
Water Flow Requirements of Post-smolt Atlantic Salmon (Salmo salar L.) Reared in Intensive Seawater Flow-through Systems: A Physiological Perspective †


 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Stock and Rearing Conditions
2.2. Experimental Design
2.3. Blood Chemistry Sampling and Analysis
2.4. Gill Activity Sampling and Analysis
2.5. External Welfare Analysis
2.6. Water Quality and Specific Oxygen Consumption

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | 0.2 L kg−1 min−1 | 0.3 L kg−1 min−1 | 0.4 L kg−1 min−1 | 0.5 L kg−1 min−1 |
|---|---|---|---|---|
| Water flow (L min−1) | 7.5 | 11.25 | 15 | 18.75 |
| Tank exchange time (min) | 66.6 | 44.4 | 33.3 | 26.6 |
| Temperature (°C) | 9.3 ± 0.01 | 9.3 ± 0.01 | 9.3 ± 0.01 | 9.3 ± 0.01 |
| O2 (mg L−1) | 7.97 ± 0.01 a | 8.10 ± 0.01b | 9.82 ± 0.01 d | 8.63 ± 0.01 c |
| pH | 6.94 ± 0.05 a | 7.20 ± 0.05 b | 7.37 ± 0.04 c | 7.46 ± 0.05 c |
| CO2 (mg L−1) | 15.74 ± 1.83 a | 8.6 ± 0.88 b | 5.60 ± 0.48 bc | 4.79 ± 0.62 c |
| TAN (mg N L−1) NH3-N (µg L−1) | 0.76 ± 0.11 a 1.0 a | 0.48 ± 0.07 b 1.2 ab | 0.35 ± 0.05b 1.3 b | 0.36 ± 0.05 b 1.6 c |
2.7. Growth and Condition Factor
2.8. Statistics
3. Results
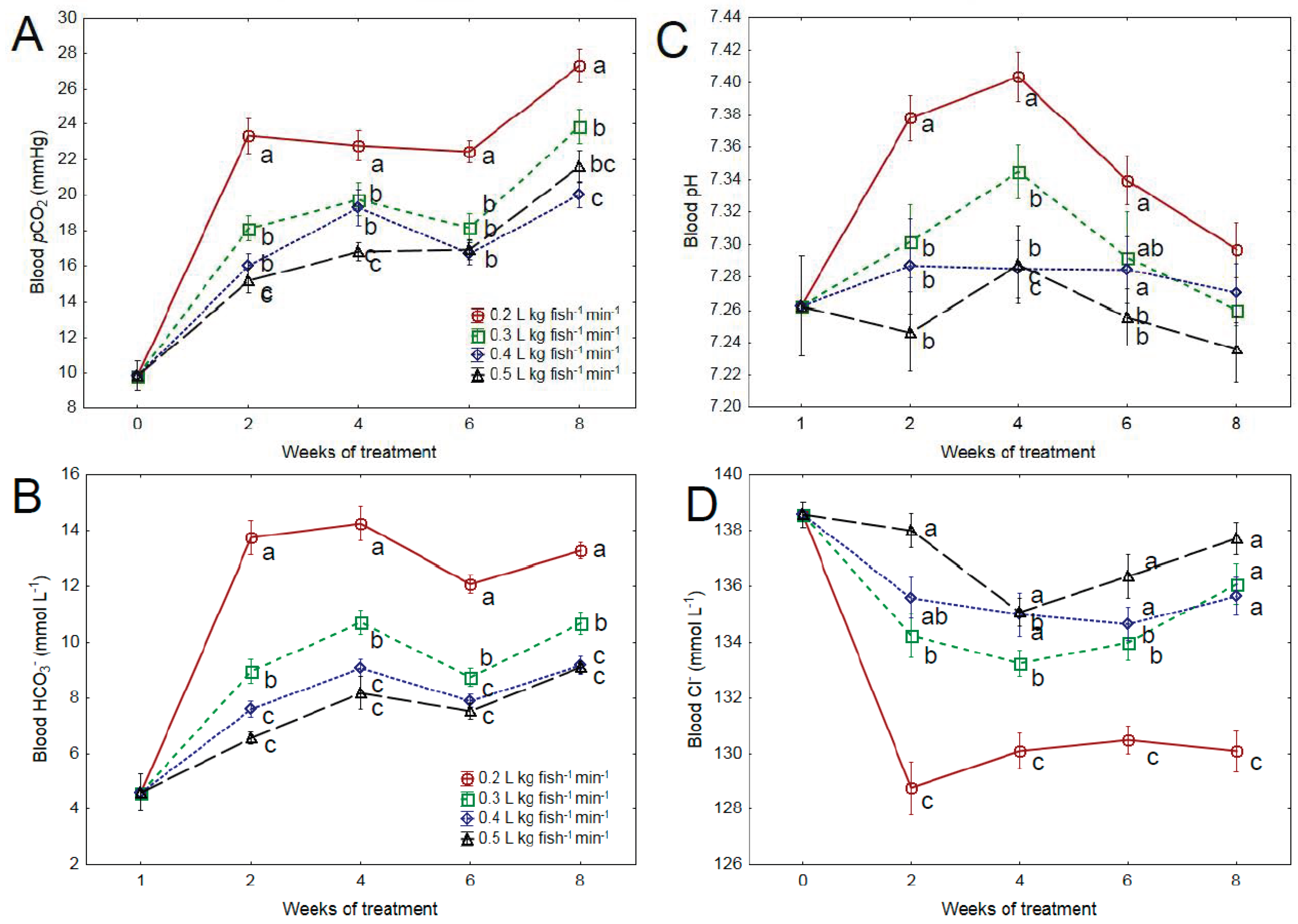
3.1. Blood Chemistry
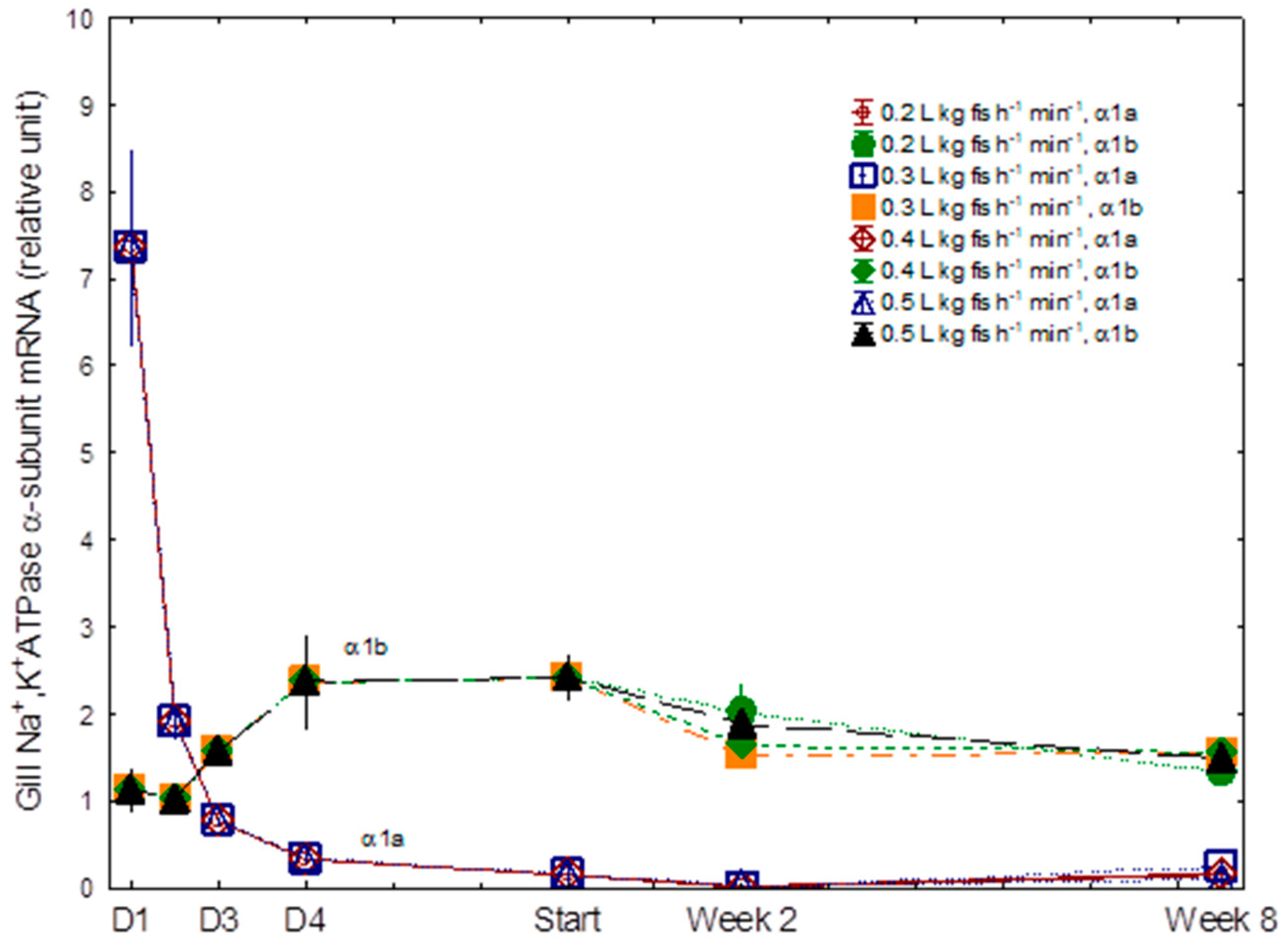
3.2. Gill NKA α1a and α1b mRNA Levels and Gill NKA
3.3. External Welfare Indicators
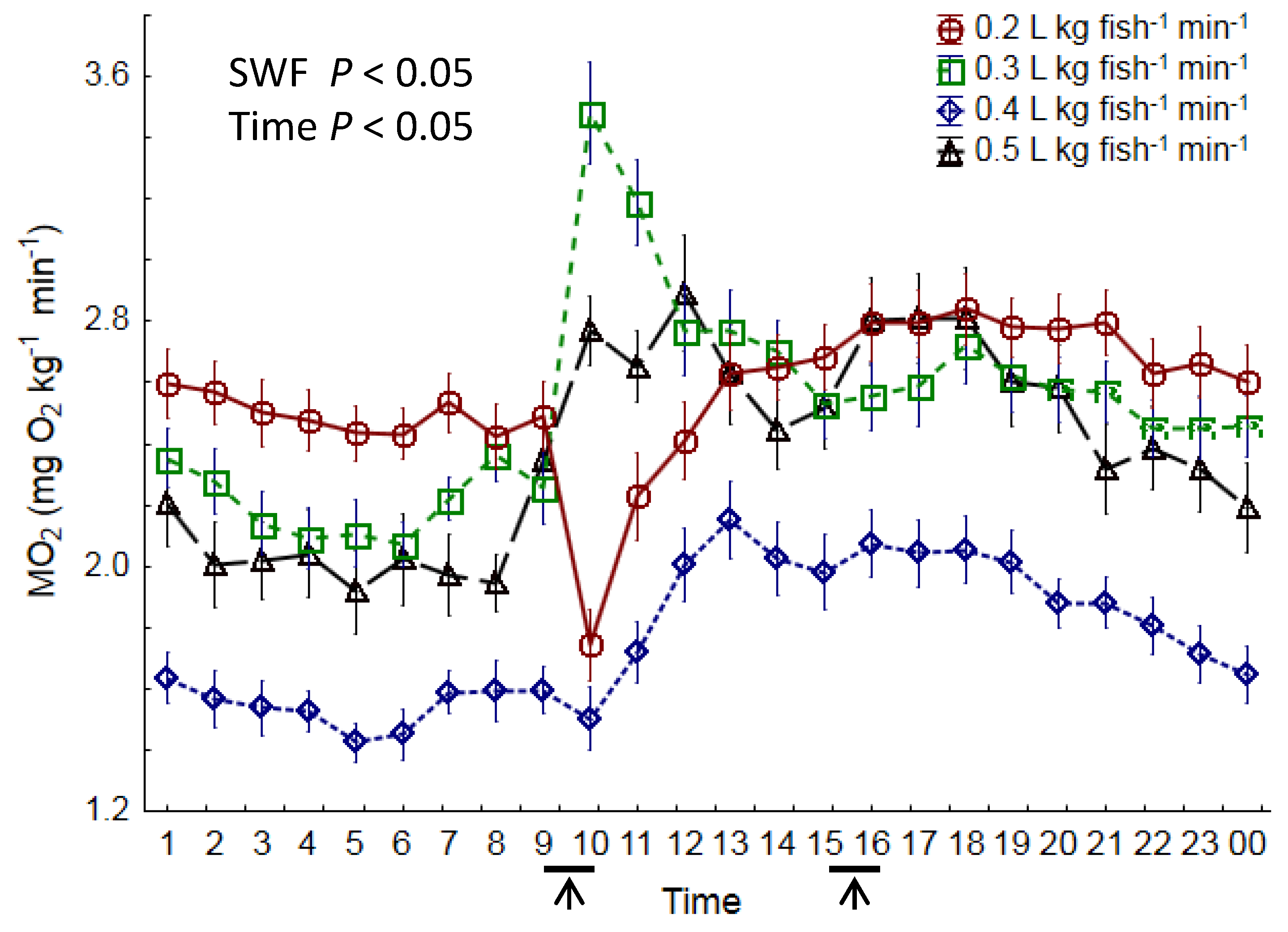
3.4. Oxygen Consumption
3.5. Growth and Condition Factors
4. Discussion
4.1. Water Quality and Blood Chemistry
4.2. Osmoregulation
4.3. External Welfare Indicators
4.4. Oxygen Consumption
4.5. Growth
4.6. Application at the Industrial Scale
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Pillay, T.V.R. Aquaculture and the Environment, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2008; p. 208. [Google Scholar]
- Gullestad, P.; Bjørgo, S.; Eithun, I.; Ervik, A.; Gudding, R.; Hansen, H.; Johansen, R.; Osland, A.; Rødseth, M.; Røsvik, I.; et al. Effektiv og Bærekraftig Arealbruk i Havbruksnæringen—Areal Til Begjær; Kystdepartementet: Oslo, Norway, 2011; p. 190. [Google Scholar]
- Thorarensen, H.; Farrell, A.P. The biological requirements for post-smolt Atlantic salmon in closed-containment systems. Aquaculture 2011, 312, 1–14. [Google Scholar] [CrossRef]
- Iversen, A.; Andreassen, O.; Hermansen, Ø.; Andre, T.; Terjesen, B.F. Aquaculture technology and competitiveness. Nofima. Rapp. 2013, 1–32. [Google Scholar]
- Fivelstad, S.; Binde, M. Effects of reduced waterflow (increased loading) in soft water on Atlantic salmon smolts (Salmo salar L.) while maintaining oxygen at constant level by oxygenation of the inlet water. Aquac. Eng. 1994, 13, 211–238. [Google Scholar] [CrossRef]
- Fivelstad, S.; Olsen, A.B.; Kloften, H.; Ski, H.; Stefansson, S. Effects of carbon dioxide on Atlantic salmon (Salmo salar L.) smolts at constant pH in bicarbonate rich freshwater. Aquaculture 1999, 78, 171–187. [Google Scholar] [CrossRef]
- Fivelstad, S.; Bergheim, A.; Holland, P.M.; Fjermedal, A.B. Water flow requirements in the intensive production of Atlantic salmon (Salmo salar L.) parr-smolt at two salinity levels. Aquaculture 2004, 231, 263–277. [Google Scholar] [CrossRef]
- Stefansson, S.O.; Nilsen, T.O.; Ebbesson, L.O.E.; Wargelius, A.; Madsen, S.S.; Bjornsson, B.T.; McCormick, S. Molecular mechanisms of continuous light inhibition of Atlantic salmon parr-smolt transformation. Aquaculture 2007, 273, 235–245. [Google Scholar] [CrossRef]
- Randall, D.J.; Tsui, T.K.N. Ammonia toxicity in fish. Mar. Poll. Bull. 2002, 45, 17–23. [Google Scholar] [CrossRef]
- Terjesen, B.F. Nitrogen excretion. In Fish Larval Physiology; Finn, R., Ed.; Science Publishers: New York, NY, USA, 2008; pp. 263–302. [Google Scholar]
- Randall, D.; Wright, P. The interaction between carbon dioxide and ammonia excretion and water pH in fish. Can. J. Zool. 1989, 67, 2936–2942. [Google Scholar] [CrossRef]
- Forsberg, O.I. The impact of varying feeding regimes on oxygen consumption and excretion of carbon dioxide and nitrogen in post-smolt Atlantic salmon Salmo salar L. Aquac. Res. 1997, 28, 29–41. [Google Scholar] [CrossRef]
- Bergheim, A.; Seymour, E.A.; Sanni, S.; Tyvold, T.; Fivelstad, S. Measurements of oxygen consumption and ammonia excretion of Atlantic salmon (Salmo salar L.) in commercial-scale, single-pass freshwater and seawater landbased culture systems. Aquac. Eng. 1991, 10, 251–267. [Google Scholar] [CrossRef]
- Sanni, S.; Forsberg, O.I. Modelling pH and carbon dioxide in single-pass sea-water aquaculture systems. Aquac. Eng. 1996, 15, 91–110. [Google Scholar] [CrossRef]
- Eddy, F.; Lomholt, J.; Weber, R.; Johansen, K. Blood respiratory properties of rainbow trout (Salmo gairdneri) kept in water of high CO2 tension. J. Exp. Biol. 1997, 67, 37–47. [Google Scholar] [CrossRef]
- Claiborne, J.B.; Edwards, S.L.; Morrison-Shetlar, A.I. Acid–base regulation in fishes: Cellular and molecular mechanisms. J. Exp. Zool. 2002, 293, 302–319. [Google Scholar] [CrossRef]
- Heisler, N. Role of ion transfer processes in acid-base regulation with temperature changes in fish. Am. J. Physiol. 1984, 246, R441–R451. [Google Scholar] [CrossRef]
- Cripps, S.J.; Bergheim, A. Solids management and removal for intensive land-based aquaculture production systems. Aquac. Eng. 2000, 22, 33–56. [Google Scholar] [CrossRef]
- Segner, H.; Sundh, H.; Buchmann, K.; Douxfils, J.; Sundell, S.K.; Mathieu, C.; Ruane, N.; Jutfelt, F.; Toften, H.; Vaughan, L. Health of farmed fish: Its relation to fish welfare and its utility as welfare indicator. Fish Physiol. Biochem. 2012, 38, 85–105. [Google Scholar] [CrossRef]
- Turnbull, J.; Kadri, S. Safeguarding the many guises of farmed fish welfare. Dis. Aquat. Org. 2007, 75, 173–182. [Google Scholar] [CrossRef]
- Kristensen, T.; Åtland, Å.; Rosten, T.; Urke, H.A.; Rosseland, B.O. Important influent-water quality parameters at freshwater production sites in two salmon producing countries. Aquac. Eng. 2009, 41, 53–59. [Google Scholar] [CrossRef]
- Wright, P.A.; Perry, S.F.; Moon, T.W. Regulation of hepatic gluconeogenesis and glycogenolysis by catecholamines in rainbow trout during environmental hypoxia. J. Exp. Biol. 1989, 147, 169–188. [Google Scholar] [CrossRef] [PubMed]
- Barton, B.A.; Iwama, G.K. Physiological changes in fish from stress in aquaculture with emphasis on the response and effects of corticosteroids. Ann. Rev. Fish Dis. 1991, 1, 3–26. [Google Scholar] [CrossRef]
- Wendelaar Bonga, S.E. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef]
- Korte, S.M.; Olivier, B.; Koolhaas, J.M. A new animal welfare concept based on allostasis. Physiol. Behav. 2007, 92, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Schreck, C.B. Stress and fish reproduction: The roles of allostasis and hormesis. Gen. Comp. Endocrinol. 2010, 165, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Sterling, P. Allostasis: A model of predictive regulation. Physiol. Behav. 2012, 106, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Broom, D.M. Animal welfare: Concepts and measurement. J. Anim. Sci. 1991, 69, 4167–4175. [Google Scholar] [CrossRef] [PubMed]
- Ellis, T.; North, B.; Scott, A.; Bromage, N.; Porter, M.; Gadd, D. The relationships between stocking density and welfare in farmed rainbow trout. J. Fish Biol. 2002, 61, 493–531. [Google Scholar] [CrossRef]
- Calabrese, S.; Nilsen, T.O.; Kolarevic, J.; Ebbesson, L.O.E.; Pedrosa, C.; Fivelstad, S.; Hosfeld, C.; Stefansson, S.; Terjesen, B.; Takle, H. Stocking density limits for post-smolt Atlantic salmon (Salmo salar L.) emphasis on production performance and welfare. Aquaculture 2007, 468, 363–370. [Google Scholar] [CrossRef]
- McCormick, S.D. Methods for Nonlethal Gill Biopsy and Measurement of Na+, K+-ATPase Activity. Can. J. Fish. Aquat. Sci. 1993, 50, 656–658. [Google Scholar] [CrossRef]
- Stefansson, S.O.; Björnsson, B.T.; Sundell, K.; Nyhammer, G.; McCormick, S.D. Physiological characteristics of wild Atlantic salmon post-smolts during estuarine and coastal migration. J. Fish Biol. 2003, 63, 942–955. [Google Scholar] [CrossRef]
- Boutilier, R.G.; Heming, T.A.; Iwama, G.K. Appendix: Physicochemical parameters for use in fish respiratory physiology. Fish Physiol. 1984, 10, 403–430. [Google Scholar]
- Boutilier, R.G.; West, T.G.; Webber, D.M.; Pogson, G.H.; Mesa, K.A.; Wells, J.; Wells, M.J. The protective effects of hypoxia-induced hypometabolism in the Nautilus. J. Comp. Physiol. B. 2000, 170, 261–268. [Google Scholar] [CrossRef]
- Harrenstien, L.A.; Tornquist, S.J.; Miller-Morgan, T.J.; Fodness, B.G.; Clifford, K.E. Evaluation of a point-of-care blood analyzer and determination of reference ranges for blood parameters in rockfish. J. Am. Vet. Med. Ass. 2005, 226, 255–265. [Google Scholar] [CrossRef]
- Cooke, S.J.; Suski, C.D.; Danylchuk, S.E.; Danylchuk, A.J.; Donaldson, M.F.; Pullen, C.; Bulte, G.; O’Toole, A.; Murchie, K.J.; Goldberg, T.J. Effects of different capture techniques on the physiological condition of bonefish Albula vulpes evaluated using field diagnostic tools. J. Fish Biol. 2008, 73, 1351–1375. [Google Scholar] [CrossRef]
- DiMaggio, M.A.; Ohs, C.L.; Petty, B.D. Evaluation of a Point-of-Care Blood Analyzer for Use in Determination of Select Hematological Indices in the Seminole Killifish. North Am. J. Aquac. 2010, 72, 261–268. [Google Scholar] [CrossRef]
- Harter, T.S.; Shartau, R.B.; Brauner, C.J.; Farrell, A.P. Validation of the i-STAT system for the analysis of blood parameters in fish. Cons. Physiol. 2014, 2, cou037. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Chomczynski, P. A reagent for the single-step simultaneous isolation of RNA, DNA and proteins from cell and tissue samples. Bio. Tech. 1993, 15, 532–535. [Google Scholar]
- Nilsen, T.O.; Ebbesson, L.O.; Madsen, S.S.; McCormick, S.D.; Anderson, E.; Bjornsson, B.T.; Prunet, P.; Stefansson, S.O. Differential expression of gill Na+, K+-ATPaseα-and β-subunits, Na+, K+, 2Cl-cotransporter and CFTR anion channel in juvenile anadromous and landlocked Atlantic salmon Salmo salar. J. Exp. Biol. 2007, 210, 2885–2896. [Google Scholar] [CrossRef]
- Nilsen, T.O.; Ebbesson, L.O.E.; Kverneland, O.G.; Kroglund, F.; Finstad, B.; Stefansson, S.O. Effects of acidic water and aluminum exposure on gill Na+, K+-ATPase α-subunit isoforms, enzyme activity, physiology and return rates in Atlantic salmon (Salmo salar L.). Aquat. Toxiol. 2010, 97, 250–259. [Google Scholar] [CrossRef]
- Pfaffl, M.W. Quantification strategies in real-time PCR. In AZ of Quantitative PCR; Bustin, S.A., Ed.; International University Line: La Jolla, CA, USA, 2004; pp. 89–113. [Google Scholar]
- Olsvik, P.A.; Lie, K.K.; Jordal, A.-E.O.; Nilsen, T.O.; Hordvik, I. Evaluation of potential reference genes in real-time RT-PCR studies of Atlantic salmon. BMC Molecul. Biol. 2005, 6, 21. [Google Scholar] [CrossRef]
- Kolarevic, J.; Baeverfjord, G.; Takle, H.; Ytteborg, E.; Reiten, B.K.M.; Nergård, S.; Terjesen, B.F. Performance and welfare of Atlantic salmon smolt reared in recirculating or flow through aquaculture systems. Aquaculture 2014, 432, 15–25. [Google Scholar] [CrossRef]
- Hoyle, I.; Oidtmann, B.; Ellis, T.; Turnbull, J.; North, B.; Nikolaidis, J.; Knowles, T.G. A validated macroscopic key to assess fin damage in farmed rainbow trout (Oncorhynchus mykiss). Aquaculture 2007, 270, 142–148. [Google Scholar] [CrossRef]
- Kolarevic, J.; Selset, R.; Felip, O.; Good, C.; Christopher, M.; Snekvik, K.; Takle, H.R.; Ytteborg, E.; Bæverfjord, G.; Åsgård, T.E.; et al. Influence of long term ammonia exposure on Atlantic salmon (Salmo salar L.) parr growth and welfare. Aquac. Res. 2013, 44, 1649–1664. [Google Scholar] [CrossRef]
- Fivelstad, S.; Olsen, A.B.; Åsgård, T.; Bæverfjord, G.; Rasmussen, T.; Vindheim, T.; Stefansson, S. Long-term sublethal effects of carbon dioxide on Atlantic salmon smolts (Salmo salar L.): Ion regulation, haematology, element composition, nephrocalcinosis and growth parameters. Aquaculture 2003, 215, 301–319. [Google Scholar] [CrossRef]
- Gebauer, R.; Eggen, G.; Hansen, E.; Eikebrokk, B. Oppdrettsteknologi; Tapir: Oslo, Norway, 1992; p. 576. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 2nd ed.; Prentice-Hall, Inc.: Englewood Cliffs, NJ, USA, 1984; p. 718. [Google Scholar]
- Lloyd, R.; White, W.R. Effect of High Concentration of Carbon Dioxide on the Ionic Composition of Rainbow Trout Blood. Nature 1967, 216, 1341–1342. [Google Scholar] [CrossRef]
- Fivelstad, S.; Haavik, H.; Lovik, G.; Olsen, A.B. Sublethal effects and safe levels of carbon dioxide in seawater for Atlantic salmon postsmolts (Salmo salar L.): Ion regulation and growth. Aquaculture 1998, 160, 305–316. [Google Scholar] [CrossRef]
- Hosfeld, C.D.; Engevik, A.; Mollan, T.; Lunde, T.M.; Waagbø, R.; Olsen, A.B.; Breck, O.; Stefansson, S.; Fivelstad, S. Long-term separate and combined effects of environmental hypercapnia and hyperoxia in Atlantic salmon (Salmo salar L.) smolts. Aquaculture 2008, 280, 146–153. [Google Scholar] [CrossRef]
- McCormick, S.D.; Regish, A.; Christensen, A. Distinct freshwater and seawater isoforms of Na+/K+-ATPase in gill chloride cells of Atlantic salmon. J. Exp. Biol. 2009, 212, 3994–4001. [Google Scholar] [CrossRef]
- Handeland, S.O.; Imsland, A.K.; Nilsen, T.O.; Ebbesson, L.O.E.; Hosfeld, C.D.; Pedrosa, C.; Toften, H.; Stefansson, S.O. Osmoregulation in Atlantic salmon Salmo salar smolts transferred to seawater at different temperatures. J. Fish Biol. 2014, 85, 1163–1176. [Google Scholar] [CrossRef]
- Sundh, H.; Olsen, R.E.; Fridell, F.; Gadan, K.; Evensen, Ø.; Glette, J.; Taranger, G.L.; Myklebust, R.; Sundell, K. The effect of hyperoxygenation and reduced flow in fresh water and subsequent infectious pancreatic necrosis virus challenge in sea water, on the intestinal barrier integrity in Atlantic salmon, Salmo salar L. J. Fish Dis. 2009, 32, 687–698. [Google Scholar] [CrossRef]
- Stien, L.H.; Bracke, M.B.M.; Folkedal, O.; Nilsson, J.; Oppedal, F.; Torgersen, T.; Kittilsen, S.; Midtlyng, P.J.; Vindas, M.A.; Øverli, Ø.; et al. Salmon Welfare Index Model (SWIM 1.0): A semantic model for overall welfare assessment of caged Atlantic salmon: Review of the selected welfare indicators and model presentation. Rev. Aquac. 2013, 5, 33–57. [Google Scholar] [CrossRef]
- Sveen, L.R.; Timmerhaus, G.; Torgersen, J.S.; Ytteborg, E.; Jørgensen, S.M.; Handeland, S.; Stefansson, S.O.; Nilsen, T.O.; Calabrese, S.; Ebbesson, L.; et al. Impact of fish density and specific water flow on skin properties in Atlantic salmon (Salmo salar L.) post-smolts. Aquaculture 2016, 464, 629–637. [Google Scholar] [CrossRef]
- Jobling, M. The influences of feeding on the metabolic rate of fishes: A short review. J. Fish Biol. 1981, 18, 385–400. [Google Scholar] [CrossRef]
- Pichavant, K.; Person-Le-Ruyet, J.; Bayon, N.L.; Severe, A.; Roux, A.L.; Boeuf, G. Comparative effects of long-term hypoxia on growth, feeding and oxygen consumption in juvenile turbot and European sea bass. J. Fish Biol. 2001, 59, 875–883. [Google Scholar] [CrossRef]
- Santos, G.A.; Schrama, J.W.; Capelle, J.; Rombout, J.H.W.M.; Verreth, J.A.J. Effects of dissolved carbon dioxide on energy metabolism and stress responses in European seabass (Dicentrarchus labrax). Aquac. Res. 2013, 44, 1370–1382. [Google Scholar] [CrossRef]
- Forsberg, O.I.; Bergheim, A. The impact of constant and fluctuating oxygen concentrations and two water consumption rates on post-smolt Atlantic salmon production parameters. Aquac. Eng. 1996, 15, 327–347. [Google Scholar] [CrossRef]
- Mathisen, F. Vannforbruksfaktor-ny Parameter for Vurdering av Intensitet og Vannbehov i Settefiskanlegg. Available online: https://www.kyst.no/vannforbruksfaktor-ny-parameter-for-vurdering-av-intensitet-og-vannbehov-i-settefiskanlegg/712880 (accessed on 15 March 2023). (In Norwegian).
- Storsul, T.; Arnfinn, A.; Dalum, A.S.; Romstad, S.; Urke, H.; Calabrese, S.; Larsson, T.; Marte, F. Aquatraz. Report to the Norwegian Directorate of Fisheries. Directorate of Fisheries: Bergen, Norway, 2021. [Google Scholar]
- Øvrebø, T.K.; Balseiro, P.; Imsland, A.K.D.; Stefansson, S.O.; Tveterås, R.; Sveier, H.; Handeland, S. Investigation of growth performance of post-smolt Atlantic salmon (Salmo salar L.) in semi closed containment system: A big-scale benchmark study. Aquac. Res. 2022, 53, 4178–4189. [Google Scholar] [CrossRef]
- Nilsen, A.; Nielsen, K.V.; Bergheim, A. A closer look at closed cages: Growth and mortality rates during production of post-smolt Atlantic salmon in marine closed confinement systems. Aquac. Eng. 2020, 91, 102124. [Google Scholar] [CrossRef]
- Føre, M.; Alver, M.; Alfredsen, J.A.; Marafioti, G.; Senneset, G.; Birkevold, J.; Willumsen, F.V.; Lange, G.; Espmark, Å.; Terjesen, B.F. Modelling growth performance and feeding behaviour of Atlantic salmon (Salmo salar L.) in commercial-size aquaculture net pens: Model details and validation through full-scale experiments. Aquaculture 2016, 464, 268–278. [Google Scholar] [CrossRef]





| Parameter | 0.2 L kg−1 min−1 | 0.3 L kg−1 min−1 | 0.4 L kg−1 min−1 | 0.5 L kg−1 min−1 |
|---|---|---|---|---|
| Cataract (0–2) | 0.1 ± 0.1 | 0.2 ± 0.1 | 0 ± 0 | 1 ± 0.1 |
| Skin (0−2) | 0.1 ± 0.1 | 0.4 ± 0.1 | 0.1 ± 0.1 | 0.4 ± 0.1 |
| Operculum (0–2) | 0 ± 0 | 0.1± 0.1 | 0.1 ± 0.1 | 0.2 ± 0.1 |
| Fin (0−5) | ||||
| Dorsal Caudal AnalPectoral Pelvic | 2.0 ± 0.4 | 2.35 ± 0.3 | 2.8 ± 0.3 | 2.3 ± 0.4 |
| 1.3 ± 0.1 | 1.5 ± 0.2 | 1.4 ± 0.2 | 1.4 ± 0.1 | |
| 0.3 ± 0.2 | 0.5 ± 0.1 | 1.2 ± 1.3 | 0.5 ± 0.2 | |
| 2.4 ± 0.1 | 2.3 ± 0.2 | 2.0 ± 0.1 | 2.2 ± 0.1 | |
| 1.4 ± 0.1 | 1.7 ± 0.1 | 1.8 ± 0.2 | 1.7 ± 0.1 |
| Parameter | 0.2 L kg−1 min−1 | 0.3 L kg−1 min−1 | 0.4 L kg−1 min−1 | 0.5 L kg−1 min−1 |
|---|---|---|---|---|
| Density range (kg m−3, n = 2) | 75.2–82.1 | 75.1–82.1 | 75.8–83.6 | 73.1–83.1 |
| Mortality (count, n = 2) | 1 | 1 | 4 | 9 |
| SGR (% day−1, n = 27–30) | 0.75 ± 0.03 | 0.78 ± 0.03 | 0.77 ± 0.03 | 0.77 ± 0.03 |
| W start (g, n = 30) | 121.1 ± 2.4 a | 113.8 ± 1.5 b | 110.7 ± 2.3 b | 108.7 ± 1.7 b |
| W end (g, n = 27–30) | 207.4 ± 7.1 | 195.0 ± 5.0 | 190.7 ± 5.6 | 187.2 ± 4.8 |
| L end (cm, n = 27–30) | 26.7 ± 0.3 | 26.4 ± 0.2 | 26.1± 0.3 | 25.9 ± 0.2 |
| K end (n = 27–30) | 1.07 ± 0.01 | 1.05 ± 0.01 | 1.06 ± 0.01 | 1.07 ± 0.01 |
| MO2 (mg kg−1 min−1, n = 28) | 2.56 ± 0.02 a | 2.55 ± 0.03 a | 1.75 ± 0.02 c | 2.38 ± 0.03 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calabrese, S.; Imsland, A.K.D.; Nilsen, T.O.; Kolarevic, J.; Ebbesson, L.O.E.; Hosfeld, C.D.; Fivelstad, S.; Pedrosa, C.; Terjesen, B.F.; Stefansson, S.O.; et al. Water Flow Requirements of Post-smolt Atlantic Salmon (Salmo salar L.) Reared in Intensive Seawater Flow-through Systems: A Physiological Perspective. Fishes 2023, 8, 285. https://doi.org/10.3390/fishes8060285
Calabrese S, Imsland AKD, Nilsen TO, Kolarevic J, Ebbesson LOE, Hosfeld CD, Fivelstad S, Pedrosa C, Terjesen BF, Stefansson SO, et al. Water Flow Requirements of Post-smolt Atlantic Salmon (Salmo salar L.) Reared in Intensive Seawater Flow-through Systems: A Physiological Perspective. Fishes. 2023; 8(6):285. https://doi.org/10.3390/fishes8060285
Chicago/Turabian StyleCalabrese, Sara, Albert K. D. Imsland, Tom Ole Nilsen, Jelena Kolarevic, Lars O. E. Ebbesson, Camilla Diesen Hosfeld, Sveinung Fivelstad, Cindy Pedrosa, Bendik Fyhn Terjesen, Sigurd O. Stefansson, and et al. 2023. "Water Flow Requirements of Post-smolt Atlantic Salmon (Salmo salar L.) Reared in Intensive Seawater Flow-through Systems: A Physiological Perspective" Fishes 8, no. 6: 285. https://doi.org/10.3390/fishes8060285
APA StyleCalabrese, S., Imsland, A. K. D., Nilsen, T. O., Kolarevic, J., Ebbesson, L. O. E., Hosfeld, C. D., Fivelstad, S., Pedrosa, C., Terjesen, B. F., Stefansson, S. O., Takle, H., Sveier, H., Mathisen, F., & Handeland, S. O. (2023). Water Flow Requirements of Post-smolt Atlantic Salmon (Salmo salar L.) Reared in Intensive Seawater Flow-through Systems: A Physiological Perspective. Fishes, 8(6), 285. https://doi.org/10.3390/fishes8060285