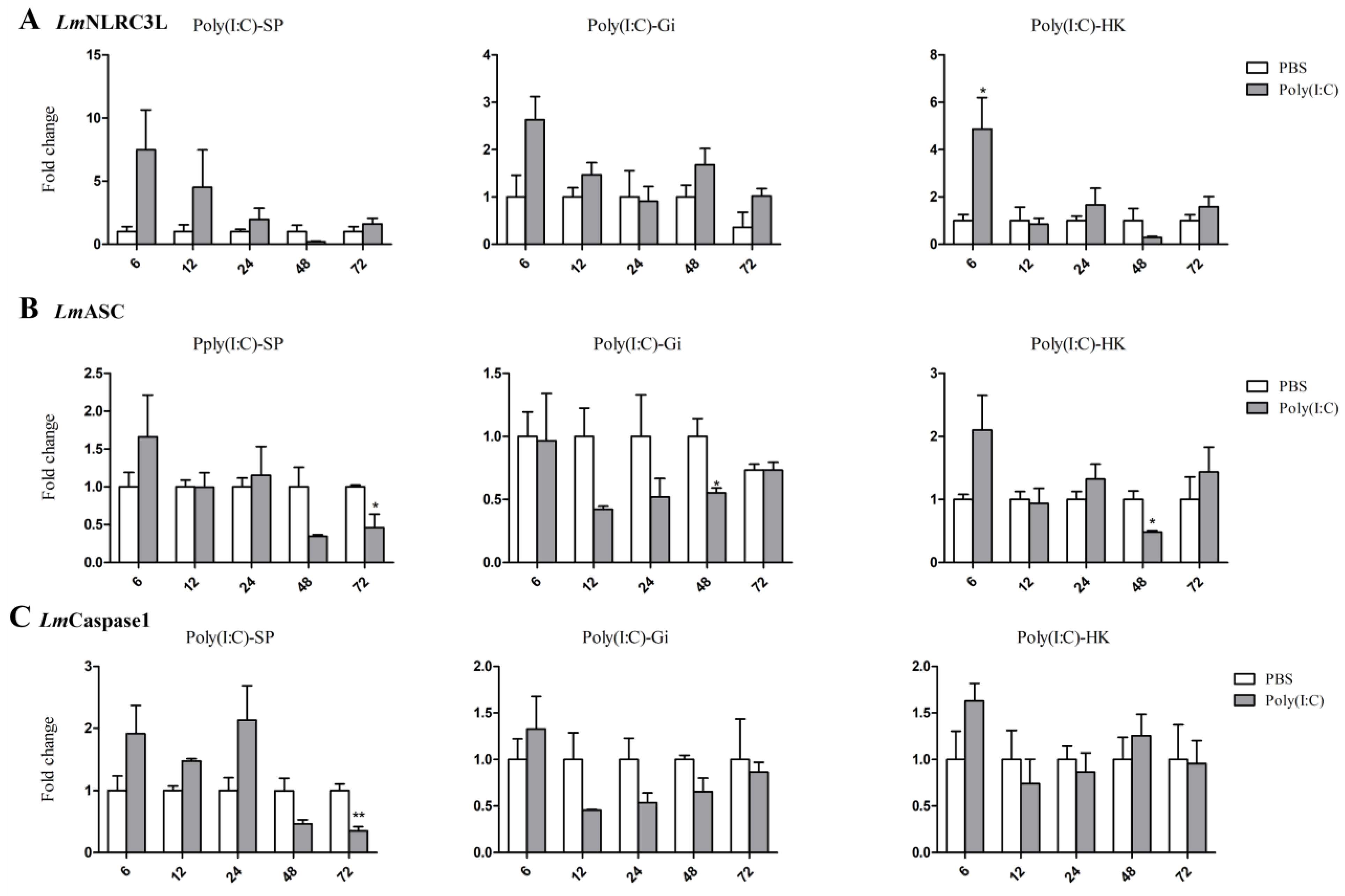
3.4. Expressions of NLRC3L, ASC, and Caspase1 of Spotted Sea Bass Following Poly (I:C) Stimulation
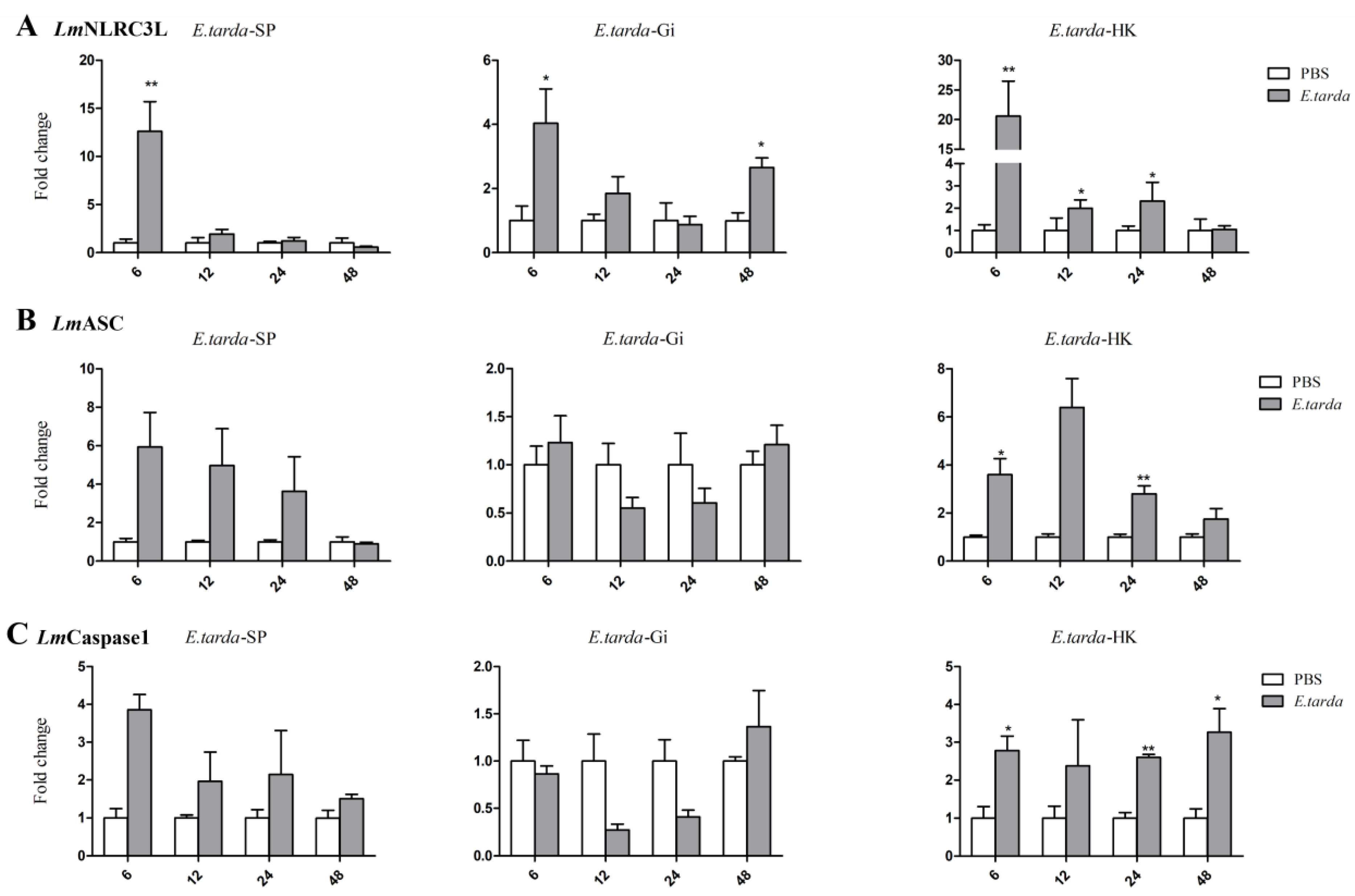
3.5. Expressions of NLRC3L, ASC, and Caspase1 of Spotted Sea Bass Following E. tarda Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Medzhitov, R. Recognition of microorganisms and activation of the immune response. Nature 2007, 449, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Chen, X.L.; Hao, G.X.; Geng, X.Y.; Zhan, W.B.; Sun, J.S. Identification and characterization of a novel NOD-like receptor family CARD domain containing 3 gene in response to extracellular ATP stimulation and its role in regulating LPS-induced innate immune response in Japanese flounder (Paralichthys olivaceus) head kidney macrophages. Fish Shellfish Immunol. 2016, 50, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Pontillo, A.; Crovella, S. NOD-Like Receptors: A Tail from Plants to Mammals Through Invertebrates. Curr. Protein Pept. Sc. 2017, 18, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Motyan, J.A.; Bagossi, P.; Benko, S.; Tozser, J. A molecular model of the full-length human NOD-like receptor family CARD domain containing 5 (NLRC5) protein. BMC Bioinform. 2013, 14, 275. [Google Scholar] [CrossRef] [PubMed]
- Gultekin, Y.; Eren, E.; Ozoren, N. Overexpressed NLRC3 Acts as an Anti-Inflammatory Cytosolic Protein. J. Innate Immun. 2015, 7, 25–36. [Google Scholar] [CrossRef]
- Eren, E.; Berber, M.; Ozoren, N. NLRC3 protein inhibits inflammation by disrupting NALP3 inflammasome assembly via competition with the adaptor protein ASC for pro-caspase-1 binding. J. Biol. Chem. 2017, 292, 12691–12701. [Google Scholar] [CrossRef]
- Reis, M.I.R.; do Vale, A.; Pereira, P.J.B.; Azevedo, J.E.; dos Santos, N.M.S. Caspase-1 and IL-1β Processing in a Teleost Fish. PLoS ONE 2012, 7, e50450. [Google Scholar] [CrossRef]
- Ghayur, T.; Banerjee, S.; Hugunin, M.; Butler, D.; Herzog, L.; Carter, A.; Quintal, L.; Sekut, L.; Talanian, R.; Paskind, M.; et al. Caspase-1 processes IFN-γ-inducing factor and regulates LPS-induced IFN-γ production. Nature 1997, 386, 619–623. [Google Scholar] [CrossRef]
- Vanaja, S.K.; Rathinam, V.A.K.; Fitzgerald, K.A. Mechanisms of inflammasome activation: Recent advances and novel insights. Trends Cell Biol. 2015, 25, 308–315. [Google Scholar] [CrossRef]
- Chuphal, B.; Rai, U.; Roy, B. Teleost NOD-like receptors and their downstream signaling pathways: A brief review. Fish Shellfish Immunol. Rep. 2022, 3, 100056. [Google Scholar] [CrossRef]
- Chang, M.X.; Xiong, F.; Wu, X.M.; Hu, Y.W. The expanding and function of NLRC3 or NLRC3-like in teleost fish: Recent advances and novel insights. Dev. Comp. Immunol. 2021, 114, 103859. [Google Scholar] [CrossRef] [PubMed]
- Sha, Z.X.; Abernathy, J.W.; Wang, S.L.; Li, P.; Kucuktas, H.; Liu, H.; Peatman, E.; Liu, Z.J. NOD-like subfamily of the nucleotide-binding domain and leucine-rich repeat containing family receptors and their expression in channel catfish. Dev. Comp. Immunol. 2009, 33, 991–999. [Google Scholar] [CrossRef] [PubMed]
- Unajak, S.; Santos, M.D.; Hikima, J.; Jung, T.S.; Kondo, H.; Hirono, I.; Aoki, T. Molecular characterization, expression and functional analysis of a nuclear oligomerization domain proteins subfamily C (NLRC) in Japanese flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2011, 31, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Shiau, C.E.; Monk, K.R.; Joo, W.; Talbot, W.S. An Anti-inflammatory NOD-like Receptor Is Required for Microglia Development. Cell Rep. 2013, 5, 1342–1352. [Google Scholar] [CrossRef] [PubMed]
- Li, J.R.; Kong, L.C.; Gao, Y.H.; Wu, C.Q.; Xu, T.J. Characterization of NLR-A subfamily members in miiuy croaker and comparative genomics revealed NLRX1 underwent duplication and lose in actinopterygii. Fish Shellfish Immunol. 2015, 47, 397–406. [Google Scholar] [CrossRef]
- Paria, A.; Deepika, A.; Sreedharan, K.; Makesh, M.; Chaudhari, A.; Purushothaman, C.S.; Thirunavukkarasu, A.R.; Rajendran, K.V. Identification of Nod like receptor C3 (NLRC3) in Asian seabass, Lates calcarifer: Characterisation, ontogeny and expression analysis after experimental infection and ligand stimulation. Fish Shellfish Immunol. 2016, 55, 602–612. [Google Scholar] [CrossRef]
- Zhou, F.J.; Zhan, Q.F.; Ding, Z.J.; Su, L.N.; Fan, J.; Cui, L.; Chen, N.; Wang, W.M.; Liu, H. A NLRC3-like gene from blunt snout bream (Megalobrama amblycephala): Molecular characterization, expression and association with resistance to Aeromonas hydrophila infection. Fish Shellfish Immunol. 2017, 63, 213–219. [Google Scholar] [CrossRef]
- Alvarez, C.A.; Ramirez-Cepeda, F.; Santana, P.; Torres, E.; Cortes, J.; Guzman, F.; Schmitt, P.; Mercado, L. Insights into the diversity of NOD-like receptors: Identification and expression analysis of NLRC3, NLRC5 and NLRX1 in rainbow trout. Mol. Immunol. 2017, 87, 102–113. [Google Scholar] [CrossRef]
- Hou, Z.M.; Ye, Z.; Zhang, D.D.; Gao, C.B.; Su, B.F.; Song, L.; Tan, F.H.; Song, H.H.; Wang, Y.; Li, C. Characterization and expression profiling of NOD-like receptor C3 (NLRC3) in mucosal tissues of turbot (Scophthalmus maximus L.) following bacterial challenge. Fish Shellfish Immunol. 2017, 66, 231–239. [Google Scholar] [CrossRef]
- Xie, J.S.; Belosevic, M. Characterization and functional assessment of the NLRC3-like molecule of the goldfish (Carassius auratus L.). Dev. Comp. Immunol. 2018, 79, 1–10. [Google Scholar] [CrossRef]
- Gao, F.Y.; Pang, J.C.; Lu, M.X.; Yang, X.L.; Zhu, H.P.; Ke, X.L.; Liu, Z.G.; Cao, J.M.; Wang, M. Molecular characterization, expression and functional analysis of NOD1, NOD2 and NLRC3 in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2018, 73, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.J.; Huang, Y.; Cao, X.C.; Yin, X.Y.; Jin, X.Y.; Liu, S.; Jiang, J.S.; Jiang, W.; Xiao, T.S.; Zhou, R.B.; et al. Functional and structural characterization of zebrafish ASC. FEBS J. 2018, 285, 2691–2707. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.N.; Wang, J.; La, H.H.; Yin, Z.X.; He, W.; Weng, S.P.; Yu, X.Q.; Chan, S.M.; He, J.G. Molecular cloning and expression analysis of the ASC gene from mandarin fish and its regulation of NF-κB activation. Dev. Comp. Immunol. 2008, 32, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Chen, X.L.; Peng, W.J.; Hao, G.X.; Geng, X.Y.; Zhan, W.B.; Sun, J.S. Cloning and characterization of apoptosis-associated speck-like protein containing a CARD domain (ASC) gene from Japanese flounder Paralichthys olivaceus. Fish Shellfish Immunol. 2016, 54, 294–301. [Google Scholar] [CrossRef]
- Xie, J.S.; Belosevic, M. Functional characterization of apoptosis-associated speck-like protein (ASC) of the goldfish (Carassius auratus L.). Dev. Comp. Immunol. 2016, 65, 201–210. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, Z.T.; Li, C.; Zhang, Y.; Wan, L.Q.; Wei, J.G.; Qin, Q.W. Characterization of orange-spotted grouper (Epinephelus coioides) ASC and caspase-1 involved in extracellular ATP-mediated immune signaling in fish. Fish Shellfish Immunol. 2020, 97, 58–71. [Google Scholar] [CrossRef]
- Wang, W.H.; Tan, J.C.; Wang, Z.; Zhang, Y.X.; Liu, Q.; Yang, D.H. Characterization of the inflammasome component SmASC in turbot (Scophthalmus maximus). Fish Shellfish Immunol. 2020, 100, 324–333. [Google Scholar] [CrossRef]
- Morimoto, N.; Okamura, Y.; Kono, T.; Sakai, M.; Hikima, J. Characterization and expression analysis of tandemly-replicated asc genes in the Japanese medaka, Oryzias latipes. Dev. Comp. Immunol. 2021, 115, 103894. [Google Scholar] [CrossRef]
- Lopez-Castejon, G.; Sepulcre, M.P.; Mulero, I.; Pelegrin, P.; Meseguer, J.; Mulero, V. Molecular and functional characterization of gilthead seabream Sparus aurata caspase-1: The first identification of an inflammatory caspase in fish. Mol. Immunol. 2008, 45, 49–57. [Google Scholar] [CrossRef]
- Li, S.; Peng, W.J.; Li, J.F.; Hao, G.X.; Geng, X.Y.; Sun, J.S. Characterization of Japanese flounder (Paralichthys olivaceus) Caspase1 involved in extracellular ATP-mediated immune signaling in fish. Fish Shellfish Immunol. 2017, 67, 536–545. [Google Scholar] [CrossRef]
- Kumaresan, V.; Ravichandran, G.; Nizam, F.; Dhayanithi, N.B.; Arasu, M.V.; Al-Dhabi, N.A.; Harikrishnan, R.; Arockiaraj, J. Multifunctional murrel caspase 1, 2, 3, 8 and 9: Conservation, uniqueness and their pathogen-induced expression pattern. Fish Shellfish Immunol. 2016, 49, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.P.; Wang, R.X.; Xu, T.J. Three representative subtypes of caspase in miiuy croaker: Genomic organization, evolution and immune responses to bacterial challenge. Fish Shellfish Immunol. 2014, 40, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.F.; Luo, G.Y.; Liang, R.; Qiu, C.L.; Yang, J.W.; Xie, L.L.; Zhang, K.L.; Tian, Y.; Wang, D.C.; Song, S.; et al. Negative Regulator Nlrc3-like Maintain the Balanced Innate Immune Response During Mycobacterial Infection in Zebrafish. Front. Immunol. 2022, 13, 893611. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.S.; Qin, Y.T.; Liu, D.J.; Wang, B.J.; Jia, Z.; Wang, J.Y.; Gao, Q.; Zou, J.; Pang, Y. The evolution and functional characterization of CXC chemokines and receptors in lamprey. Dev. Comp. Immunol. 2021, 116, 103905. [Google Scholar] [CrossRef]
- Yi, L.; Nie, P.; Yu, H.B.; Xie, H.X. Regulation of Type III Secretion of Translocon and Effector Proteins by the EsaB/EsaL/EsaM Complex in Edwardsiella tarda. Infect. Immun. 2017, 85, e00322-1. [Google Scholar] [CrossRef]
- Li, X.; Yuan, S.Y.; Sun, Z.S.; Lei, L.N.; Wan, S.; Wang, J.Y.; Zou, J.; Gao, Q. Gene identification and functional analysis of peptidoglycan recognition protein from the spotted sea bass (Lateolabrax maculatus). Fish Shellfish Immunol. 2020, 106, 1014–1024. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, Q.; Wang, J.Y.; Tian, J.Y.; Song, Y.J.; Xie, H.X.; Chang, M.X.; Nie, P.; Gao, Q.; Zou, J. Transcriptomic responses of S100 family to bacterial and viral infection in zebrafish. Fish Shellfish Immunol. 2019, 94, 685–696. [Google Scholar] [CrossRef]
- Li, M.; Wang, Q.L.; Lu, Y.; Chen, S.L.; Li, Q.; Sha, Z.X. Expression profiles of NODs in channel catfish (Ictalurus punctatus) after infection with Edwardsiella tarda, Aeromonas hydrophila, Streptococcus iniae and channel catfish hemorrhage reovirus. Fish Shellfish Immunol. 2012, 33, 1033–1041. [Google Scholar] [CrossRef]
- Li, J.Y.; Gao, K.; Shao, T.; Fan, D.D.; Hu, C.B.; Sun, C.C.; Dong, W.R.; Lin, A.F.; Xiang, L.X.; Shao, J.Z. Characterization of an NLRP1 Inflammasome from Zebrafish Reveals a Unique Sequential Activation Mechanism Underlying Inflammatory Caspases in Ancient Vertebrates. J. Immunol. 2018, 201, 1946–1966. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence (5′ to 3′) | Application | Efficiency |
|---|---|---|---|
| NLRC3L-CF | CGCAGCTTGTTTCCTCCATCTG | Verify the CDS | / |
| NLRC3L-CR | CGCAGCTTGTTTCCTCCATCTG | Verify the CDS | / |
| NLRC3L-qF | ATGGTGGTTACTTTGTCAGGCAC | Real-time PCR | 93% |
| NLRC3L-qR | ACTGAGGAACCAGACCTGAACCC | Real-time PCR | |
| ASC-F | GGACACACTGGAGGACTTGTC | Partial cloning | / |
| ASC-R | GGTCAGCAATGAGAAATTGCTC | Partial cloning | / |
| ASC-Rf1 | CCTCAAAGCTGGTCGATGTTGC | 3′RACE | / |
| ASC-Rf2 | CCATCTTGGATGAGCTCCTCG | 3′RACE | / |
| ASC-Rr1 | GTGGCGGAACTTGGCAAAGTC | 5′RACE | / |
| ASC-Rr2 | CTCCACCCTGTTGCGTCTGAC | 5′RACE | / |
| ASC-CF | GTTTGGGATCTCCTGGTGAGA | Verify the CDS | / |
| ASC-CR | GTTCAGGTAGACTGTACGTTTGG | Verify the CDS | / |
| ASC-qF | GTGGATAAACATCGAGTCGAGC | Real-time PCR | 86% |
| ASC-qR | GCAACATCGACCAGCTTTGAG | Real-time PCR | |
| Casp1-F | GGGGAGAAAGACTCAATACTTGAGG | Partial cloning | / |
| Casp1-R | TCTGTCTTTGGTTGCCATCTGTC | Partial cloning | / |
| Casp1-Rf1 | CCCTGACACCGTCTCATATAGACA | 3′RACE | / |
| Casp1-Rf2 | CTCACAGAAGGATGACATTGACGA | 3′RACE | / |
| Casp1-Rr1 | TCAAGTATTGAGTCTTTCTCCCC | 5′RACE | / |
| Casp1-Rr2 | TTCCTGCTGGCAACGTTTCCTT | 5′RACE | / |
| Casp1-Rr3 | AGGACAGACCCAGTTCAGCGTAA | 5′RACE | / |
| Casp1-CF | GAAACATCGTTGTCGTCTCG | Verify the CDS | / |
| Casp1-CR | AATGTGGCTACAAGCCCGTG | Verify the CDS | / |
| Casp1-qF1 | CTGTCCTTGGTGTCAACTGGAC | Real-time PCR | 90% |
| Casp1-qR1 | CTATATGAGACGGTGTCAGGGGTG | Real-time PCR | |
| Ef-1α-qF | ATCTCTGGATGGCACGGAGA | Real-time PCR | 97% |
| Ef-1α-qR | CAGTGTGGTTCCGCTAGCAT | Real-time PCR | |
| APG | CCAGACTCGTGGCTGATGCAGGGGGGGGGGGGGGGG | 5′RACE | / |
| AP | CCAGACTCGTGGCTGATGCA | 5′RACE | / |
| UPM long | CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT | 3′RACE | / |
| UPM short | CTAATACGACTCACTATAGGGC | 3′RACE | / |
| NUP | AAGCAGTGGTATCAACGCAGAGT | 3′RACE | / |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, S.; Sun, Z.; Gao, Q.; Li, Z.; Qi, Z.; Zheng, S.; Liu, D. Molecular Characterization and Expression Analysis of NLRC3-like, ASC, and Caspase1 in Spotted Sea Bass (Lateolabrax maculatus). Fishes 2023, 8, 378. https://doi.org/10.3390/fishes8070378
Yuan S, Sun Z, Gao Q, Li Z, Qi Z, Zheng S, Liu D. Molecular Characterization and Expression Analysis of NLRC3-like, ASC, and Caspase1 in Spotted Sea Bass (Lateolabrax maculatus). Fishes. 2023; 8(7):378. https://doi.org/10.3390/fishes8070378
Chicago/Turabian StyleYuan, Shuya, Zhaosheng Sun, Qian Gao, Zhen Li, Zhitao Qi, Sidi Zheng, and Danjie Liu. 2023. "Molecular Characterization and Expression Analysis of NLRC3-like, ASC, and Caspase1 in Spotted Sea Bass (Lateolabrax maculatus)" Fishes 8, no. 7: 378. https://doi.org/10.3390/fishes8070378
APA StyleYuan, S., Sun, Z., Gao, Q., Li, Z., Qi, Z., Zheng, S., & Liu, D. (2023). Molecular Characterization and Expression Analysis of NLRC3-like, ASC, and Caspase1 in Spotted Sea Bass (Lateolabrax maculatus). Fishes, 8(7), 378. https://doi.org/10.3390/fishes8070378