
Effects of γ-Irradiated Date (Phoenix dactylifera) Fruit on Growth, Immunological and Antioxidant Parameters of Goldfish (Carassius auratus)



Abstract
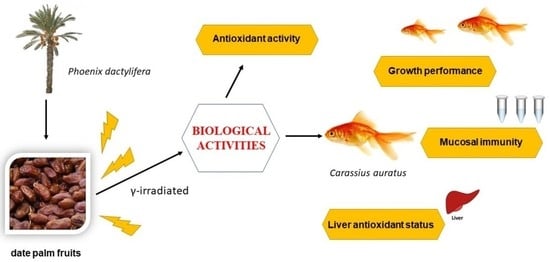
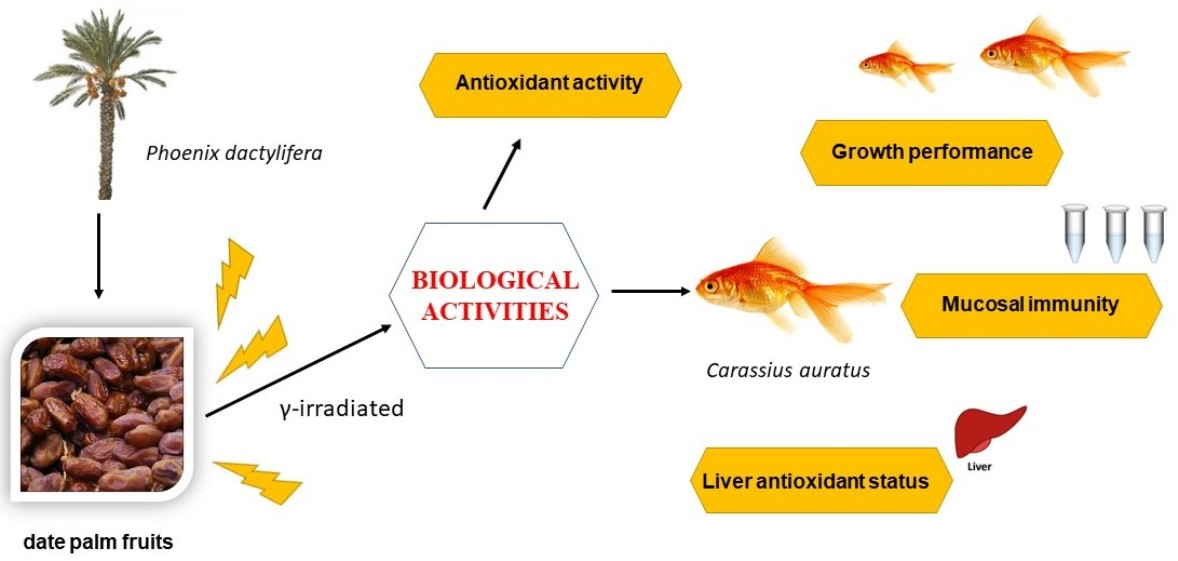
1. Introduction
2. Materials and Methods
2.1. P. dactylifera Extract and Diet Preparation
2.2. Characterization of IPFE
2.3. Fish Rearing and Experimental Design
2.4. Fish Growth and Sampling
2.5. Skin Mucus Immunity
2.6. Liver Antioxidant Parameters
2.7. Statistical Analysis
3. Results
3.1. Analysis of IPFE
3.2. Fish Growth
3.3. Mucus Immune Parameters
3.4. Liver Antioxidant Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Noga, E.J. Fish Disease: Diagnosis and Treatment, 2nd ed.; Willey-Blackwell: Hoboken, NJ, USA, 2010; 536p. [Google Scholar]
- Cardoso, P.H.M.; Moreno, A.M.; Moreno, L.Z.; Oliveira, C.H.; Baroni, F.A.; Maganha, S.R.L. Infectious diseases in aquarium ornamental pet fish: Prevention and control measures. Braz. J. Vet. Res. Anim. Sci. 2019, 56, e151697. [Google Scholar] [CrossRef]
- Roberts, H.E.; Palmeiro, B.; Weber, E.S. Bacterial and parasitic diseases of pet fish. Vet. Clin. N. Am. Exot. Anim. Pract. 2009, 12, 609–638. [Google Scholar] [CrossRef] [PubMed]
- Soni, M.; Oureshi, Q.A.; Mishra, M.; Nishad, C.S.; Chhaba, B.; Das, S.A. Common Aeromonas Infections in Ornamental Fishes: A review. Biol. Forum. 2021, 13, 433–439. [Google Scholar]
- Herrera, M.; Mancera, J.M.; Costas, B. The Use of Dietary Additives in Fish Stress Mitigation: Comparative Endocrine and Physiological Responses. Front. Endocrinol. 2019, 10, 447. [Google Scholar] [CrossRef]
- Hasan, M.T.; Jang, W.J.; Lee, J.M.; Lee, B.J.; Hur, S.W.; Lim, S.G. Effects of Immunostimulants, Prebiotics, Probiotics, Synbiotics, and Potentially Immunoreactive Feed Additives on Olive Flounder (Paralichthys olivaceus): A Review. Rev. Fish. Sci. Aquac. 2019, 27, 417–437. [Google Scholar] [CrossRef]
- Mohan, K.; Ravichandran, S.; Muralisankar, T.; Uthayakumar, V.; Chandirasekar, R.; Seedevi, P.; Rajan, D.K. Potential uses of fungal polysaccharides as immunostimulants in fish and shrimp aquaculture: A review. Aquaculture 2019, 500, 250–263. [Google Scholar] [CrossRef]
- Vijayaram, S.; Sun, Y.Z.; Zuorro, A.; Ghafarifarsani, H.; Doan, H.V.; Hoseinifar, S.H. Bioactive immunostimulants as health-promoting feed additives in aquaculture: A review. Fish Shellfish. Immunol. 2022, 130, 294–308. [Google Scholar] [CrossRef]
- Hai, N.V. The use of medicinal plants as immunostimulants in aquaculture: A review. Aquaculture 2015, 446, 88–96. [Google Scholar]
- Ashraf, Z.; Hamidi-Esfahani, Z. Date and Date Processing: A Review. Food Rev. Int. 2011, 27, 101–133. [Google Scholar] [CrossRef]
- Hussain, M.I.; Farooq, M.; Syed, Q.A. Nutritional and biological characteristics of the date palm fruit (Phoenix dactylifera L.)—A review. Food Biosci. 2020, 34, 100509. [Google Scholar] [CrossRef]
- Younas, A.; Naqvi, S.A.; Khan, M.R.; Shabbir, M.A.; Jatoi, M.A.; Anwar, F. Functional food and nutra-pharmaceutical perspectives of date (Phoenix dactylifera L.) fruit. J. Food Biochem. 2020, 44, e13332. [Google Scholar] [CrossRef] [PubMed]
- Esteban, M.A.; Cordero, H.; Martínez-Tomé, M.; Jiménez-Monreal, A.M.; Bakhrouf, A.; Mahdhi, A. Effect of dietary supplementation of probiotics and palm fruits extracts on the antioxidant enzyme gene expression in the mucosae of gilthead seabream (Sparus aurata L.). Fish Shellfish. Immunol. 2014, 39, 532–540. [Google Scholar] [CrossRef] [PubMed]
- Guardiola, F.A.; Porcino, C.; Cerezuela, R.; Cuesta, A.; Faggio, C.; Esteban, M.A. Impact of date palm fruits extracts and probiotic enriched diet on antioxidant status, innate immune response and immune-related gene expression of European seabass (Dicentrarchus labrax). Fish Shellfish. Immunol. 2016, 52, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Hoseinifar, S.H.; Dadar, M.; Khalili, M.; Cerezuela, R.; Esteban, M.Á. Effect of dietary supplementation of palm fruit extracts on the transcriptomes of growth, antioxidant enzyme and immune-related genes in common carp (Cyprinus carpio) fingerlings. Aquac. Res. 2017, 48, 3684–3692. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Khalili, M.; Rufchaei, R.; Raeisi, M.; Attar, M.; Cordero, H. Effects of date palm fruit extracts on skin mucosal immunity, immune related genes expression and growth performance of common carp (Cyprinus carpio) fry. Fish Shellfish Immunol. 2015, 47, 706–711. [Google Scholar] [CrossRef]
- Ahuja, S.; Kumar, M.; Kumar, P.; Gupta, V.K.; Singhal, R.K.; Yadavet, A. Metabolic and biochemical changes caused by gamma irradiation in plants. J. Radioanal. Nucl. Chem. 2014, 300, 199–212. [Google Scholar] [CrossRef]
- Rezanejad, R.; Ojagh, S.M.; Heidarieh, M.; Raeisi, M.; Rafiee, G.; Alishahi, A.R. Characterization of Gamma-Irradiated Rosmarinus officinalis L. (Rosemary). Turk. J. Pharm. Sci. 2019, 16, 43–47. [Google Scholar] [CrossRef]
- Zarbakhsh, S.; Rastegar, S. Influence of postharvest gamma irradiation on the antioxidant system, microbial and shelf life quality of three cultivars of date fruits (Phoenix dactylifera L.). Sci. Hortic. 2019, 247, 275–286. [Google Scholar] [CrossRef]
- Sheikhzadeh, N.; Ahmadifar, E.; Dawood, M.A.O.; Soltani, M. Dietary sodium propionate enhanced the growth performance, immune-related genes expression, and resistance against Ichthyophthirius multifiliis in goldfish (Carassius auratus). Aquaculture 2021, 540, 736720. [Google Scholar] [CrossRef]
- Mousavi, S.; Sheikhzadeh, N.; Tayefi-Nasrabadi, H.; Alizadeh-Salteh, S.; Khani Oushani, A.; Firouzamandi, M.; Mardani, K. Administration of grape (Vitis vinifera) seed extract to rainbow trout (Oncorhynchus mykiss) modulates growth performance, some biochemical parameters, and antioxidant-relevant gene expression. Fish Physiol. Biochem. 2020, 46, 777–786. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Yen, G.C.; Chen, H.Y. Antioxidant Activity of Various Tea Extracts in Relation to Their Antimutagenicity. J. Agric. Food Chem. 1995, 43, 27–32. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Meth Enzymol. 1999, 299, 152–178. [Google Scholar]
- Chang, C.; Yang, M.; Wen, H.; Chern, J. Estimation of total flavonoid content in propolis by two complementary Colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar]
- Sheikhzadeh, N.; Karimi Pashaki, A.; Nofouzi, K.; Heidarieh, M.; Tayefi-Nasrabadi, H. Effects of dietary Ergosan on cutaneous mucosal immune response in rainbow trout (Oncorhynchus mykiss). Fish Shellfish. Immunol. 2012, 32, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Siwicki, A.K.; Anderson, D.P.; Rumsey, G.L. Dietary intake of immunostimulants by rainbow trout affects non-specific immunity and protection against furunculosis. Vet. Immunol. Immunop. 1994, 41, 125–139. [Google Scholar] [CrossRef]
- Ahn, D.U.; Olson, D.G.; Jo, C.; Chen, X.; Wu, C.; Lee, J.I. Effect of muscle type, packaging, and irradiation on lipid oxidation, volatile production and color in raw pork patties. Meat Sci. 1998, 49, 27–39. [Google Scholar] [CrossRef]
- Góth, L.A. Simple method for determination of serum catalase and revision of reference range. Clin. Chim. Acta 1991, 196, 143–152. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jacoby, W.B. Glutathione S-transferases, the first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 321–336. [Google Scholar]
- Maqsood, S.; Adiamo, O.; Ahmad, M.; Mudgil, P. Bioactive compounds from date fruit and seed as potential nutraceutical and functional food ingredients. Food Chem. 2020, 308, 125522. [Google Scholar] [CrossRef]
- Heidarieh, M.; Nabipour, A.; Shahbazi, S.; Shawrang, P.; Zhang, B. Effect of gamma irradiation processing on total phenol and antioxidant capacities of the Iranian extract of propolis. Radiochim. Acta 2021, 109, 635–641. [Google Scholar] [CrossRef]
- Saafi, E.B.; Arem, A.E.; Issaoui, M.; Hammami, M.; Achour, L. Phenolic content and antioxidant activity of four date palm (Phoenix dactylifera L.) fruit varieties grown in Tunisia. Int. J. Food Sci. Technol. 2009, 44, 2314–2319. [Google Scholar] [CrossRef]
- Hasan, N.; Amon, Z.; Nor, A.I.; Monkhtarrudin, N.; Esa, N.; Azlan, A. Nutritional composition and in vitro evaluation of the antioxidant properties of various dates extracts (Phoenix dactylifera L.) from Libya. Asian J. Clin. Nutr. 2010, 2, 208–214. [Google Scholar] [CrossRef]
- Cabo Verde, S.; Trigo, M.J.; Sousa, M.B.; Ferreira, A.; Ramos, C.; Nunes, I. Effects of gamma radiation on raspberries: Safety and quality issues. J. Toxicol. Environ. Health 2013, 76, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Harrison, K.; Were, L.M. Effect of gamma irradiation on total phenolic content yield and antioxidant capacity of Almond skin extracts. Food Chem. 2007, 102, 932–937. [Google Scholar] [CrossRef]
- Guo, C.; Yang, J.; Wei, J.; Li, Y.; Xu, J.; Jiang, Y. Antioxidant activities of peel, pulp and seed fractions of common fruits as determined by FRAP assay. Nutr. Res. 2003, 23, 1719–1726. [Google Scholar] [CrossRef]
- Thongprajukaew, K.; Kovitvadhi, U.; Kovitvadhi, S.; Somsueb, P.; Rungruangsak-Torrissen, K. Effects of different modified diets on growth, digestive enzyme activities and muscle compositions in juvenile Siamese fighting fish (Betta splendens Regan, 1910). Aquaculture 2011, 322, 1–9. [Google Scholar] [CrossRef]
- Sheikhzadeh, N.; Chehrara, F.; Heidarieh, M.; Katayoon, N.; Baradaran, B. Effects of irradiated Ergosan on the growth performance and mucus biological components of rainbow trout Oncorhynchus mykiss. Chin. J. Ocean. Limnol. 2016, 34, 13–18. [Google Scholar] [CrossRef]
- Heidarieh, M.; Banaee, M.; Heidarieh, H.; Gholamhosseini, A. Effects of gamma-irradiated ethanolic extract of Iranian propolis on growth performance, immunological and haematological parameters in juvenile common carp (Cyprinus carpio L.). Int. J. Aquat. Biol. 2022, 9, 393–402. [Google Scholar]
- Khosravi, M.; Dastar, B.; Aalami, M.; Shawrang, P.; Ashayerizade, O. Comparison of gamma–Irradiation and enzyme supplementation to eliminate antinutritional factors in rice bran in broiler chicken diets. Livest. Sci. 2016, 191, 51–56. [Google Scholar] [CrossRef]
- Eid, N.; Enani, S.; Walton, G.; Corona, G.; Costabile, A.; Gibson, G. The impact of date palm fruits and their component polyphenols, on gut microbial ecology, bacterial metabolites and colon cancer cell Proliferation. J. Nutr. Sci. 2014, 3, e46. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Yi, B.; Zhong, R.; Wang, M.; Zhang, S.; Ma, J. From gut microbiota to host appetite: Gut microbiota-derived metabolites as key regulators. Microbiome 2021, 9, 162. [Google Scholar] [CrossRef] [PubMed]
- Gomez, G.; Pei, J.; Mwangi, W.; Adams, L.G.; Rice-Ficht, A.; Ficht, T.A. Immunogenic and Invasive Properties of Brucella melitensis 16M Outer Membrane Protein Vaccine Candidates Identified via a Reverse Vaccinology Approach. PLoS ONE 2013, 8, e59751. [Google Scholar] [CrossRef] [PubMed]
- Nematallahi, M.A.; Izadi, M.; Ziaie, F.; Mirvaghefi, A.; Heidarieh, M.; Mahmoodi, S. Comparison between of effect of Ergosan particles and nano-particles as produced by using Gamma rays and ultrasonic waves on the growth and stress parameters indices of rainbow trout (Oncorhynchus mykiss). Aquac. Res. 2020, 7, 123–131. [Google Scholar]
- Ahmadifar, E.; Yousefi, M.; Karimi, M.; Fadaei Raieni, R.; Dadar, M.; Sevdan Yilmaz, S. Benefits of Dietary Polyphenols and Polyphenol-Rich Additives to Aquatic Animal Health: An Overview. Rev. Fish Sci. Aquac. 2020, 29, 478–511. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef]
- Lin, S.; Wang, Z.; Lam, K.L.; Zeng, S.; Tan, B.K.; Hu, J. Role of intestinal microecology in the regulation of energy metabolism by dietary polyphenols and their metabolites. Food Nutr. Res. 2019, 63, 1518. [Google Scholar] [CrossRef]
- Neyestani, T.R. Polyphenols and Immunity. In Wild-Type Food in Health Promotion and Disease Prevention; Watson, R.R., DeMeester, F., Eds.; Humana Press Inc.: Totowa, NJ, USA, 2008; pp. 413–434. [Google Scholar]
- Mohammadi, M.; Soltani, M.; Siahpoosh, A.; Shamsaei Mehrjan, M. Effects of date palm (Phoenix dactylifera) seed extract on heavy metals concentrations in carp (Cyprinus carpio). Pol. J. Environ. Stud. 2016, 25, 1117–1123. [Google Scholar] [CrossRef]
- Huang, S.J.; Mau, J.L. Antioxidant properties of methanolic extracts from Antrodia camphorata with various doses of γ-irradiation. Food Chem. 2007, 105, 1702–1710. [Google Scholar] [CrossRef]
- Adamo, M.; Capitani, D.; Mannina, L.; Cristinzio, M.; Ragni, P.; Tata, A. Decontamination treatment by ionizing radiation. Radiat. Phys. Chem. 2004, 71, 167–170. [Google Scholar] [CrossRef]
- Olszowy, M. What is responsible for antioxidant properties of polyphenolic compounds from plants? Plant Physiol. Biochem. 2019, 144, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Bhuyan, U.; Handique, J.G. Chapter 6-Plant polyphenols as potent antioxidants: Highlighting the mechanism of antioxidant activity and synthesis/development of some polyphenol conjugates. In Studies in Natural Products Chemistry; Atta-Ur Rahman, Ed.; Elsevier: Amsterdam, The Netherlands, 2022; Volume 75, pp. 243–266. [Google Scholar]
- Harrell, C.R.; Djonov, V.; Fellabaum, C.; Volarevic, V. Risks of Using Sterilization by Gamma Radiation: The Other Side of the Coin. Int. J. Med. Sci. 2018, 15, 274–279. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Components | (%) |
|---|---|
| Kilkafish meal a | 18 |
| Soybean meal b | 35 |
| Wheat flour | 26 |
| Cottonseed meal | 15 |
| Cellulose | 1 |
| Vitamin mixture c | 2.5 |
| Mineral mixture d | 2.5 |
| Chemical composition (% dry matter) | |
| Dry matter | 87.80 |
| Crude protein | 33.14 |
| Crude lipid | 6.18 |
| Ash | 5.57 |
| Gross energy (kcal kg−1) | 3948.91 |
| Extract | DPPH (µ/mL) | ABTS (µ/mL) | Total Phenol (mg GAE/g Extract) | Total Flavonoid (mg Quercetin/g Extract) |
|---|---|---|---|---|
| IPFE0 | 0.53 ± 0.01 c | 3.02 ± 0.15 c | 6.13 ± 0.32 c | 3.61 ± 0.22 b |
| IPFE5 | 0.71 ± 0.01 a | 4.76 ± 0.29 a | 7.62 ± 0.19 a | 4.80 ± 0.34 a |
| IPFE10 | 0.59 ± 0.01 b | 3.96 ± 0.34 b | 6.81 ± 0.08 b | 3.90 ± 0.42 a |
| Treatment Groups | Growth Indices | ||||||
|---|---|---|---|---|---|---|---|
| Initial Weight (g) | Initial Length (cm) | Final Weight (g) | Final Length (cm) | Dry Feed Intake (g) | SGR | FCR | |
| CTR | 5.53 ± 0.12 | 4.83 ± 0.10 | 9.21 ± 0.38 c | 6.69 ± 0.36 c | 10.12 ± 0.05 c | 1.25 ± 0.02 c | 2.77 ± 0.09 c |
| IPFE0 | 5.50 ± 0.11 | 4.77 ± 0.12 | 10.18 ± 0.25 b | 7.06 ± 0.11 c | 12.08 ± 0.08 b | 1.53 ± 0.07 b | 2.58 ± 0.15 bc |
| IPFE5 | 5.58 ± 0.11 | 4.85 ± 0.09 | 11.45 ± 0.51 a | 7.93 ± 0.20 a | 13.35 ± 0.08 a | 1.79 ± 0.22 a | 2.29 ± 0.06 a |
| IPFE10 | 5.52 ± 0.10 | 4.80 ± 0.13 | 11.08 ± 0.09 a | 7.51 ± 0.09 b | 13.67 ± 0.09 a | 1.71 ± 0.06 b | 2.46 ± 0.09 b |
| Treatment Groups | Mucus Immunological Parameters | ||||
|---|---|---|---|---|---|
| Lysozyme Activity (U mg−1) | Alkaline Phosphatase (U mg−1) | Protease (U mg−1) | Ig Level (mg ml−1) | MIC | |
| CTR | 36.13 ± 0.46 d | 23.19 ± 0.16 c | 35.08 ± 2.11 d | 8.01 ± 1.62 b | 0.67 ± 0.12 b |
| IPFE0 | 60.27 ± 1.01 c | 88.27 ± 1.21 b | 58.11 ± 3.07 c | 12.86 ± 2.98 a | 2.33 ± 0.48 ab |
| IPFE5 | 78.97 ± 1.89 a | 96.27 ± 1.55 a | 69.42 ± 1.88 a | 13.98 ± 1.31 a | 6.67 ± 0.33 a |
| IPFE10 | 70.15 ± 1.58 b | 87.29 ± 0.98 b | 63.61 ± 2.09 b | 12.65 ± 1.02 a | 5.33 ± 0.33 ab |
| Treatment Groups | Liver Antioxidant Parameters | ||||
|---|---|---|---|---|---|
| Lipid Peroxidation Product (µmol mg−1) | Superoxide Dismutase (U mg−1) | Catalase (U mg−1) | Glutathione S-Transferase (U mg−1) | Glutathione Peroxidase | |
| (U mg−1) | |||||
| CTR | 30.36 ± 1.28 b | 6.25 ± 0.15 c | 9.25 ± 0.93 c | 9.02 ± 0.21 c | 10.09 ± 0.26 d |
| IPFE0 | 15.75 ± 0.33 a | 7.18 ± 0.26 b | 10.55 ± 1.02 b | 9.83 ± 0.15 b | 14.88 ± 0.48 c |
| IPFE5 | 16.11 ± 1.07 a | 9.04 ± 0.82 a | 12.81 ± 1.03 a | 11.63 ± 0.19 a | 20.06 ± 0.31 a |
| IPFE10 | 15.87 ± 1.08 a | 8.93 ± 1.03 a | 12.51 ± 1.19 a | 11.41 ± 0.32 a | 17.43 ± 0.38 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heidarieh, M.; Gholamhosseini, A.; Sheikhzadeh, N.; Esteban, M.A. Effects of γ-Irradiated Date (Phoenix dactylifera) Fruit on Growth, Immunological and Antioxidant Parameters of Goldfish (Carassius auratus). Fishes 2023, 8, 251. https://doi.org/10.3390/fishes8050251
Heidarieh M, Gholamhosseini A, Sheikhzadeh N, Esteban MA. Effects of γ-Irradiated Date (Phoenix dactylifera) Fruit on Growth, Immunological and Antioxidant Parameters of Goldfish (Carassius auratus). Fishes. 2023; 8(5):251. https://doi.org/10.3390/fishes8050251
Chicago/Turabian StyleHeidarieh, Marzieh, Amin Gholamhosseini, Najmeh Sheikhzadeh, and Maria Angeles Esteban. 2023. "Effects of γ-Irradiated Date (Phoenix dactylifera) Fruit on Growth, Immunological and Antioxidant Parameters of Goldfish (Carassius auratus)" Fishes 8, no. 5: 251. https://doi.org/10.3390/fishes8050251
APA StyleHeidarieh, M., Gholamhosseini, A., Sheikhzadeh, N., & Esteban, M. A. (2023). Effects of γ-Irradiated Date (Phoenix dactylifera) Fruit on Growth, Immunological and Antioxidant Parameters of Goldfish (Carassius auratus). Fishes, 8(5), 251. https://doi.org/10.3390/fishes8050251