
Different Diets Based on Yellow Mealworm (Tenebrio molitor)—Part A: Facing the Decrease in Omega−3 Fatty Acids in Fillets of Rainbow Trout (Oncorhynchus mykiss)

 ,
,  , , ,
, , ,  , , , ,
, , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Experimental Animals and Rearing Conditions
2.3. Growth Trial and Sample Collection
2.4. Final Product Quality
2.4.1. Instrumental Texture and Colour
2.4.2. Direct Sensorial Analyses
2.5. Analytical Determinations
2.5.1. Chemical Analyses
2.5.2. Fatty Acid Determination
2.5.3. Liver Intermediary Metabolism
2.5.4. Liver Antioxidant Status and Fish Welfare Indicators
2.6. Histomorphology
2.6.1. Samples Processing
2.6.2. Liver Histomorphology Analysis
2.7. Statistical Analyses
3. Results and Discussion
3.1. Growth Performance
3.2. Final Product Quality
3.2.1. Fillet Proximate and Fatty Acid Composition
3.2.2. Fillet Perceived Quality
3.3. Liver Performance and Antioxidant Status
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- Oliva-Teles, A.; Couto, A.; Enes, P.; Peres, H. Dietary protein requirements of fish—A meta-analysis. Rev. Aquac. 2020, 12, 1445–1477. [Google Scholar] [CrossRef]
- Cashion, T.; Le Manach, F.; Zeller, D.; Pauly, D. Most fish destined for fishmeal production are food-grade fish. Fish Fish. 2017, 18, 837–844. [Google Scholar] [CrossRef]
- Glencross, B.; Rutherford, N.; Jones, B. Evaluating options for fishmeal replacement in diets for juvenile barramundi (Lates calcarifer). Aquac. Nutr. 2011, 17, e722–e732. [Google Scholar] [CrossRef]
- Glencross, B.; Blyth, D.; Irvin, S.; Bourne, N.; Campet, M.; Boisot, P.; Wade, N.M. An evaluation of the complete replacement of both fishmeal and fish oil in diets for juvenile Asian seabass, Lates calcarifer. Aquaculture 2016, 451, 298–309. [Google Scholar] [CrossRef]
- Krogdahl, Å.; Penn, M.; Thorsen, J.; Refstie, S.; Bakke, A.M. Important antinutrients in plant feedstuffs for aquaculture: An update on recent findings regarding responses in salmonids. Aquac. Res. 2010, 41, 333–344. [Google Scholar] [CrossRef]
- Urán, P.A.; Gonçalves, A.A.; Taverne-Thiele, J.J.; Schrama, J.W.; Verreth, J.A.; Rombout, J.H. Soybean meal induces intestinal inflammation in common carp (Cyprinus carpio L.). Fish Shellfish. Immunol. 2008, 25, 751–760. [Google Scholar] [CrossRef]
- Urán, P.A.; Schrama, J.W.; Rombout, J.H.; Taverne-Thiele, J.J.; Obach, A.; Koppe, W.; Verreth, J.A. Time-related changes of the intestinal morphology of Atlantic salmon, Salmo salar L., at two different soybean meal inclusion levels. J. Fish Dis. 2009, 32, 733–744. [Google Scholar] [CrossRef]
- Djordjevic, B.; Morales-Lange, B.; Øverland, M.; Mercado, L.; Lagos, L. Immune and proteomic responses to the soybean meal diet in skin and intestine mucus of Atlantic salmon (Salmo salar L.). Aquac. Nutr. 2021, 27, 929–940. [Google Scholar] [CrossRef]
- Mundheim, H.; Aksnes, A. Growth, feed efficiency and digestibility in salmon (Salmo salar L.) fed different dietary proportions of vegetable protein sources in combination with two fish meal qualities. Aquaculture 2004, 237, 315–331. [Google Scholar] [CrossRef]
- Albrektsen, S.; Mundheim, H.; Aksnes, A. Growth, feed efficiency, digestibility and nutrient distribution in Atlantic cod (Gadus morhua) fed two different fish meal qualities at three dietary levels of vegetable protein sources. Aquaculture 2006, 261, 626–640. [Google Scholar] [CrossRef]
- Van Huis, A. Potential of insects as food and feed in assuring food security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef]
- Van Huis, A. Insects as food and feed, a new emerging agricultural sector: A review. J. Insects Food Feed 2020, 6, 27–44. [Google Scholar] [CrossRef]
- Barroso, F.G.; de Haro, C.; Sánchez-Muros, M.-J.; Venegas, E.; Martínez-Sánchez, A.; Pérez-Bañón, C. The potential of various insect species for use as food for fish. Aquaculture 2014, 422–423, 193–201. [Google Scholar] [CrossRef]
- Makkar, H.P.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Belforti, M.; Gai, F.; Lussiana, C.; Renna, M.; Malfatto, V.; Rotolo, L.; De Marco, M.; Dabbou, S.; Schiavone, A.; Zoccarato, I.; et al. Tenebrio molitor meal in rainbow trout (Oncorhynchus mykiss) diets: Effects on animal performance, nutrient digestibility and chemical composition of fillets. Ital. J. Anim. Sci. 2015, 14, 4170. [Google Scholar] [CrossRef]
- Melenchón, F.; Larrán, A.M.; de Mercado, E.; Hidalgo, M.C.; Cardenete, G.; Barroso, F.G.; Fabrikov, D.; Lourenço, H.M.; Pessoa, M.F.; Tomás-Almenar, C. Potential use of black soldier fly (Hermetia illucens) and mealworm (Tenebrio molitor) insectmeals in diets for rainbow trout (Oncorhynchus mykiss). Aquac. Nutr. 2021, 27, 491–505. [Google Scholar] [CrossRef]
- Melenchón, F.; de Mercado, E.; Pula, H.J.; Cardenete, G.; Barroso, F.G.; Fabrikov, D.; Lourenço, H.M.; Pessoa, M.-F.; Lagos, L.; Weththasinghe, P.; et al. Fishmeal dietary replacement up to 50%: A comparative study of two insect meals for rainbow trout (Oncorhynchus mykiss). Animals 2022, 12, 179. [Google Scholar] [CrossRef]
- Renna, M.; Schiavone, A.; Gai, F.; Dabbou, S.; Lussiana, C.; Malfatto, V.; Prearo, M.; Capucchio, M.T.; Biasato, L.; Biasibetti, E.; et al. Evaluation of the suitability of a partially defatted black soldier fly (Hermetia illucens L.) larvae meal as ingredient for rainbow trout (Oncorhynchus mykiss Walbaum) diets. J. Anim. Sci. 2017, 8, 57. [Google Scholar] [CrossRef]
- Chemello, G.; Renna, M.; Caimi, C.; Guerreiro, I.; Oliva-Teles, A.; Enes, P.; Biasato, I.; Schiavone, A.; Gai, F.; Gasco, L. Partially defatted Tenebrio molitor larva meal in diets for grow-out rainbow trout, Oncorhynchus mykiss (Walbaum): Effects on growth performance, diet digestibility and metabolic responses. Animals 2020, 10, 229. [Google Scholar] [CrossRef]
- Cardinaletti, G.; Di Marco, P.; Daniso, E.; Messina, M.; Donadelli, V.; Finoia, M.G.; Petochi, T.; Fava, F.; Faccenda, F.; Contò, M.; et al. Growth and welfare of rainbow trout (Oncorhynchus mykiss) in response to graded levels of insect and poultry by-product meals in fishmeal-free diets. Animals 2022, 12, 1698. [Google Scholar] [CrossRef]
- Henry, M.A.; Gai, F.; Enes, P.; Pérez-Jiménez, A.; Gasco, L. Effect of partial dietary replacement of fishmeal by yellow mealworm (Tenebrio molitor) larvae meal on the innate immune response and intestinal antioxidant enzymes of rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2018, 83, 308–313. [Google Scholar] [CrossRef]
- Xu, X.; Ji, H.; Yu, H.; Zhou, J. Influence of dietary black soldier fly (Hermetia illucens Linnaeus) pulp on growth performance, antioxidant capacity and intestinal health of juvenile mirror carp (Cyprinus carpio var. specularis). Aquac. Nutr. 2020, 26, 432–443. [Google Scholar] [CrossRef]
- Kumar, V.; Fawole, F.J.; Romano, N.; Hossain, M.S.; Labh, S.N.; Overturf, K.; Small, B.C. Insect (black soldier fly, Hermetia illucens) meal supplementation prevents the soybean meal-induced intestinal enteritis in rainbow trout and health benefits of using insect oil. Fish Shellfish Immunol. 2021, 109, 116–124. [Google Scholar] [CrossRef] [PubMed]
- St-Hilaire, S.; Sheppard, C.; Tomberlin, J.K.; Irving, S.; Newton, L.; McGuire, M.A.; Mosley, E.E.; Hardy, R.W.; Sealey, W. Fly prepupae as a feedstuff for rainbow trout, Oncorhynchus mykiss. J. World Aquac. Soc. 2007, 38, 59–67. [Google Scholar] [CrossRef]
- Fabrikov, D.; Barroso, F.G.; Sánchez-Muros, M.J.; Hidalgo, M.C.; Cardenete, G.; Tomás-Almenar, C.; Melenchón, F.; Guil-Guerrero, J.L. Effect of feeding with insect meal diet on the fatty acid compositions of sea bream (Sparus aurata), tench (Tinca tinca) and rainbow trout (Oncorhynchus mykiss) fillets. Aquaculture 2021, 545, 737170. [Google Scholar] [CrossRef]
- Shafique, L.; Abdel-Latif, H.; Hassan, F.-U.; Alagawany, M.; Naiel, M.; Dawood, M.; Yilmaz, S.; Liu, Q. The feasibility of using yellow mealworms (Tenebrio molitor): Towards a sustainable aquafeed industry. Animals 2021, 11, 811. [Google Scholar] [CrossRef]
- Jeong, S.M.; Khosravi, S.; Kim, K.-W.; Lee, B.-J.; Hur, S.-W.; You, S.-G.; Lee, S.-M. Potential of mealworm, Tenebrio molitor, meal as a sustainable dietary protein source for juvenile black porgy, Acanthopagrus schlegelii. Aquac. Rep. 2022, 22, 100956. [Google Scholar] [CrossRef]
- Ulbritch, T.L.V.; Southgate, D.A.T. Coronary heart disease: Seven dietary factors. Lancet 1991, 8773, 985–992. [Google Scholar] [CrossRef]
- Orkusz, A. Edible insects versus meat—Nutritional comparison: Knowledge of their composition is the key to good health. Nutrients 2021, 13, 1207. [Google Scholar] [CrossRef]
- Belghit, I.; Liland, N.S.; Gjesdal, P.; Biancarosa, I.; Menchetti, E.; Li, Y.; Waagbo, R.; Krogdahl, Å.; Lock, E.-J. Black soldier fly larvae meal can replace fish meal in diets of sea-water phase Atlantic salmon (Salmo salar). Aquaculture 2019, 503, 609–619. [Google Scholar] [CrossRef]
- Basto, A.; Calduch-Giner, J.; Oliveira, B.; Petit, L.; Sá, T.; Maia, M.R.G.; Fonseca, S.C.; Matos, E.; Pérez-Sánchez, J.; Valente, L.M.P. The use of defatted Tenebrio molitor larvae meal as a main protein source is supported in European sea bass (Dicentrarchus labrax) by data on growth performance, lipid metabolism, and flesh quality. Front. Physiol. 2021, 12, 659567. [Google Scholar] [CrossRef]
- Bordignon, F.; Gasco, L.; Birolo, M.; Trocino, A.; Christian, C.; Ballarin, C.; Bortoletti, M.; Nicoletto, C.; Maucieri, C.; Xiccato, G. Performance and fillet traits of rainbow trout (Oncorhynchus mykiss) fed different levels of Hermetia illucens meal in a low-tech aquaponic system. Aquaculture 2022, 546, 737279. [Google Scholar] [CrossRef]
- Sealey, W.M.; Gaylord, T.G.; Barrows, F.T.; Tomberlin, J.K.; McGuire, M.A.; Ross, C.; St-Hilaire, S. Sensory analysis of rainbow trout, Oncorhynchus mykiss, fed enriched black soldier fly prepupae, Hermetia illucens. J. World Aquac. Soc. 2011, 42, 34–45. [Google Scholar] [CrossRef]
- Barroso, F.G.; Sánchez-Muros, M.J.; Rincón, M.A.; Rodríguez-Rodríguez, M.; Fabrikov, D.; Morote, E.; Guil-Guerrero, J.L. Production of n-3-rich insects by bioaccumulation of fishery waste. J. Food Compos. Anal. 2019, 82, 103237. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Zimbelli, A.; Randazzo, B.; Compagni, M.D.; Truzzi, C.; Antonucci, M.; Riolo, P.; Loreto, N.; Osimani, A.; Milanović, V.; et al. Black Soldier Fly (Hermetia illucens) reared on roasted coffee by-product and Schizochytrium sp. as a sustainable terrestrial ingredient for aquafeeds production. Aquaculture 2020, 518, 734659. [Google Scholar] [CrossRef]
- Fabrikov, D.; Morote, E.; Montes, J.; Sánchez-Muros, M.J.; Barroso, F.G.; Rodríguez-Rodríguez, M.; González-Fernández, M.J.; Guil-Guerrero, J.L. Facing the challenge of discarded fish: Improving nutritional quality of two insect species larvae for use as feed and food. J. Insects Food Feed 2021, 7, 345–355. [Google Scholar] [CrossRef]
- Santigosa, E.; Brambilla, F.; Milanese, L. Microalgae oil as an effective alternative source of EPA and DHA for gilthead seabream (Sparus aurata) aquaculture. Animals 2021, 11, 971. [Google Scholar] [CrossRef]
- Santigosa, E.; Constant, D.; Prudence, D.; Wahli, T.; Verlhac-Trichet, V. A novel marine algal oil containing both EPA and DHA is an effective source of omega-3 fatty acids for rainbow trout (Oncorhynchus mykiss). JWAS 2020, 51, 649–665. [Google Scholar] [CrossRef]
- Li, S.; Ji, H.; Zhang, B.; Zhou, J.; Yu, H. Defatted black soldier fly (Hermetia illucens) larvae meal in diets for juvenile Jian carp (Cyprinus carpio var. Jian): Growth performance, antioxidant enzyme activities, digestive enzyme activities, intestine and hepatopancreas histological structure. Aquaculture 2017, 477, 62–70. [Google Scholar] [CrossRef]
- Su, J.; Gong, Y.; Cao, S.; Lu, F.; Han, D.; Liu, H.; Jin, J.; Yang, Y.; Zhu, X.; Xie, S. Effects of dietary Tenebrio molitor meal on the growth performance, immune response and disease resistance of yellow catfish (Pelteobagrus fulvidraco). Fish Shellfish Immunol. 2017, 69, 59–66. [Google Scholar] [CrossRef]
- Lu, R.; Chen, Y.; Yu, W.; Lin, M.; Yang, G.; Qin, C.; Meng, X.; Zhang, Y.; Ji, H.; Nie, G. Defatted black soldier fly (Hermetia illucens) larvae meal can replace soybean meal in juvenile grass carp (Ctenopharyngodon idellus) diets. Aquac. Rep. 2020, 18, 100520. [Google Scholar] [CrossRef]
- Hender, A.; Siddik, M.A.B.; Howieson, J.; Fotedar, R. Black soldier fly, Hermetia illucens as an alternative to fishmeal protein and fish oil: Impact on growth, immune response, mucosal barrier status, and flesh quality of juvenile barramundi, Lates calcarifer (Bloch, 1790). Biology 2021, 10, 505. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Randazzo, B.; Secci, G.; Notarstefano, V.; Giorgini, E.; Lock, E.J.; Parisi, G.; Olivotto, I. Application of laboratory methods for understanding fish responses to black soldier fly (Hermetia illucens) based diets. J. Insects Food Feed 2021, 8, 1173–1195. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Fish; National Academies Press: Washington, DC, USA, 1993. [Google Scholar]
- Blanco Cachafeiro, M.C. La Trucha: Cría Industrial, 2nd ed.; Mundi-Prensa: Madrid, Spain, 2005. [Google Scholar]
- AENOR UNE 173300; Pisciculture. Guide to Good Practice for Sacrifice. AENOR: Madrid, Spain, 2016.
- European Parliament. Book Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes; European Parliament: Strasbourg, France, 2010; pp. 33–78. [Google Scholar]
- Real Decreto 53/2013, de 1 de Febrero, por el que se Establecen las Normas Básicas Aplicables para la Protección de los Animales Utilizados en Experimentación y Otros Fines Científicos, Incluyendo la Docencia. 2013. Available online: https://www.boe.es/eli/es/rd/2013/02/01/53 (accessed on 27 January 2023).
- Rosenthal, A.J. Relation between instrumental and sensory measures of food texture. In Food Texture: Measurement and Perception; Aspen Publications: Boston, MA, USA, 1999. [Google Scholar]
- Skrede, G.; Storebakken, T.; Næs, T. Color evaluation in raw, baked and smoked flesh of rainbow trout (Onchorhynchus mykiss) fed astaxanthin or canthaxanthin. J. Food Sci. 1990, 55, 1574–1578. [Google Scholar] [CrossRef]
- Tomás-Almenar, C.; Larrán, A.M.; de Mercado, E.; Sanz-Calvo, M.A.; Hernández, D.; Riaño, B.; García-González, M.C. Scenedesmus Almeriensis from an integrated system waste-nutrient, as sustainable protein source for feed to rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 497, 422–430. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Communities International, 18th ed.; AOAC: Rockville, MD, USA, 2005. [Google Scholar]
- Saint-Denis, T.; Goupy, J. Optimization of a nitrogen analyser based on the Dumas method. Anal. Chim. Acta 2004, 515, 191–198. [Google Scholar] [CrossRef]
- Janssen, R.H.; Vincken, J.-P.; van den Broek, L.A.M.; Fogliano, V.; Lakemond, C.M.M. Nitrogen-to-protein conversion factors for three edible insects: Tenebrio Molitor, Alphitobius Diaperinus, and Hermetia Illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef]
- Gamage, A.; Shahidi, F. Use of chitosan for the removal of metal ion contaminants and proteins from water. Food Chem. 2007, 104, 989–996. [Google Scholar] [CrossRef]
- Pessoa, M.F.; Scotti-Campos, P.; Pais, I.; Feteiro, A.; Canuto, D.; Simões, M.; Pelica, J.; Pataco, I.; Ribeiro, V.; Reboredo, F.H.; et al. Nutritional profile of the Portuguese cabbage (Brassica Oleracea L. var. costata) and its relationship with the elemental soil analysis. Emir. J. Food Agric. 2016, 28, 381–388. [Google Scholar] [CrossRef]
- ISO 13730; Meat and Meat Products—Determination of Total Phosphorus Content—Spectrometric Method: ISO Technical Committee TC 34/SC 6. ISO: Geneva, Switzerland, 1996.
- Lepage, G.; Roy, C.C. Improved recovery of fatty acid through direct transesterification without prior extraction or purification. J. Lipid Res. 1984, 25, 1391–1396. [Google Scholar] [CrossRef]
- Rodríguez-Ruiz, J.; Belarbi, E.H.; Sánchez, J.L.G.; Alonso, D.L. Rapid simultaneous lipid extraction and transesterification for fatty acid analyses. Biotechnol. Tech. 1998, 12, 689–691. [Google Scholar] [CrossRef]
- Guil-Guerrero, J.L.; Gómez-Mercado, F.; Ramos-Bueno, R.P.; Rincón-Cervera, M.Á.; Venegas-Venegas, E. Restricted-range boraginaceae species constitute potential sources of valuable fatty acids. J. Am. Oil Chem. Soc. 2014, 91, 301–308. [Google Scholar] [CrossRef]
- Cladis, D.P.; Kleiner, A.C.; Freiser, H.H.; Santerre, C.R. Fatty acid profiles of commercially available finfish fillets in the United States. Lipids 2014, 49, 1005–1018. [Google Scholar] [CrossRef]
- García-Márquez, J.; Galafat, A.; Vizcaíno, A.J.; Barany, A.; Martos-Sitcha, J.A.; Mancera, J.M.; Acién, G.; Figueroa, F.L.; Alarcón, F.J.; Arijo, S.; et al. Dietary use of the microalga Chlorella fusca Improves growth, metabolism, and digestive functionality in thick-lipped grey mullet (Chelon labrosus, Risso 1827) juveniles. Front. Mar. Sci. 2022, 9, 902203. [Google Scholar] [CrossRef]
- Furné, M.; Morales, A.E.; Trenzado, C.E.; García-Gallego, M.; Hidalgo, M.C.; Domezain, A.; Rus, A.S. The metabolic effects of prolonged starvation and refeeding in sturgeon and rainbow trout. J. Comp. Physiol. B 2012, 182, 63–76. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Pérez-Jiménez, A.; Hidalgo, M.C.; Morales, A.E.; Arizcun, M.; Abellán, E.; Cardenete, G. Antioxidant enzymatic defenses and oxidative damage in Dentex dentex fed on different dietary macronutrient levels. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2009, 150, 537–545. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for statistical computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 27 April 2023).
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio, PBC: Boston, MA, USA, 2022; Available online: http://www.rstudio.com/ (accessed on 27 April 2023).
- Liland, N.S.; Araujo, P.; Xu, X.X.; Lock, E.-J.; Radhakrishnan, G.; Prabhu, A.J.P.; Belghit, I. A meta-analysis on the nutritional value of insects in aquafeeds. J. Insects Food Feed 2021, 7, 743–759. [Google Scholar] [CrossRef]
- Weththasinghe, P.; Hansen, J.Ø.; Mydland, L.T.; Øverland, M. A systematic meta-analysis based review on black soldier fly (Hermetia illucens) as a novel protein source for salmonids. Rev. Aquac. 2021, 14, 938–956. [Google Scholar] [CrossRef]
- Rema, P.; Saravanan, S.; Armenjon, B.; Motte, C.; Dias, J. Graded incorporation of defatted yellow mealworm (Tenebrio molitor) in rainbow trout (Oncorhynchus mykiss) diet improves growth performance and nutrient retention. Animals 2019, 9, 187. [Google Scholar] [CrossRef]
- Guerreiro, I.; Castro, C.; Antunes, B.; Coutinho, F.; Rangel, F.; Couto, A.; Serra, C.R.; Peres, H.; Pousão-Ferreira, P.; Matos, E.; et al. Catching black soldier fly for meagre: Growth, whole-body fatty acid profile and metabolic responses. Aquaculture 2020, 516, 734613. [Google Scholar] [CrossRef]
- Coutinho, F.; Castro, C.; Guerreiro, I.; Rangel, F.; Couto, A.; Serra, C.R.; Peres, H.; Pousão-Ferreira, P.; Rawski, M.; Oliva-Teles, A.; et al. Mealworm larvae meal in diets for meagre juveniles: Growth, nutrient digestibility and digestive enzymes activity. Aquaculture 2021, 535, 736362. [Google Scholar] [CrossRef]
- Estévez, A.; Blanco, B.; Fernández, L.; Ferreira, M.; Soula, M. Effects of alternative and sustainable ingredients, insect meal, microalgae and protein and lipid from tuna cooking water, on meagre (Argyrosomus regius) growth, food conversion and muscle and liver composition. Aquaculture 2022, 548, 737549. [Google Scholar] [CrossRef]
- Taufek, N.M.; Aspani, F.; Muin, H.; Raji, A.A.; Razak, S.A.; Alias, Z. The effect of dietary cricket meal (Gryllus bimaculatus) on growth performance, antioxidant enzyme activities, and haematological response of African catfish (Clarias gariepinus). Fish Physiol. Biochem. 2016, 42, 1143–1155. [Google Scholar] [CrossRef] [PubMed]
- Rapatsa, M.; Moyo, N.A.G. Evaluation of Imbrasia belina meal as a fishmeal substitute in Oreochromis mossambicus diets: Growth performance, histological analysis and enzyme activity. Aquac. Rep. 2017, 5, 18–26. [Google Scholar] [CrossRef]
- Sankian, Z.; Khosravi, S.; Kim, Y.-O.; Lee, S.-M. Effects of dietary inclusion of yellow mealworm (Tenebrio molitor) meal on growth performance, feed utilization, body composition, plasma biochemical indices, selected immune parameters and antioxidant enzyme activities of mandarin fish (Siniperca scherzeri) juveniles. Aquaculture 2018, 496, 79–87. [Google Scholar] [CrossRef]
- Vongvichith, B.; Morioka, S.; Sugita, T.; Phousavanh, N.; Phetsanghanh, N.; Chantasone, P.; Pommachan, P.; Nakamura, S. Evaluation of the efficacy of aquaculture feeds for the climbing perch Anabas testudineus: Replacement of fishmeal by black soldier fly Hermetia illucens prepupae. Fish. Sci. 2020, 86, 145–151. [Google Scholar] [CrossRef]
- Agbohessou, P.S.; Mandiki, S.N.M.; Gougbédji, A.; Megido, R.C.; Hossain, M.S.; de Jaeger, P.; Larondelle, Y.; Francis, F.; Lalèye, P.A.; Kestemont, P. Total replacement of fish meal by enriched-fatty acid Hermetia illucens meal did not substantially affect growth parameters or innate immune status and improved whole body biochemical quality of Nile tilapia juveniles. Aquac. Nutr. 2021, 27, 880–896. [Google Scholar] [CrossRef]
- Gebremichael, A.; Sándor, Z.J.; Kucska, B. Does dietary inclusion of defatted yellow mealworm (Tenebrio molitor) affect growth and body composition of juvenile common carp (Cyprinus carpio)? S. Afr. J. Anim. Sci. 2022, 52, 444–451. [Google Scholar] [CrossRef]
- Mastoraki, M.; Ferrándiz, P.M.; Vardali, S.C.; Kontodimas, D.C.; Kotzamanis, Y.P.; Gasco, L.; Chatzifotis, S.; Antonopoulou, E. A comparative study on the effect of fish meal substitution with three different insect meals on growth, body composition and metabolism of European sea bass (Dicentrarchus labrax L.). Aquaculture 2020, 528, 735511. [Google Scholar] [CrossRef]
- Jeong, S.-M.; Khosravi, S.; Yoon, K.-Y.; Kim, K.-W.; Lee, B.-J.; Hur, S.-W.; Lee, S.-M. Mealworm, Tenebrio molitor, as a feed ingredient for juvenile olive flounder, Paralichthys olivaceus. Aquac. Rep. 2021, 20, 100747. [Google Scholar] [CrossRef]
- Mancini, S.; Medina, I.; Iaconisi, V.; Gai, F.; Basto, A.; Parisi, G. Impact of black soldier fly larvae meal on the chemical and nutritional characteristics of rainbow trout fillets. Animal 2018, 12, 1672–1681. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.S.; Fawole, F.J.; Labh, S.N.; Small, B.C.; Overturf, K.; Kumar, V. Insect meal inclusion as a novel feed ingredient in soy-based diets improves performance of rainbow trout (Oncorhynchus mykiss). Aquaculture 2021, 544, 737096. [Google Scholar] [CrossRef]
- Turchini, G.M.; Francis, D.S.; Senadheera, S.P.S.D.; Thanuthong, T.; de Silva, S.S. Fish oil replacement with different vegetable oils in Murray cod: Evidence of an “omega-3 sparing effect” by other dietary fatty acids. Aquaculture 2011, 315, 250–259. [Google Scholar] [CrossRef]
- Emery, J.A.; Norambuena, F.; Trushenski, J.; Turchini, G.M. Uncoupling EPA and DHA in fish nutrition: Dietary demand is limited in Atlantic salmon and effectively met by DHA alone. Lipids 2016, 51, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Turchini, G.; Francis, D.S.; Liang, M.; Mock, T.S.; Rombenso, A.; Ai, Q. Are fish what they eat? A fatty acid’s perspective. Prog. Lipid Res. 2020, 80, 101064. [Google Scholar] [CrossRef]
- Piedecausa, M.A.; Mazón, M.J.; García, B.G.; Hernández, M.D. Effects of total replacement of fish oil by vegetable oils in the diets of sharpsnout seabream (Diplodus puntazzo). Aquaculture 2007, 263, 211–219. [Google Scholar] [CrossRef]
- Karalazos, V.; Bendiksen, E.Å.; Bell, J.G. Interactive effects of dietary protein/lipid level and oil source on growth, feed utilisation and nutrient and fatty acid digestibility of Atlantic salmon. Aquaculture 2011, 311, 193–200. [Google Scholar] [CrossRef]
- Mozanzadeh, M.T.; Agh, N.; Yavari, V.; Marammazi, J.G.; Mohammadian, T.; Gisbert, E. Partial or total replacement of dietary fish oil with alternative lipid sources in silvery-black porgy (Sparidentex hasta). Aquaculture 2016, 451, 232–240. [Google Scholar] [CrossRef]
- Tocher, D.R. Fatty acid requirements in ontogeny of marine and freshwater fish. Aquac. Res. 2010, 41, 717–732. [Google Scholar] [CrossRef]
- Tocher, D.R. Omega-3 long-chain polyunsaturated fatty acids and aquaculture in perspective. Aquaculture 2015, 449, 94–107. [Google Scholar] [CrossRef]
- Gause, B.R.; Trushenski, J.T. Sparing fish oil with beef tallow in feeds for rainbow trout: Effects of inclusion rates and finishing on production performance and tissue fatty acid composition. N. Am. J. Aquac. 2013, 75, 495–511. [Google Scholar] [CrossRef]
- Asghari, M.; Shabanpour, B.; Pakravan, S. Evaluation of some qualitative variations in frozen fillets of beluga (Huso huso) fed by different carbohydrate to lipid ratios. J. Food Sci. Technol. 2014, 51, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Pavón, Y.; Cian, R.E.; Soldini, M.A.C.; Hernández, D.R.; Sánchez, S.; Drago, S.R. Sensory and instrumental textural changes in fillets from Pacú (Piaractus mesopotamicus) fed different diets. J. Texture Stud. 2018, 49, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Chaiyapechara, S.; Liu, K.K.M.; Barrows, F.T.; Hardy, R.W.; Dong, F.M. Proximate composition, lipid oxidation, and sensory characteristics of fillets from rainbow trout Oncorhynchus mykiss fed diets containing 10% to 30% lipid. J. World Aquac. Soc. 2007, 34, 266–277. [Google Scholar] [CrossRef]
- Stadtlander, T.; Stamer, A.; Buser, A.; Wohlfahrt, J.; Leiber, F.; Sandrock, C. Hermetia illucens meal as fish meal replacement for rainbow trout on farm. J. Insects Food Feed 2017, 3, 165–175. [Google Scholar] [CrossRef]
- Bruni, L.; Randazzo, B.; Cardinaletti, G.; Zarantoniello, M.; Mina, F.; Secci, G.; Tulli, F.; Olivotto, I.; Parisi, G. Dietary inclusion of full-fat Hermetia illucens prepupae meal in practical diets for rainbow trout (Oncorhynchus mykiss): Lipid metabolism and fillet quality investigations. Aquaculture 2020, 529, 735678. [Google Scholar] [CrossRef]
- Iaconisi, V.; Bonelli, A.; Pupino, R.; Gai, F.; Parisi, G. Mealworm as dietary protein source for rainbow trout: Body and fillet quality traits. Aquaculture 2018, 484, 197–204. [Google Scholar] [CrossRef]
- Iaconisi, V.; Marono, S.; Parisi, G.; Gasco, L.; Genovese, L.; Maricchiolo, G.; Bovera, F.; Piccolo, G. Dietary inclusion of Tenebrio molitor larvae meal: Effects on growth performance and final quality treats of blackspot sea bream (Pagellus bogaraveo). Aquaculture 2017, 476, 49–58. [Google Scholar] [CrossRef]
- Borgogno, M.; Dinnella, C.; Iaconisi, V.; Fusi, R.; Scarpaleggia, C.; Schiavone, A.; Monteleone, E.; Gasco, L.; Parisi, G. Inclusion of Hermetia illucens larvae meal on rainbow trout (Oncorhynchus mykiss) feed: Effect on sensory profile according to static and dynamic evaluations. J. Sci. Food Agric. 2017, 97, 3402–3411. [Google Scholar] [CrossRef]
- Finke, M.D. Complete nutrient composition of commercially raised invertebrates used as food for insectivores. Zoo Biol. 2002, 21, 269–285. [Google Scholar] [CrossRef]
- Hoffmann, L.; Rawski, M.; Nogales-Merida, S.; Mazurkiewicz, J. Dietary inclusion of Tenebrio molitor meal in sea trout larvae rearing: Effects on fish growth performance, survival, condition, and GIT and liver enzymatic activity. Ann. Anim. Sci. 2020, 20, 579–598. [Google Scholar] [CrossRef]
- Nogueira, M.S.; Scolaro, B.; Milne, G.L.; Castro, I.A. Oxidation products from omega-3 and omega-6 fatty acids during a simulated shelf life of edible oils. LWT—Food Sci. Technol. 2019, 101, 113–122. [Google Scholar] [CrossRef]
- Secci, G.; Mancini, S.; Iaconisi, V.; Gasco, L.; Basto, A.; Parisi, G. Can the inclusion of black soldier fly (Hermetia illucens) in diet affect the flesh quality/nutritional traits of rainbow trout (Oncorhynchus mykiss) after freezing and cooking? Int. J. Food Sci. Nutr. 2019, 70, 161–171. [Google Scholar] [CrossRef]
- Hidalgo, M.C.; Morales, A.E.; Pula, H.J.; Tomás-Almenar, C.; Sánchez-Muros, M.J.; Melenchón, F.; Fabrikov, D.; Cardenete, G. Oxidative metabolism of gut and innate immune status in skin and blood of tench (Tinca tinca) fed with different insect meals (Hermetia illucens and Tenebrio molitor). Aquaculture 2022, 529, 735731. [Google Scholar] [CrossRef]
- Sreejai, R.; Jaya, D.S. Studies on the changes in lipid peroxidation and antioxidants in fishes exposed to hydrogen sulfide. Toxicol. Int. 2010, 17, 71–77. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Proximate Composition | Fishmeal LT94 | T. molitor | Defatted T. molitor |
|---|---|---|---|
| Moisture (%) | 5.00 | 6.50 | 2.60 |
| Crude protein (%) | 69.40 | 52.80 | 70.70 |
| Crude lipid (%) | 12.30 | 28.50 | 8.90 |
| Chitin (%) | - | 3.20 | 5.50 |
| Ash (%) | 16.00 | 2.91 | 3.96 |
| Calcium (g/kg) | 38.00 | 0.51 | 1.10 |
| Phosphorus (g/kg) | 26.00 | 6.50 | 8.20 |
| Calcium:Phosphorus ratio | 1.46 | 0.08 | 0.13 |
| Amino acid composition (g/100 g insect meal) | |||
| Essential amino acids | |||
| Arginine | 3.48 | 2.59 | 3.39 |
| Histidine | 1.23 | 1.60 | 2.22 |
| Isoleucine | 2.90 | 2.18 | 2.92 |
| Leucine | 4.50 | 3.55 | 4.82 |
| Lysine | 5.95 | 3.44 | 4.68 |
| Methionine | 1.68 | 0.68 | 0.93 |
| Phenylalanine | 2.93 | 2.10 | 3.18 |
| Threonine | 3.21 | 1.85 | 2.54 |
| Valine | 5.24 | 3.40 | 4.32 |
| Non-essential amino acids | |||
| Alanine | 5.20 | 3.62 | 4.70 |
| Aspartate | 7.86 | 4.03 | 5.53 |
| Cysteine | 0.50 | 0.40 | 0.43 |
| Glutamate | 9.09 | 5.52 | 7.48 |
| Glycine | 3.60 | 2.69 | 3.46 |
| Proline | 3.61 | 3.22 | 4.05 |
| Serine | 2.78 | 2.09 | 2.84 |
| Tyrosine | 2.35 | 5.89 | 8.06 |
| (a) | |||||
|---|---|---|---|---|---|
| Ingredients (%; On Dry Basis) | C | T | dT | TO1 | TO2 |
| Fishmeal LT94 1 | 36.78 | 18.28 | 18.48 | 18.28 | 18.28 |
| TM meal 2 | - | 19.05 | - | 19.05 | 19.05 |
| Defatted TM meal 3 | - | - | 18.29 | - | - |
| Enriched omega−3 oil (EO) | - | - | - | 3.09 | 7.24 |
| Wheat gluten 4 | 11.05 | 12.41 | 10.65 | 12.41 | 12.41 |
| Soybean protein concentrate 5 | 15.09 | 16.40 | 15.41 | 16.40 | 16.40 |
| Wheat meal 6 | 16.16 | 16.98 | 16.19 | 16.98 | 16.98 |
| Soybean lecithin 7 | 1.27 | 0.48 | 1.27 | 0.48 | 0.48 |
| Fish oil 8 | 11.87 | 8.68 | 11.90 | 5.60 | 1.45 |
| Vitamin and mineral premix 9 | 1.95 | 1.93 | 1.95 | 1.93 | 1.93 |
| Goma guar 10 | 1.95 | 1.93 | 1.95 | 1.93 | 1.93 |
| Blood meal 10 | 3.89 | 3.86 | 3.90 | 3.86 | 3.86 |
| Methionine 10 | 0.20 | 0.50 | 0.50 | 0.50 | 0.50 |
| Lysine 10 | - | 0.10 | 0.10 | 0.10 | 0.10 |
| Proximate Composition (%; On Dry Basis) | C | T | dT | TO1 | TO2 |
| Crude protein | 49.19 | 48.76 | 48.98 | 49.25 | 48.23 |
| Crude lipid | 17.80 | 18.58 | 17.86 | 18.99 | 19.12 |
| Crude fibre | 0.97 | 2.15 | 2.35 | 2.25 | 2.47 |
| Ash | 8.63 | 8.49 | 6.31 | 6.22 | 6.12 |
| Calcium | 0.43 | 0.24 | 0.17 | 0.18 | 0.20 |
| Phosphorus | 0.31 | 0.24 | 0.24 | 0.24 | 0.24 |
| Calcium:Phosphorus ratio | 1.38 | 1.00 | 0.73 | 0.77 | 0.86 |
| (b) | |||||
| Amino Acid Composition (g/100 g Wet Feed) | C | T | dT | TO1 | TO2 |
| Essential amino acids | |||||
| Arginine | 2.10 | 2.35 | 2.08 | 2.01 | 2.00 |
| Histidine | 1.09 | 1.02 | 1.05 | 1.05 | 1.04 |
| Isoleucine | 1.50 | 1.69 | 1.45 | 1.56 | 1.44 |
| Leucine | 3.00 | 2.99 | 3.02 | 2.90 | 2.88 |
| Lysine | 2.94 | 3.66 | 2.77 | 2.70 | 2.66 |
| Methionine | 1.01 | 1.15 | 1.23 | 1.18 | 1.13 |
| Phenylalanine | 2.07 | 2.19 | 2.01 | 1.96 | 1.95 |
| Threonine | 1.48 | 1.56 | 1.48 | 1.31 | 1.36 |
| Valine | 2.10 | 2.07 | 2.16 | 2.25 | 2.10 |
| Non-essential amino acids | |||||
| Alanine | 2.01 | 2.06 | 2.18 | 1.95 | 1.94 |
| Aspartate | 3.32 | 3.56 | 3.35 | 3.10 | 3.12 |
| Cysteine | 0.46 | 0.66 | 0.46 | 0.43 | 0.44 |
| Glutamate | 7.15 | 7.64 | 7.41 | 7.44 | 7.44 |
| Glycine | 1.92 | 2.20 | 1.91 | 1.81 | 1.80 |
| Proline | 2.46 | 2.51 | 2.67 | 2.74 | 2.61 |
| Serine | 1.76 | 1.87 | 1.91 | 1.65 | 1.79 |
| Tyrosine | 1.63 | 1.78 | 2.94 | 2.12 | 2.42 |
| Mineral Composition (g/100 g Wet Feed) | C | T | dT | TO1 | TO2 |
| Sodium | 0.25 | 0.14 | 0.14 | 0.13 | 0.14 |
| Magnesium | 0.07 | 0.08 | 0.08 | 0.08 | 0.07 |
| Silicon | 0.11 | 0.11 | 0.09 | 0.10 | 0.13 |
| Phosphorus | 0.29 | 0.22 | 0.22 | 0.22 | 0.22 |
| Sulfur | 0.20 | 0.21 | 0.20 | 0.22 | 0.21 |
| Chlorine | 0.34 | 0.26 | 0.23 | 0.25 | 0.25 |
| Potassium | 0.34 | 0.38 | 0.44 | 0.46 | 0.45 |
| Calcium | 0.40 | 0.22 | 0.16 | 0.17 | 0.19 |
| (c) | |||||
| Fatty Acid Composition (g/100 Total FA) | C | T | dT | TO1 | TO2 |
| C14:0 | 3.68 | 3.26 | 2.55 | 3.16 | 3.20 |
| C16:0 | 21.13 | 19.37 | 17.35 | 20.75 | 22.74 |
| C16:1n−7 | 4.68 | 2.97 | 3.56 | 3.02 | 2.00 |
| C18:0 | 5.67 | 4.78 | 4.96 | 3.95 | 2.87 |
| C18:1n−9 | 13.67 | 23.05 | 16.17 | 20.47 | 17.14 |
| C18:1n−7 | 2.60 | 1.54 | 1.42 | 0.00 | 0.00 |
| C18:2n−6 | 8.77 | 18.69 | 13.75 | 15.31 | 15.21 |
| C18:3n−3 | 1.12 | 1.33 | 1.56 | 0.95 | 0.87 |
| C18:4n−3 | 0.82 | 0.44 | 0.89 | 0.61 | 0.51 |
| C20:1n−9 | 0.98 | 0.76 | 1.75 | 1.14 | 0.56 |
| C20:4n−6 | 1.66 | 1.22 | 1.56 | 1.17 | 1.26 |
| C20:5n−3 | 8.31 | 3.92 | 6.42 | 6.33 | 8.51 |
| C22:5n−3 | 1.33 | 0.89 | 1.49 | 1.08 | 1.12 |
| C22:6n−3 | 18.79 | 14.05 | 19.07 | 16.10 | 19.20 |
| Other (up to 100%) | 6.79 | 3.73 | 7.50 | 5.96 | 4.81 |
| ∑Saturated fatty acids (SFA) | 30.48 | 27.41 | 24.87 | 27.86 | 28.81 |
| ∑Monounsaturated fatty acids (MUFA) | 21.93 | 28.32 | 22.91 | 24.63 | 19.70 |
| ∑Polyunsaturated fatty acids (PUFA) | 40.80 | 40.54 | 44.73 | 41.56 | 46.67 |
| ∑Omega−3 (n−3) | 30.37 | 20.63 | 29.42 | 25.07 | 30.20 |
| ∑Omega−6 (n−6) | 10.43 | 19.91 | 15.31 | 16.49 | 16.47 |
| ∑Omega−6/∑Omega−3 (n−6/n−3) | 0.34 | 0.97 | 0.52 | 0.66 | 0.55 |
| ∑Total fatty acids | 100 | 100 | 100 | 100 | 100 |
| Growth Performance | C | T | dT | TO1 | TO2 | SEM |
|---|---|---|---|---|---|---|
| IBW (g) | 46.11 | 46.25 | 46.08 | 46.20 | 46.05 | 0.11 |
| Initial length (cm) | 15.93 | 16.00 | 15.88 | 15.95 | 15.95 | 0.05 |
| FBW (g) | 417.13 | 413.9 | 417.38 | 417.72 | 411.09 | 3.49 |
| Final length (cm) | 29.94 | 29.86 | 30.06 | 30.01 | 29.84 | 0.12 |
| SGR (%/day) | 2.47 | 2.46 | 2.48 | 2.47 | 2.46 | 0.01 |
| DFI (g/100 g fish·day) | 1.50 | 1.48 | 1.49 | 1.48 | 1.48 | 0.01 |
| FCR | 0.86 | 0.85 | 0.86 | 0.85 | 0.85 | 0.005 |
| CF (g/cm3) | 1.55 | 1.55 | 1.54 | 1.55 | 1.55 | 0.02 |
| Proximate Composition (Wet Basis) | C | T | dT | TO1 | TO2 | SEM |
|---|---|---|---|---|---|---|
| Moisture (%) | 73.55 a | 71.86 b | 72.95 ab | 72.53 ab | 73.03 ab | 0.31 |
| Protein (%) | 20.29 | 20.69 | 20.39 | 20.28 | 20.11 | 0.27 |
| Fat (%) | 1.14 b | 3.06 a | 1.88 ab | 3.16 a | 2.27 ab | 0.41 |
| Ash (%) | 1.30 | 1.32 | 1.31 | 1.33 | 1.31 | 0.02 |
| Calcium (mg/kg) | 182.00 | 162.13 | 164.63 | 147.25 | 126.38 | 12.41 |
| Phosphorus (g/kg) | 2.58 | 2.60 | 2.53 | 2.62 | 2.45 | 0.02 |
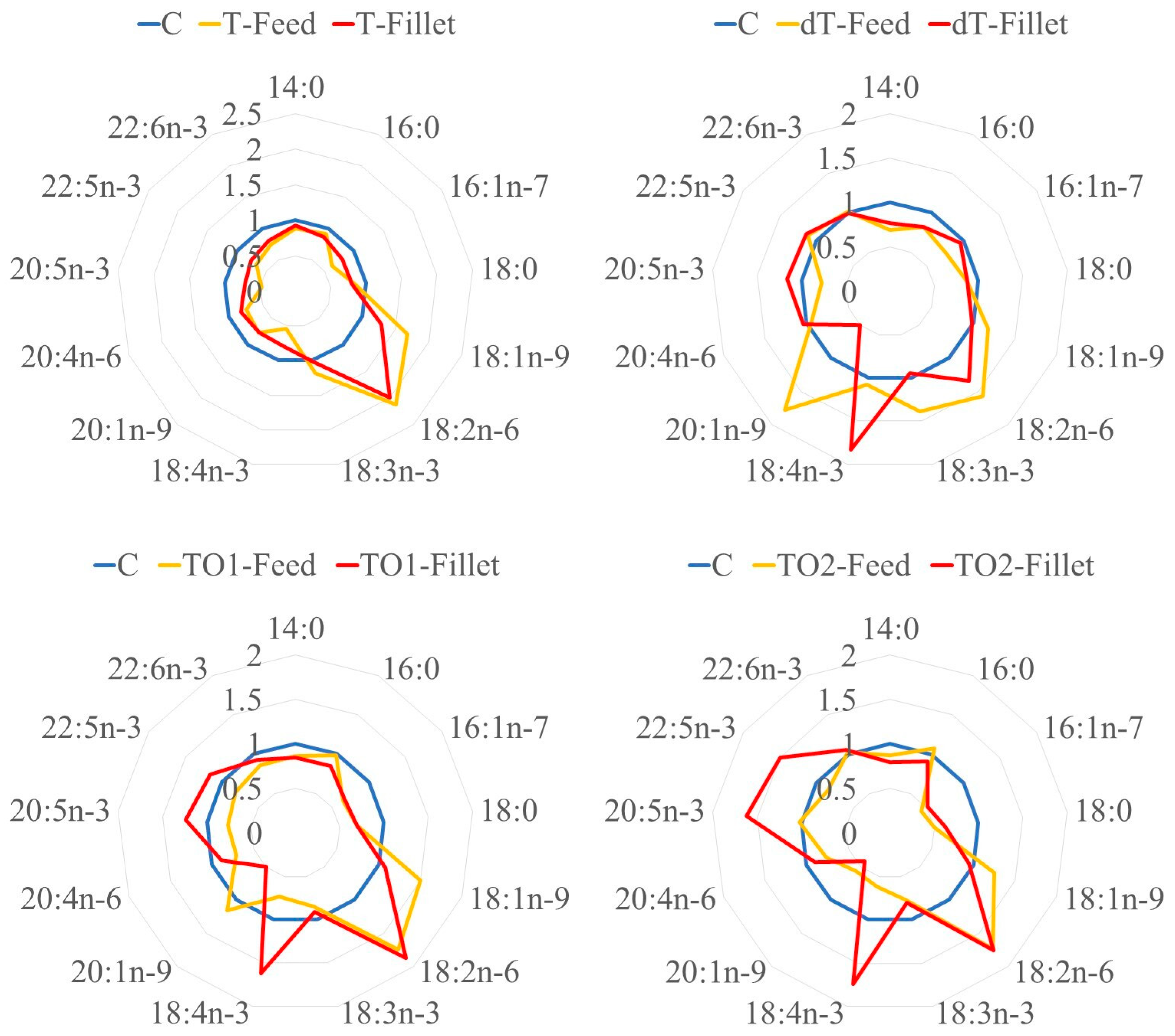
| Fatty Acids Profile of Fillets (% of Total FA) | C | T | dT | TO1 | TO2 | SEM |
|---|---|---|---|---|---|---|
| C14:0 | 3.06 a | 2.84 ab | 2.36 c | 2.60 bc | 2.45 c | 0.07 |
| C16:0 | 23.20 a | 20.09 bc | 18.97 c | 19.59 c | 21.08 b | 0.28 |
| C16:1n−7 | 4.36 a | 3.48 b | 4.18 a | 2.93 c | 2.26 d | 0.12 |
| C18:0 | 6.28 a | 5.05 c | 5.53 b | 4.37 d | 3.92 e | 0.09 |
| C18:1n−9 | 19.54 bc | 25.16 a | 19.28 bc | 21.15 b | 18.60 c | 0.46 |
| C18:1n−7 | 2.89 a | 1.87 c | 2.09 b | 1.35 d | 0.99 e | 0.03 |
| C18:2n−6 | 7.13 e | 14.19 a | 9.57 d | 13.36 b | 12.54 c | 0.14 |
| C18:3n−3 | 0.97 ab | 0.99 a | 0.92 bc | 0.88 c | 0.78 d | 0.01 |
| C18:4n−3 | 0.31 | 0.24 | 0.57 | 0.50 | 0.54 | 0.08 |
| C20:1n−9 | 1.48 a | 1.14 b | 0.75 c | 0.74 c | 0.63 c | 0.06 |
| C20:4n−6 | 1.29 a | 1.05 b | 1.34 a | 1.14 ab | 1.17 ab | 0.05 |
| C20:4n−3 | 0.29 b | 0.00 c | 0.62 a | 0.41 ab | 0.36 b | 0.05 |
| C20:5n−3 | 3.29 c | 2.34 d | 3.83 bc | 4.09 b | 5.33 a | 0.15 |
| C22:5n−3 | 1.13 c | 0.86 d | 1.29 b | 1.31 b | 1.69 a | 0.03 |
| C22:6n−3 | 18.95 a | 15.22 b | 18.86 a | 17.51 ab | 19.94 a | 0.69 |
| Other (up to 100%) | 5.82 | 5.47 | 9.84 | 8.07 | 7.72 | 0.43 |
| ∑Saturated fatty acids (SFA) | 32.53 a | 27.98 b | 26.85 b | 26.56 b | 27.44 b | 0.40 |
| ∑Monounsaturated fatty acids (MUFA) | 28.28 b | 31.65 a | 26.31 b | 26.16 b | 22.47 c | 0.62 |
| ∑Polyunsaturated fatty acids (PUFA) | 33.38 c | 34.90 c | 37.00 bc | 39.21 ab | 42.36 a | 0.91 |
| ∑Omega−3 (n−3) | 24.95 b | 19.65 c | 26.08 ab | 24.71 b | 28.65 a | 0.85 |
| ∑Omega−6 (n−6) | 8.42 e | 15.24 a | 10.91 d | 14.50 b | 13.71 c | 0.15 |
| ∑Omega−6/∑Omega−3 (n−6/n−3) | 0.34 d | 0.78 a | 0.42 cd | 0.59 b | 0.48 c | 0.02 |
| Peroxidability index (PI) | 195.51 b | 163.09 c | 203.75 ab | 196.46 ab | 224.63 a | 6.50 |
| Texture Parameters in Raw Fillets | C | T | dT | TO1 | TO2 | SEM |
|---|---|---|---|---|---|---|
| Brittleness | 64.13 | 58.76 | 71.32 | 78.60 | 61.16 | 7.26 |
| Hardness | 182.34 | 144.48 | 156.93 | 179.01 | 121.70 | 23.49 |
| Cohesiveness | 0.44 | 0.49 | 0.42 | 0.40 | 0.45 | 0.03 |
| Adhesiveness | −3.78 | −3.88 | −3.01 | −2.82 | −3.96 | 0.36 |
| Elasticity | 1.00 | 1.00 | 1.01 | 1.02 | 1.01 | 0.01 |
| Chewiness | 79.65 | 72.47 | 66.74 | 73.70 | 54.25 | 11.93 |
| Gumminess | 79.58 | 72.37 | 66.30 | 72.46 | 53.73 | 11.96 |
| Colour Parameters in Raw Fillets | C | T | dT | TO1 | TO2 | SEM |
|---|---|---|---|---|---|---|
| Lightness (L*) | 58.08 ab | 57.16 b | 59.05 ab | 59.55 a | 58.95 ab | 0.54 |
| Redness (a*) | −1.57 a | −1.74 ab | −1.80 ab | −1.74 ab | −2.08 b | 0.09 |
| Yellowness (b*) | 7.64 | 6.48 | 5.84 | 6.90 | 7.05 | 0.52 |
| Chroma (C*ab) | 7.83 | 6.74 | 6.13 | 7.13 | 7.38 | 0.48 |
| Hue (H°ab) | 102.91 | 106.04 | 107.82 | 104.35 | 106.14 | 1.68 |
| Attributes in Raw Fillets | C | T | dT | TO1 | TO2 | SEM |
|---|---|---|---|---|---|---|
| Acceptability | 2.87 | 2.70 | 2.78 | 1.97 | 2.40 | 0.19 |
| Colour | 5.37 | 5.09 | 5.50 | 5.99 | 5.48 | 0.27 |
| Texture | 2.74 | 1.98 | 2.16 | 1.87 | 1.85 | 0.20 |
| Odour | 2.75 | 2.68 | 2.55 | 2.67 | 2.70 | 0.43 |
| Quality Index (QI) | 3.12 | 3.77 | 4.00 | 3.85 | 4.69 | 0.42 |
| Attributes in Cooked Fillets | C | T | dT | TO1 | TO2 | SEM |
|---|---|---|---|---|---|---|
| Appearance | ||||||
| Brightness | 9.04 a | 9.03 a | 8.68 ab | 8.93 ab | 7.27 b | 0.38 |
| Exudate colour | 7.48 | 7.53 | 8.06 | 7.88 | 8.17 | 0.66 |
| Colour uniformity | 7.84 | 7.86 | 6.88 | 7.06 | 6.31 | 0.67 |
| Colour intensity | 6.83 | 7.03 | 5.18 | 5.23 | 3.86 | 1.00 |
| Odour intensity | 6.58 | 5.95 | 5.05 | 4.70 | 5.88 | 0.53 |
| Taste | ||||||
| Sweet | 2.78 | 1.56 | 1.17 | 2.58 | 1.29 | 0.61 |
| Salty | 2.12 | 2.08 | 2.11 | 1.81 | 2.65 | 0.35 |
| Bitter | 0.73 | 1.44 | 1.50 | 1.11 | 1.69 | 0.43 |
| Acid | 0.40 | 0.49 | 0.92 | 0.64 | 0.60 | 0.35 |
| Texture | ||||||
| Hardness | 2.82 | 2.40 | 3.40 | 2.01 | 3.28 | 0.61 |
| Juiciness | 5.59 | 4.90 | 4.10 | 6.22 | 3.93 | 0.61 |
| Fatty | 2.24 | 3.28 | 2.75 | 2.60 | 2.15 | 0.48 |
| Elastic/gumminess | 4.14 | 3.22 | 4.80 | 3.57 | 3.77 | 0.57 |
| Teeth adherence | 3.82 | 2.94 | 4.56 | 3.82 | 4.23 | 0.55 |
| Flavor | ||||||
| Rancid | 1.18 | 1.35 | 1.22 | 1.05 | 1.64 | 0.31 |
| Vegetable | 1.57 | 0.93 | 0.99 | 0.88 | 0.89 | 0.23 |
| Mud/earthy | 3.28 | 3.66 | 2.92 | 2.58 | 3.24 | 0.42 |
| Sea | 3.07 | 2.82 | 2.43 | 2.70 | 3.18 | 0.35 |
| Fatty sensation | 1.70 | 2.01 | 2.04 | 2.15 | 1.44 | 0.47 |
| Liver Intermediary Metabolism Enzymes | C | T | dT | TO1 | TO2 | SEM |
|---|---|---|---|---|---|---|
| FBPase (mU/mg prot) | 80.20 | 85.22 | 62.57 | 62.41 | 83.08 | 6.60 |
| PK (mU/mg prot) | 399.91 | 740.88 | 913.83 | 817.13 | 774.73 | 115.64 |
| GPT (mU/mg prot) | 539.02 | 556.58 | 486.27 | 529.78 | 534.36 | 57.53 |
| GOT (mU/mg prot) | 3310.83 | 3049.77 | 2731.50 | 2377.50 | 2391.62 | 267.35 |
| GDH (mU/mg prot) | 851.22 | 738.61 | 652.25 | 683.12 | 758.58 | 60.79 |
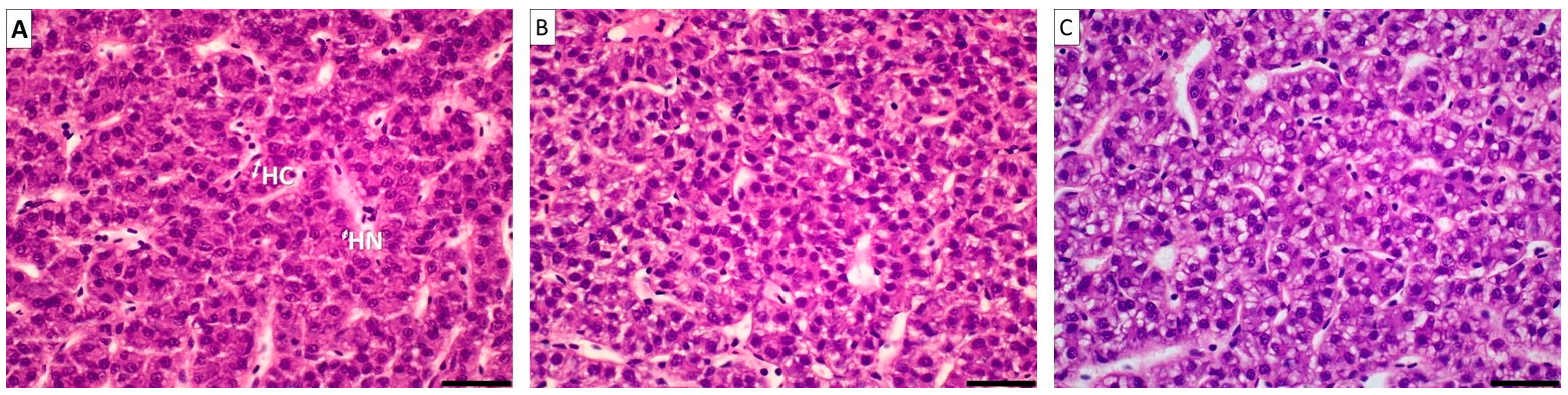
| Liver Histomorphology (μm) | C | T | dT | TO1 | TO2 | SEM |
|---|---|---|---|---|---|---|
| Hepatocyte nucleus width | 2.05 | 2.04 | 2.01 | 2.09 | 2.10 | 0.04 |
| Hepatocyte cytoplasm width | 4.45 | 4.42 | 4.51 | 4.59 | 4.62 | 0.05 |
| Lipid accumulation in hepatocytes | + | + | ++ | ++ | ++ | |
| Presence of necrotic focuses | − | − | − | − | − |
| Liver Antioxidant Enzymes and Lipid Peroxidation | C | T | dT | TO1 | TO2 | SEM |
|---|---|---|---|---|---|---|
| SOD (U/mg prot.) | 282.36 | 271.14 | 304.95 | 299.62 | 277.21 | 10.42 |
| CAT (U/mg prot.) | 325.45 | 342.72 | 374.34 | 378.66 | 368.94 | 18.48 |
| GPx (mU/mg prot.) | 16.53 a | 13.10 b | 14.03 ab | 10.41 c | 10.26 c | 0.62 |
| GR (mU/mg prot.) | 7.73 | 6.30 | 6.82 | 6.29 | 6.65 | 0.56 |
| G6PDH (mU/mg prot.) | 57.74 | 50.21 | 47.67 | 44.28 | 44.08 | 7.31 |
| MDA (nmol/g tissue) | 65.06 b | 65.06 b | 89.58 ab | 99.54 a | 111.32 a | 7.12 |
| Plasma Metabolites | C | T | dT | TO1 | TO2 | SEM |
| Glucose (nmol/L) | 3.77 | 3.69 | 3.63 | 3.76 | 3.71 | 0.13 |
| Triglycerides (nmol/L) | 4.90 | 5.81 | 4.87 | 5.86 | 5.61 | 0.81 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melenchón, F.; Larrán, A.M.; Sanz, M.-Á.; Rico, D.; Fabrikov, D.; Barroso, F.G.; Galafat, A.; Alarcón, F.J.; Morales, A.E.; Hidalgo, M.C.; et al. Different Diets Based on Yellow Mealworm (Tenebrio molitor)—Part A: Facing the Decrease in Omega−3 Fatty Acids in Fillets of Rainbow Trout (Oncorhynchus mykiss). Fishes 2023, 8, 286. https://doi.org/10.3390/fishes8060286
Melenchón F, Larrán AM, Sanz M-Á, Rico D, Fabrikov D, Barroso FG, Galafat A, Alarcón FJ, Morales AE, Hidalgo MC, et al. Different Diets Based on Yellow Mealworm (Tenebrio molitor)—Part A: Facing the Decrease in Omega−3 Fatty Acids in Fillets of Rainbow Trout (Oncorhynchus mykiss). Fishes. 2023; 8(6):286. https://doi.org/10.3390/fishes8060286
Chicago/Turabian StyleMelenchón, Federico, Ana María Larrán, Miguel-Ángel Sanz, Daniel Rico, Dmitri Fabrikov, Fernando G. Barroso, Alba Galafat, Francisco Javier Alarcón, Amalia E. Morales, M. Carmen Hidalgo, and et al. 2023. "Different Diets Based on Yellow Mealworm (Tenebrio molitor)—Part A: Facing the Decrease in Omega−3 Fatty Acids in Fillets of Rainbow Trout (Oncorhynchus mykiss)" Fishes 8, no. 6: 286. https://doi.org/10.3390/fishes8060286
APA StyleMelenchón, F., Larrán, A. M., Sanz, M.-Á., Rico, D., Fabrikov, D., Barroso, F. G., Galafat, A., Alarcón, F. J., Morales, A. E., Hidalgo, M. C., Lourenço, H. M., Pessoa, M.-F., & Tomás-Almenar, C. (2023). Different Diets Based on Yellow Mealworm (Tenebrio molitor)—Part A: Facing the Decrease in Omega−3 Fatty Acids in Fillets of Rainbow Trout (Oncorhynchus mykiss). Fishes, 8(6), 286. https://doi.org/10.3390/fishes8060286