

Immunoregulation and Resistance to Aquatic Pathogens with Dietary Nucleotides in Pacific White Shrimp, Litopenaeus vannamei



Abstract
1. Introduction
2. Materials and Methods
2.1. Feeding Trial
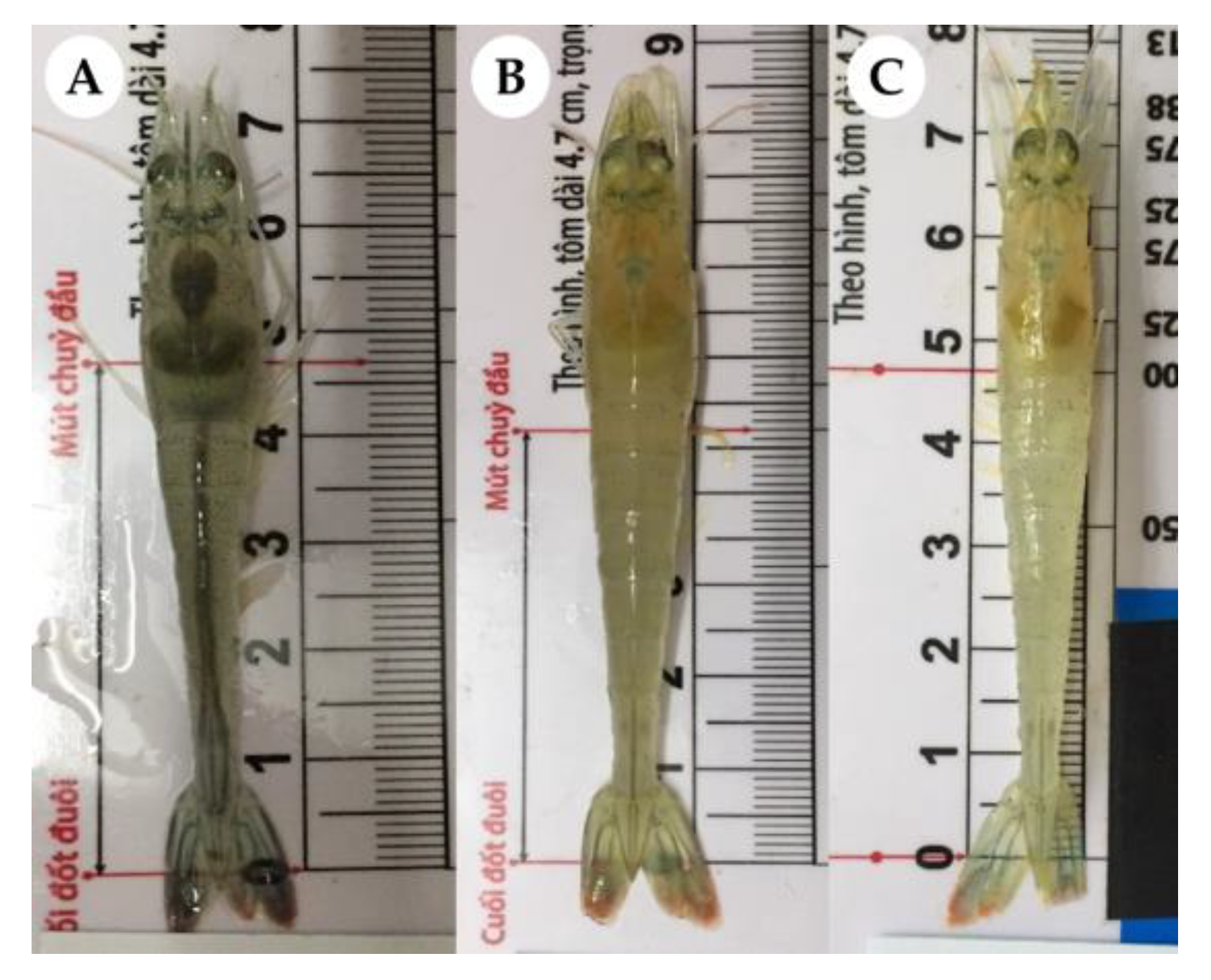
2.2. Challenge Trial
2.3. Statistical Analysis
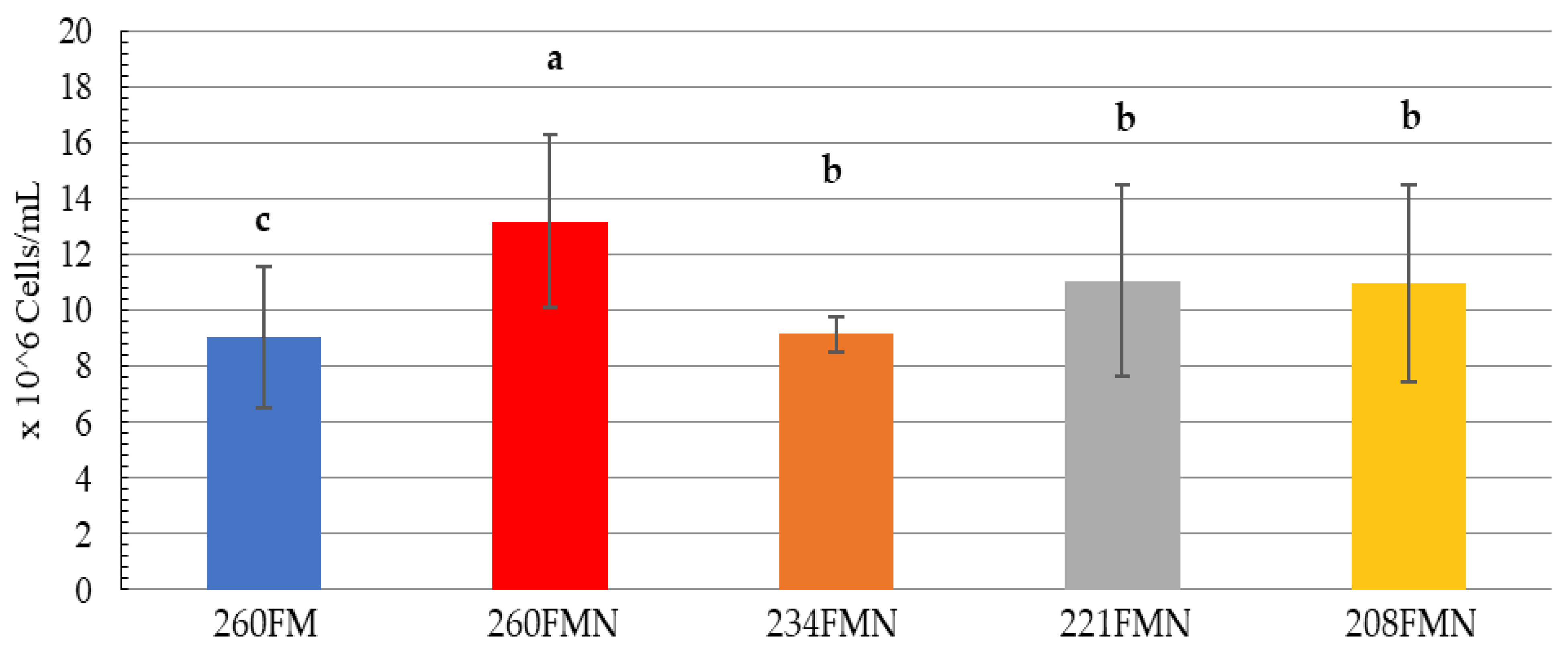
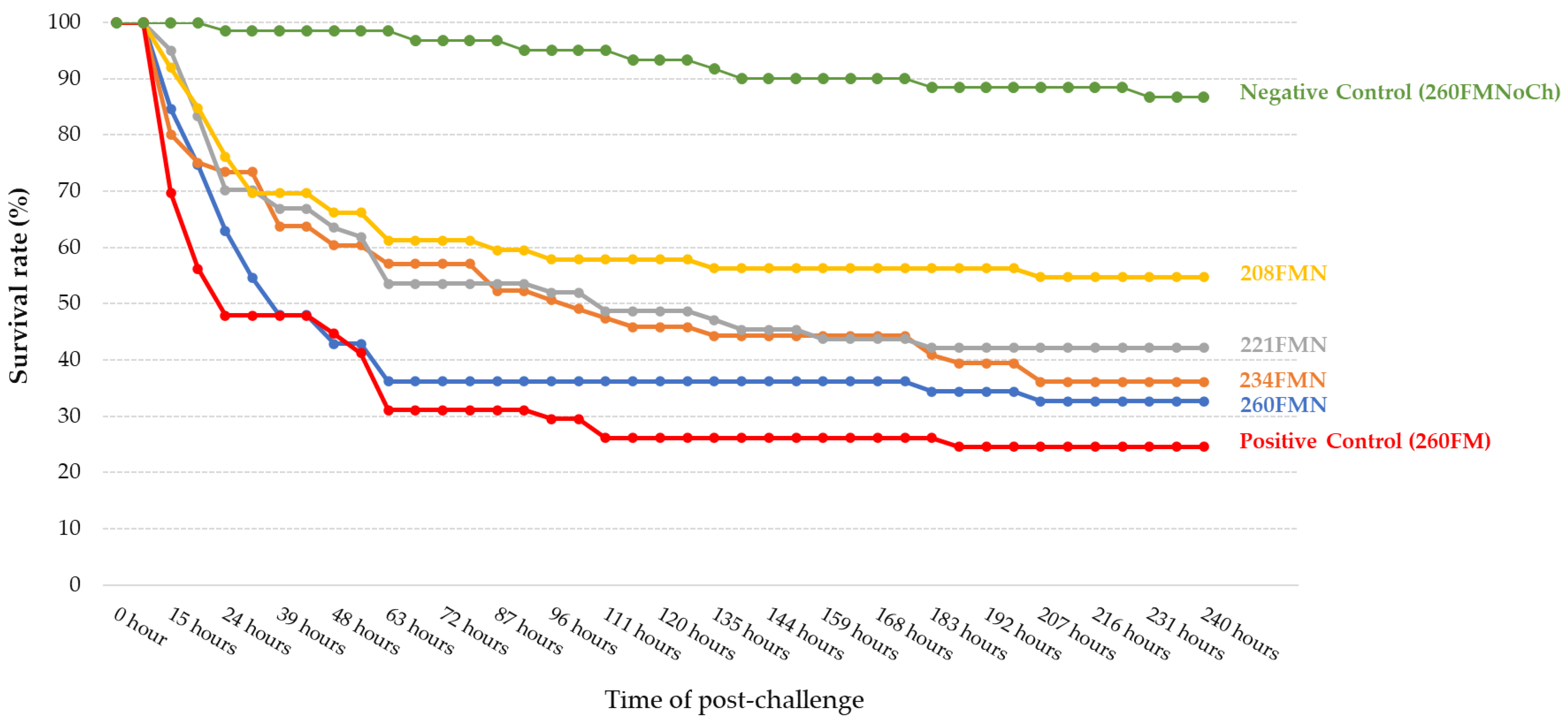
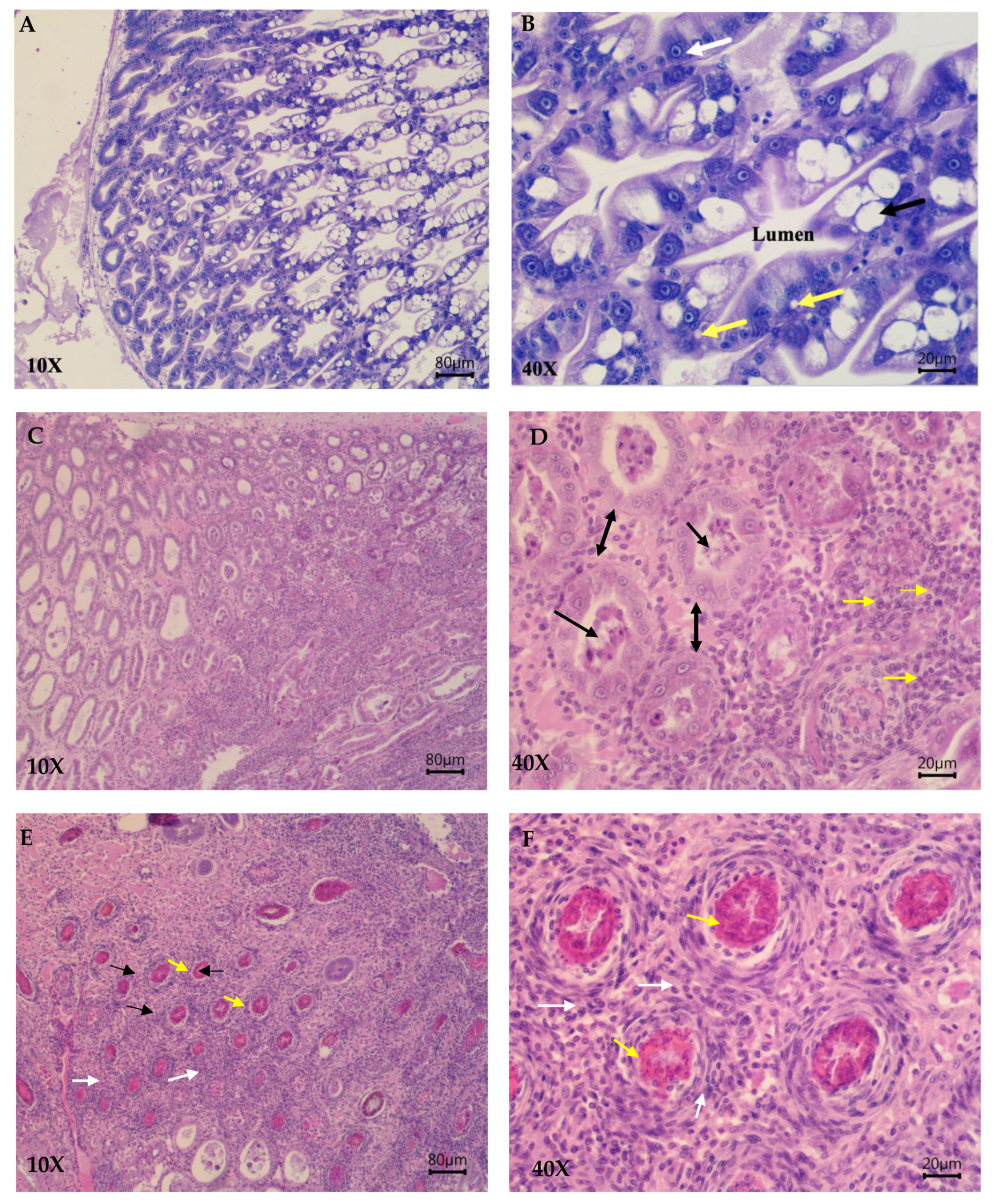
3. Results
3.1. Feeding Trial
3.2. Challenge Trial
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2022; Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar]
- Vietnam Directorate of Fisheries. Report No. 331/BC-TCTK; General Statistics Office of Vietnam: Hanoi, Vietnam, 2022.
- Yue, Y.R.; Liu, Y.J.; Tian, L.X.; Gan, L.; Yang, H.J.; Liang, G.Y. Effects of Replacing Fish Meal with Soybean Meal and Peanut Meal on Growth, Feed Utilization and Haemolymph Indexes for Juvenile White Shrimp Litopenaeus Vannamei, Boone. Aquac. Res. 2012, 43, 1687–1696. [Google Scholar] [CrossRef]
- Kumar, R.; Ng, T.H.; Wang, H.C. Acute Hepatopancreatic Necrosis Disease in Penaeid Shrimp. Rev. Aquac. 2020, 12, 1867–1880. [Google Scholar] [CrossRef]
- Tran, L.; Nunan, L.; Redman, R.M.; Mohney, L.L.; Pantoja, C.R.; Fitzsimmons, K.; Lightner, D.V. Determination of the Infectious Nature of the Agent of Acute Hepatopancreatic Necrosis Syndrome Affecting Penaeid Shrimp. Dis. Aquat. Organ. 2013, 105, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Shinn, A.P.; Pratoomyot, J.; Griffiths, D.; Trong, T.Q.; Vu, N.T.; Jiravanichpaisal, P.; Briggs, M. Asian Shrimp Production and the Economic Costs of Disease. Asian Fish. Sci. 2018, 661, 29–58. [Google Scholar] [CrossRef]
- Kumar, R.; Chauhan, S.B.; Ng, S.S.; Sundar, S.; Engwerda, C.R. Immune Checkpoint Targets for Host-Directed Therapy to Prevent and Treat Leishmaniasis. Front. Immunol. 2017, 8, 1492. [Google Scholar] [CrossRef]
- Segarra, S. Nutritional Modulation of the Immune Response Mediated by Nucleotides in Canine Leishmaniosis. Microorganisms 2021, 9, 2601. [Google Scholar] [CrossRef]
- Fontana, L.; Martínez-Augustin, O.; Gil, Á. Role of Dietary Nucleotides in Immunity. Funct. Food Rev. 2010, 2, 91–100. [Google Scholar] [CrossRef]
- Gil, A. Modulation of the Immune Response Mediated by Dietary Nucleotides. Eur. J. Clin. Nutr. 2002, 56, S1–S4. [Google Scholar] [CrossRef]
- Hess, J.R.; Greenberg, N.A. The Role of Nucleotides in the Immune and Gastrointestinal Systems: Potential Clinical Applications. Nutr. Clin. Pract. 2012, 27, 281–294. [Google Scholar] [CrossRef]
- Reda, R.M.; Selim, K.M.; Mahmoud, R.; El-Araby, I.E. Effect of Dietary Yeast Nucleotide on Antioxidant Activity, Non-Specific Immunity, Intestinal Cytokines, and Disease Resistance in Nile Tilapia. Fish Shellfish Immunol. 2018, 80, 281–290. [Google Scholar] [CrossRef]
- Selim, K.M.; Reda, R.M.; Mahmoud, R.; El-Araby, I.E. Effects of Nucleotides Supplemented Diets on Growth Performance and Expressions of Ghrelin and Insulin-like Growth Factor Genes in Nile Tilapia, Oreochromis Niloticus. J. Appl. Aquac. 2020, 32, 157–174. [Google Scholar] [CrossRef]
- Yaseen, M.S.; Abdelaziz, M.; Abdel-Moneam, D.A.; Abd-Elhay, E.; Wassif, I.M.; Moustafa, M. Efficacy of Dietary Nucleotides (NucleoforceTM) on Growth, Haemato-Immunological Response and Disease Resistance in Pangasianodon Hypophthalmus Fish (Sauvage, 1878) in Egypt. Egypt. J. Aquat. Biol. Fish. 2020, 24, 405–424. [Google Scholar] [CrossRef]
- El-Nokrashy, A.M.; El-Banna, R.A.; Edrise, B.M.; Abdel-Rahim, M.M.; Jover-Cerdá, M.; Tomás-Vidal, A.; Prince, A.; Davies, S.J.; El-Haroun, E.R.; Goda, A.M.A.S. Impact of Nucleotide Enriched Diets on the Production of Gilthead Seabream, Sparus Aurata Fingerlings by Modulation of Liver Mitochondrial Enzyme Activitity, Antioxidant Status, Immune Gene Expression, and Gut Microbial Ecology. Aquaculture 2021, 535, 736398. [Google Scholar] [CrossRef]
- Novriadi, R.; Ilham, I.; Roigé, O.; Segarra, S. Effects of Dietary Nucleotides Supplementation on Growth, Total Haemocyte Count, Lysozyme Activity and Survival upon Challenge with Vibrio Harveyi in Pacific White Shrimp, Litopenaeus Vannamei. Aquac. Rep. 2021, 21, 100840. [Google Scholar] [CrossRef]
- Borda, E.; Estévez, A.; Tort, L. Effect of the Free Nucleotides on the Immune Response during First Stage of Fattening in Gilthhead Bream (Sparus Aurata). In Proceedings of the Aquaculture Europe, Trondheim, Norway, 5–9 August 2005; pp. 5–9. [Google Scholar]
- Estruch, G.; Tortosa, I.; Monge, R.; Godoy, S. Inclusión de Aditivos En Dietas Vegetales Para Doradas. Efectos Sobre El Crecimiento, Parámetros Biométricos y Nutritivos, Digestibilidad e Histología. In Proceedings of the Actas del XV Congreso Nacional y I Congreso Ibérico de Acuicultura, Guayaquil, Ecuador, 19–22 October 2015. [Google Scholar]
- Magouz, F.I.; Abdel-Rahim, M.M.; Lotfy, A.M.; Mosbah, A.; Alkafafy, M.; Sewilam, H.; Dawood, M.A.O. Dietary Nucleotides Enhanced Growth Performance, Carcass Composition, Blood Biochemical, and Histology Features of European Sea Bass, Dicentrarchus labrax L. Aquac. Rep. 2021, 20, 100738. [Google Scholar] [CrossRef]
- Borda, E.; de la Fuente, E. Efficacy of Dietary Nucleotides Supplementation on the Immune Response in Atlantic Salmon (Salmo salar): Effects on Salt Water Transfer and Vaccination. In Proceedings of the Aquaculture Europe, Krakow, Poland, 15–18 September 2008. [Google Scholar]
- Sáenz de Rodrigáñez, M.A.; Barata, M.; Dias, J.; Pousão-Ferreira, P.; Morales, G.; Márquez, L.; Moyano, F.J.; Ribeiro, L. Evaluation of the Effect of Nucleotides on Intestinal Function in Juveniles of Meagre (Argyrosomus regius) Fed on High Plant Protein Diets. In Proceedings of the Aquaculture Europe, Prague, Czech Republic, 1–5 September 2012. [Google Scholar]
- Romano, N.; Fischer, H.; Rossi, W.; Quintero, H.; Limbaugh, N. Effects of Bioprocessed Soybean Meal and Nucleotide Supplementation on Growth, Physiology and Histomorphology in Largemouth Bass, Micropterus Salmoides, Juveniles. Comp. Biochem. Physiol. 2021, 260, 111038. [Google Scholar] [CrossRef]
- Novriadi, R.; Roigé, O.; Segarra, S. Effects of Dietary Nucleotide Supplementation on Performance, Profitability, and Disease Resistance of Litopenaeus Vannamei Cultured in Indonesia under Intensive Outdoor Pond Conditions. Animals 2022, 12, 2036. [Google Scholar] [CrossRef]
- Mastan, S.A. Use of Immunostimulants in Aquaculture Disease Management. Int. J. Fish. Aquat. Stud. 2015, 2, 277–280. [Google Scholar]
- del Valle, J.C.; Bonadero, M.C.; Ferández-Gimenez, A.V. Saccharomyces Cerevisiae as Probiotic, Prebiotic, Synbiotic, Postbiotics and Parabiotics in Aquaculture: An Overview. Aquaculture 2023, 1, 739342. [Google Scholar] [CrossRef]
- Bell, T.A.; Lightner, D.V. A Handbook of Normal Penaeid Shrimp Histology; World Aquaculture Society: Baton Rouge, LA, USA, 1988. [Google Scholar]
- Lightner, D.V. A Handbook of Shrimp Pathology and Diagnostic Procedures for Diseases of Cultured Penaeid Shrimp; World Aquaculture Society: Baton Rouge, LA, USA, 1996. [Google Scholar]
- Tang, K.F.J.; Bondad-Reantaso, M.G.; Arthur, J.R.; MacKinnon, B.; Hao, B.; Alday-Sanz, V.; Liang, Y.; Dong, X. Shrimp Acute Hepatopancreatic Necrosis Disease Strategy Manual; FAO Fisheries and Aquaculture Circular No. 1190; FAO: Rome, Italy, 2020. [Google Scholar]
- Hoe, T.D. Finding the Reason for the High Production Cost of Vietnamese Shrimp. Vietnamagriculture 2023. Available online: https://vietnamagriculture.nongnghiep.vn/finding-the-reason-for-the-high-production-cost-of-vietnamese-shrimp-d348607.html (accessed on 31 May 2023).
- Luc, H.Q. Vietnamese Shrimp’s Global Integration. Vietnam Fisheries Magazine, 28 June 2021; 10–13. [Google Scholar]
- Vicentini, I.B.F.; Ribeiro, K.; Papa, L.P.; Junior, J.M.; Vicentini, C.A.; Moraes-Valenti, P.M.C. Histoarchitectural features of the hepatopancreas of the Amazon river prawn Macrobrachium amazonicum. Int. J. Morphol. 2009, 27, 121–128. [Google Scholar]
- Wang, X.; Li, E.; Xu, C.; Qin, J.G.; Wang, S.; Chen, X.; Cai, Y.; Chen, K.; Gan, L.; Yu, N.; et al. Growth, body composition, ammonia tolerance and hepatopancreas histology of white shrimp Litopenaeus vannamei fed diets containing diferente carbohydrate sources at low salinity. Aquac. Res. 2014, 47, 1932–1943. [Google Scholar] [CrossRef]
- Lee, P.G.; Meyers, S.P. Chemoattraction and Feeding Stimulation. In Crustacean Nutrition, Advances in World Aquaculture; The World Aquaculture Society: Baton Rouge, LA, USA, 1997; pp. 292–352. [Google Scholar]
- Suresh, A.V.; Kumaraguru Vasagam, K.P.; Nates, S. Attractability and Palatability of Protein Ingredients of Aquatic and Terrestrial Animal Origin, and Their Practical Value for Blue Shrimp, Litopenaeus Stylirostris Fed Diets Formulated with High Levels of Poultry Byproduct Meal. Aquaculture 2011, 319, 132–140. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (g/kg) | 260FM | 260FMN | 234FMN | 221FMN | 208FMN |
|---|---|---|---|---|---|
| Fish Meal (anchovy) 1 | 260.00 | 260.00 | 234.00 | 221.00 | 208.00 |
| Soybean meal (48% CP) | 360.00 | 360.00 | 386.00 | 400.00 | 413.00 |
| Fish and tuna oil | 16.00 | 16.00 | 17.50 | 18.20 | 18.60 |
| Wheat flour | 278.00 | 277.00 | 278.00 | 282.00 | 283.60 |
| Krill meal 1 | 0.00 | 0.00 | 2.00 | 3.00 | 5.00 |
| Wheat gluten meal (78% CP) | 20.00 | 20.00 | 20.00 | 20.00 | 20.00 |
| Choline chloride (60% choline) | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 |
| Cholesterol feed grade | 0.40 | 0.40 | 0.48 | 0.51 | 0.55 |
| Soy lecithin | 20.00 | 20.00 | 20.00 | 20.00 | 20.00 |
| Mono calcium phosphate | 17.00 | 17.00 | 9.57 | 4.54 | - |
| Salt (NaCl) | 0.00 | 0.00 | 2.00 | - | - |
| Ascorbyl-monophosphate 2 | 1.25 | 1.25 | 1.25 | 1.25 | 1.25 |
| Mix of Vitamin + Minerals 3 | 4.00 | 4.00 | 3.75 | 3.75 | 3.75 |
| DL-Methionine | 2.20 | 2.20 | 2.50 | 2.60 | 2.80 |
| L-Threonine | 0.00 | 0.00 | 0.30 | 0.30 | 0.40 |
| L-Lysine | 0.00 | 0.00 | 0.50 | 0.70 | 0.90 |
| Carboxymethylcellulose | 10.0 | 10.00 | 10.00 | 10.00 | 10.00 |
| Calcium propionate | 0.75 | 0.75 | 0.75 | 0.75 | 0.75 |
| Antioxidant 4 | 0.40 | 0.40 | 0.40 | 0.40 | 0.40 |
| Nucleotides 5 | 0 | 1.0 | 1.0 | 1.0 | 1.0 |
| Diet formulation cost (USD/kg) * | 0.8371 | 0.8849 | 0.8719 | 0.8659 | 0.8618 |
| % vs. Control (260FM) | - | +5.71% | +4.16% | +3.45% | +2.95% |
| Nutrient Component (%) | 260FM | 260FMN | 234FMN | 221FMN | 208FMN |
|---|---|---|---|---|---|
| Moisture | 10.33 | 10.32 | 10.45 | 10.55 | 10.63 |
| Crude protein | 40.12 | 40.10 | 39.83 | 39.76 | 39.69 |
| Crude lipids | 6.77 | 6.77 | 6.76 | 6.75 | 6.73 |
| Crude fiber | 1.35 | 1.35 | 1.41 | 1.45 | 1.48 |
| Ash | 8.04 | 8.04 | 7.29 | 6.50 | 5.96 |
| Lysine | 2.49 | 2.49 | 2.49 | 2.48 | 2.48 |
| Methionine | 1.00 | 1.00 | 1.00 | 0.99 | 1.00 |
| Threonine | 1.51 | 1.51 | 1.52 | 1.52 | 1.52 |
| Cholesterol | 1.47 | 1.47 | 1.48 | 1.47 | 1.48 |
| Phospholipids | 2.26 | 2.26 | 2.25 | 2.25 | 2.24 |
| 260FM | 260FMN | 234FMN | 221FMN | 208FMN | |
|---|---|---|---|---|---|
| Initial mean weight (g) | 1.32 ± 0.08 a | 1.38 ± 0.08 a | 1.36 ± 0.09 a | 1.43 ± 0.12 a | 1.36 ± 0.05 a |
| Initial biomass (g) | 52.80 ± 3.35 a | 55.20 ± 3.35 a | 54.40 ± 3.58 a | 57.20 ± 4.82 a | 54.40 ± 2.19 a |
| Final mean weight (g) | 18.87 ± 1.32 ab | 19.56 ± 1.12 b | 17.81 ± 0.85 a | 18.15 ± 0.93 ab | 17.61 ± 0.87 a |
| Final biomass (g) | 690.32 ± 47.08 ab | 711.08 ± 27.62 b | 665.44 ± 18.52 ab | 671.66 ± 42.18 ab | 654.64 ± 25.57 a |
| Mean weight gain (g) | 17.55 ± 1.30 ab | 18.18 ± 1.08 b | 16.45 ± 0.82 a | 16.72 ± 0.82 a | 16.25 ± 0.88 a |
| ADG (g/day) | 0.31 ± 0.02 ab | 0.32 ± 0.02 b | 0.29 ± 0.01 a | 0.30 ± 0.01 a | 0.29 ± 0.02 a |
| SGR (%/day) | 4.75 ± 0.15 b | 4.74 ± 0.11 b | 4.59 ± 0.12 ab | 4.54 ± 0.07 a | 4.57 ± 0.12 ab |
| Feed consumption (g) | 829.96 ± 19.41 b | 845.62 ± 10.68 b | 844.78 ± 19.51 b | 844.54 ± 10.16 b | 810.68 ± 4.16 a |
| FCR | 1.31 ± 0.10 a | 1.29 ± 0.05 a | 1.38 ± 0.07 a | 1.38 ± 0.09 a | 1.35 ± 0.05 a |
| 260FM | 260FMN | 234FMN | 221FMN | 208FMN | |
|---|---|---|---|---|---|
| TOTAL BIOMASS (kg/tank) | 0.690 | 0.711 | 0.665 | 0.672 | 0.655 |
| Total biomass (kg/m3) | 1.725 | 1.778 | 1.663 | 1.680 | 1.638 |
| Biomass (Tons/ha) | 17.250 | 17.775 | 16.625 | 16.800 | 16.375 |
| Final mean weight (g/shrimp) | 18.87 | 19.56 | 17.81 | 18.15 | 17.61 |
| Counts/kg | 53 | 51 | 56 | 55 | 57 |
| Price (USD/kg) | 4.97 | 5.15 | 4.88 | 4.90 | 4.86 |
| Total cumulative feed (kg/tank) | 0.830 | 0.846 | 0.845 | 0.845 | 0.811 |
| Total cumulative feed (tons/ha) | 20.750 | 21.150 | 21.125 | 21.125 | 20.275 |
| Feed cost (USD/kg) | 0.837 | 0.885 | 0.872 | 0.866 | 0.862 |
| FCR | 1.31 | 1.29 | 1.38 | 1.38 | 1.35 |
| Survival rate (%) | 91.50 | 91.00 | 93.50 | 92.50 | 93.00 |
| Revenue (USD/ha) | 82,972.50 | 88,875.00 | 78,968.75 | 80,304.00 | 77,453.75 |
| Production cost (Feed and related costs) (USD/ha) | 48,758.42 | 51,041.38 | 48,765.74 | 49,139.80 | 47,380.05 |
| Production cost (USD/kg) (1.73 USD/Kg shrimp) | 2.83 | 2.87 | 2.93 | 2.92 | 2.89 |
| Profit (USD/ha) | 39,836.67 | 58,817.67 | 49,128.44 | 50,552.09 | 48,422.60 |
| ROI (%) | 75.83 | 79.35 | 66.37 | 67.52 | 67.97 |
| % improved ROI vs. 260FM | +4.64% | −12.48% | −10.96% | −10.37% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Segarra, S.; Chau, T.; Hoang, P.; Tran, L. Immunoregulation and Resistance to Aquatic Pathogens with Dietary Nucleotides in Pacific White Shrimp, Litopenaeus vannamei. Fishes 2023, 8, 308. https://doi.org/10.3390/fishes8060308
Segarra S, Chau T, Hoang P, Tran L. Immunoregulation and Resistance to Aquatic Pathogens with Dietary Nucleotides in Pacific White Shrimp, Litopenaeus vannamei. Fishes. 2023; 8(6):308. https://doi.org/10.3390/fishes8060308
Chicago/Turabian StyleSegarra, Sergi, Thanh Chau, Phuc Hoang, and Loc Tran. 2023. "Immunoregulation and Resistance to Aquatic Pathogens with Dietary Nucleotides in Pacific White Shrimp, Litopenaeus vannamei" Fishes 8, no. 6: 308. https://doi.org/10.3390/fishes8060308
APA StyleSegarra, S., Chau, T., Hoang, P., & Tran, L. (2023). Immunoregulation and Resistance to Aquatic Pathogens with Dietary Nucleotides in Pacific White Shrimp, Litopenaeus vannamei. Fishes, 8(6), 308. https://doi.org/10.3390/fishes8060308