
Piscine orthoreovirus Genotype-1 (PRV-1) in Wild Pacific Salmon of British Columbia, Canada: 2011–2020


Abstract
1. Introduction
2. Material and Methods
2.1. Sample Collection
2.1.1. Freshwater: Hatchery and Lower Fraser River
2.1.2. Marine Waters: Strait of Georgia/Lower Johnstone Strait
2.1.3. Marine Waters: West Coast Vancouver Island
2.1.4. Laboratory Dissections
2.2. RNA Extraction and Reverse Transcription
2.3. PRV-1 qPCR Screening
2.4. Genetic Stock Identification
3. Results
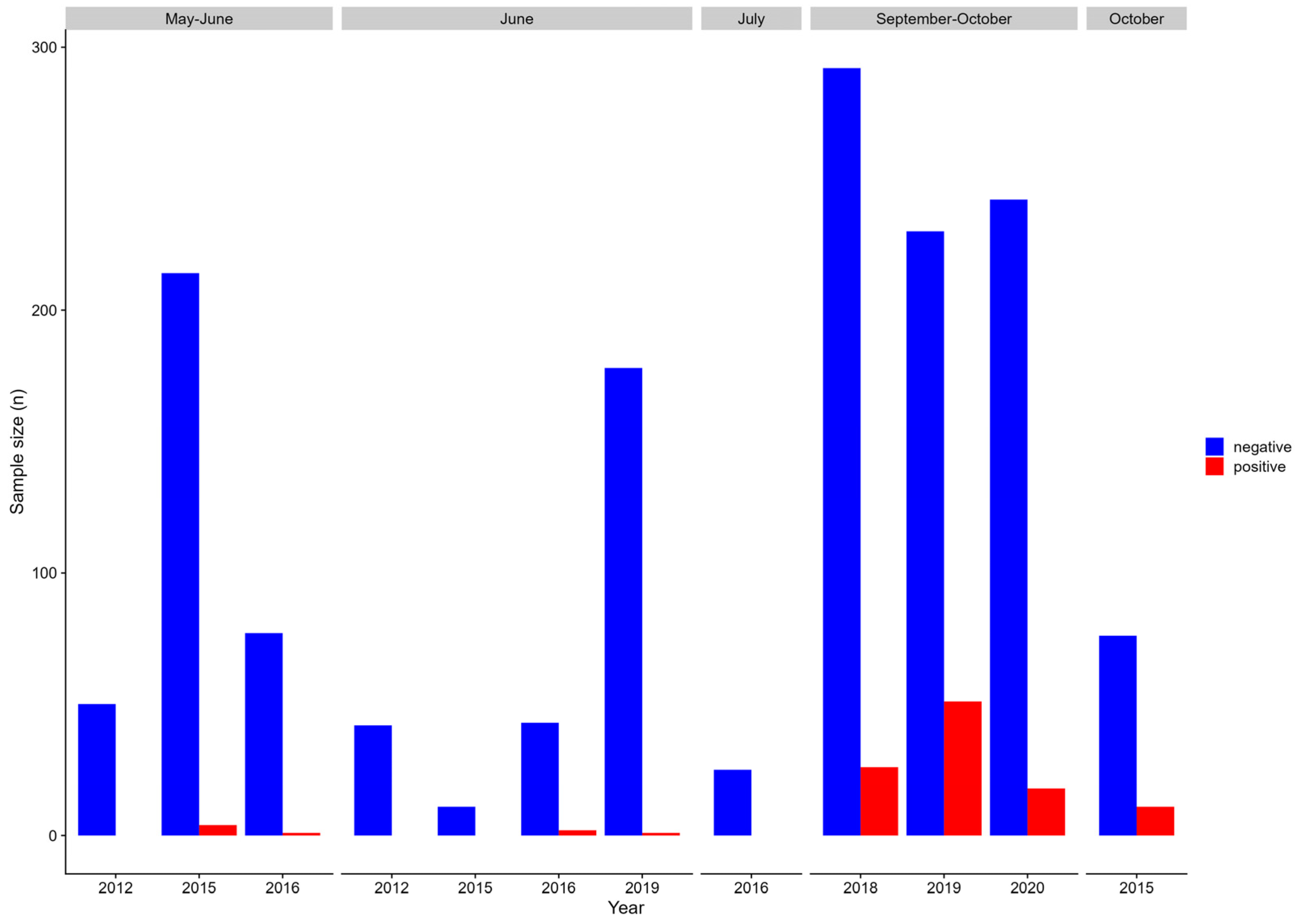
3.1. Prevalence of PRV-1 in Juvenile Chum, Pink and Sockeye Salmon
3.2. Prevalence of PRV-1 in Coho Salmon
3.3. Prevalence of PRV-1 in Chinook Salmon
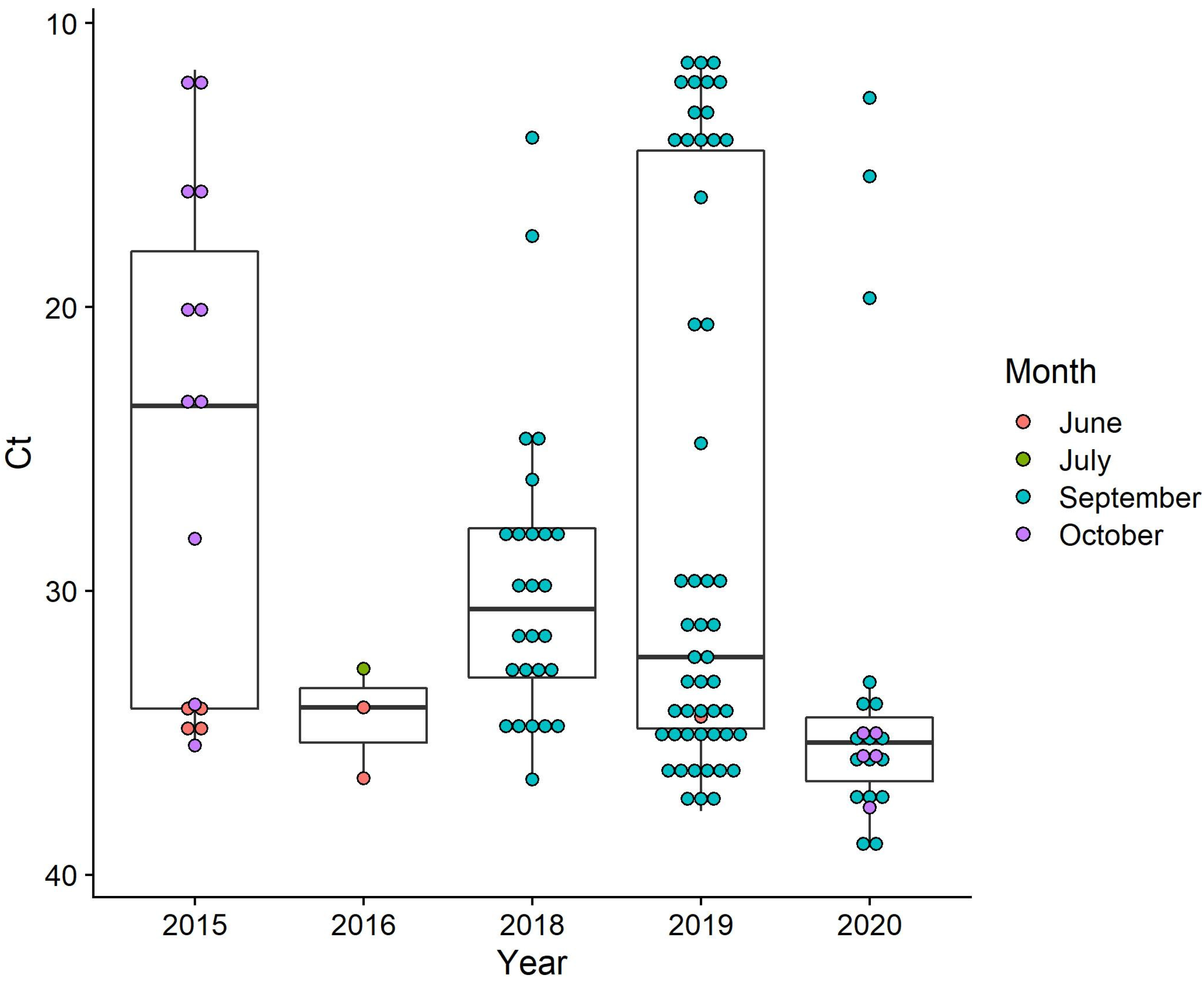
3.3.1. Relative PRV-1 Loads in Juvenile Chinook Salmon
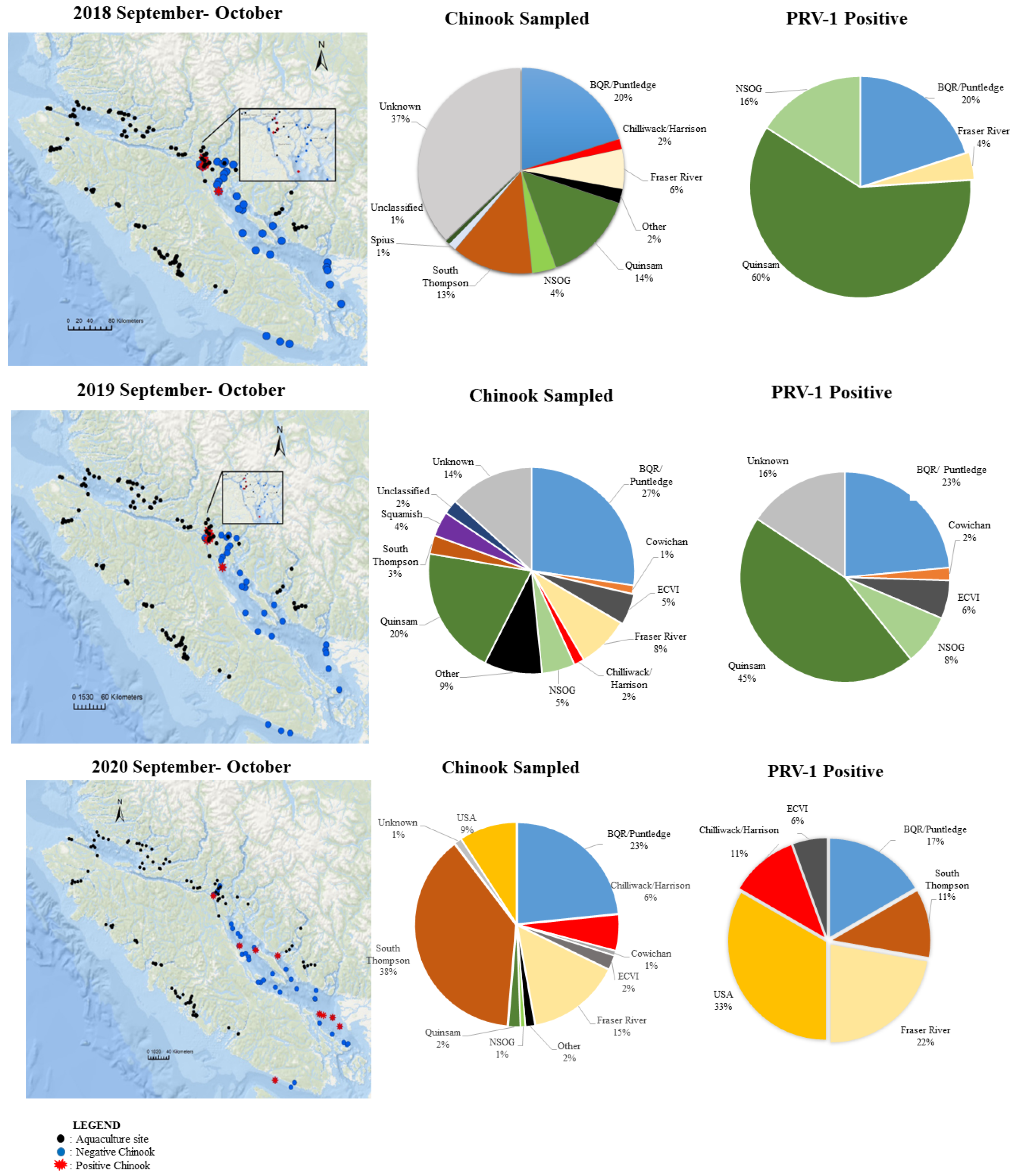
3.3.2. Stock Composition of PRV-1-Infected Juvenile Chinook Salmon
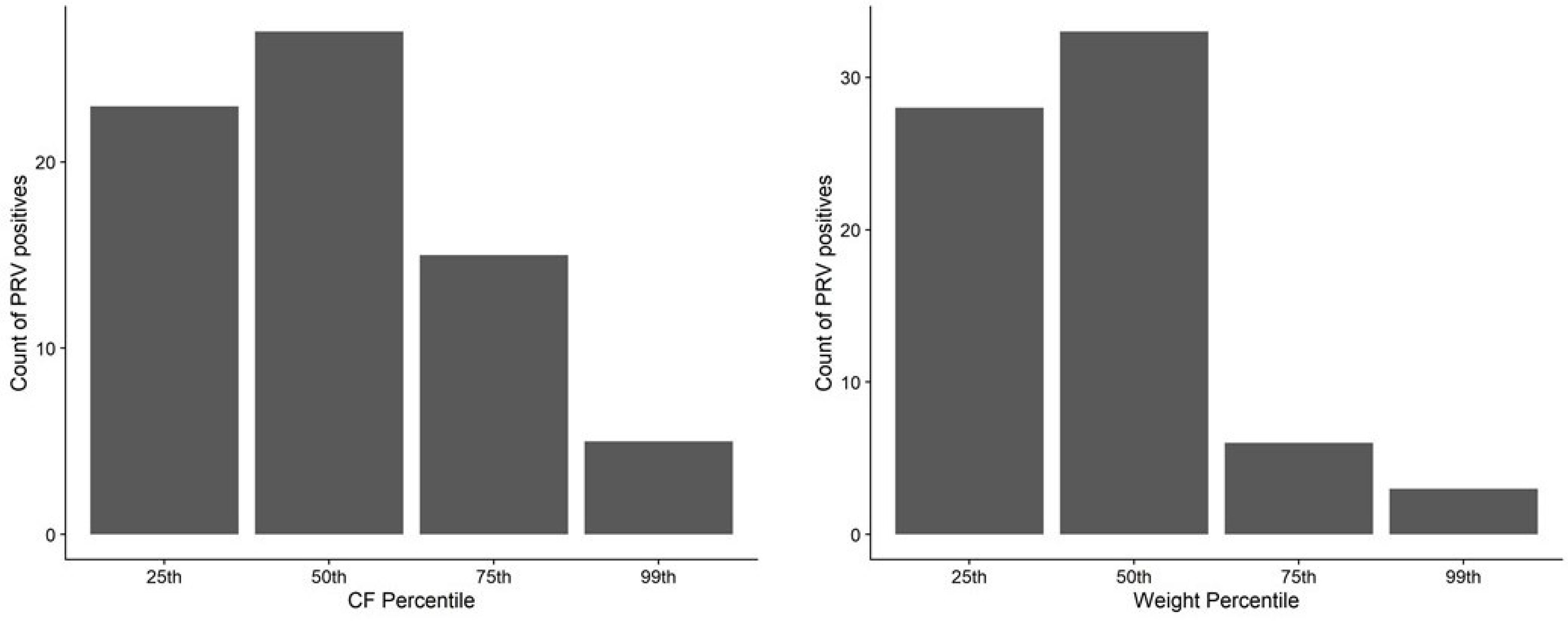
3.3.3. Relationship between Body Weight and Condition Factor and PRV-1 Infection Status in Juvenile Chinook Salmon
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Palacios, G.; Lovoll, M.; Tengs, T.; Hornig, M.; Hutchison, S.; Hui, J.; Kongtorp, R.T.; Savji, N.; Bussetti, A.V.; Solovyov, A.; et al. Heart and skeletal muscle inflammation of farmed salmon is associated with infection with a novel Reovirus. PLoS ONE 2010, 5, e11487. [Google Scholar] [CrossRef] [PubMed]
- Dhamotharan, K.; Vendramin, N.; Markussen, T.; Wessel, O.; Cuenca, A.; Nyman, I.B.; Olsen, A.B.; Tengs, T.; Dahle, M.K.; Rimstad, E. Molecular and antigenic characterization of Piscine orthoreovirus (PRV) from Rainbow Trout (Oncorhynchus mykiss). Viruses 2018, 10, 170. [Google Scholar] [CrossRef]
- Takano, T.; Nawata, A.; Sakai, T.; Matsuyama, T.; Ito, T.; Kurita, J.; Terashima, S.; Yasuike, M.; Nakamura, Y.; Fujiwara, A.; et al. Full genome sequencing and confirmation of the causative agent of Erythrocytic Inclusion Body Syndrome in Coho salmon identifies a new type of Piscine orthoreovirus. PLoS ONE 2016, 11, e0165424. [Google Scholar] [CrossRef]
- Polinski, M.P.; Vendramin, N.; Cuenca, A.; Garver, K.A. Piscine orthoreovirus: Biology and distribution in farmed and wild fish. J. Fish Dis. 2020, 43, 1331–1352. [Google Scholar] [CrossRef]
- Jia, B.B.; Delphino, M.; Awosile, B.; Hewison, T.; Whittaker, P.; Morrison, D.; Kamaitis, M.; Siah, A.; Milligan, B.; Johnson, S.C.; et al. Review of infectious agent occurrence in wild salmonids in British Columbia, Canada. J. Fish Dis. 2020, 43, 153–175. [Google Scholar] [CrossRef] [PubMed]
- Di Cicco, E.; Ferguson, H.W.; Schulze, A.D.; Kaukinen, K.H.; Li, S.; Vanderstichel, R.; Wessel, O.; Rimstad, E.; Gardner, I.A.; Hammell, K.L.; et al. Heart and skeletal muscle inflammation (HSMI) disease diagnosed on a British Columbia salmon farm through a longitudinal farm study. PLoS ONE 2017, 12, e0171471. [Google Scholar] [CrossRef] [PubMed]
- Di Cicco, E.; Ferguson, H.W.; Kaukinen, K.H.; Schulze, A.D.; Li, S.R.; Tabata, A.; Gunther, O.P.; Mordecai, G.; Suttle, C.A.; Miller, K.M. The same strain of Piscine orthoreovirus (PRV-1) is involved in the development of different, but related, diseases in Atlantic and Pacific salmon in British Columbia. Facets 2018, 3, 599–641. [Google Scholar] [CrossRef]
- Polinski, M.P.; Gross, L.A.; Marty, G.D.; Garver, K.A. Heart inflammation and piscine orthoreovirus genotype-1 in Pacific Canada Atlantic salmon net-pen farms: 2016-2019. BMC Vet. Res. 2022, 18, 306. [Google Scholar] [CrossRef]
- Mordecai, G.J.; Miller, K.M.; Bass, A.L.; Bateman, A.W.; Teffer, A.K.; Caleta, J.M.; Di Cicco, E.; Schulze, A.D.; Kaukinen, K.H.; Li, S.R.; et al. Aquaculture mediates global transmission of a viral pathogen to wild salmon. Sci. Adv. 2021, 7, eabe2592. [Google Scholar] [CrossRef]
- Nekouei, O.; Vanderstichel, R.; Kaukinen, K.H.; Thakur, K.; Ming, T.; Patterson, D.A.; Trudel, M.; Neville, C.; Miller, K.M. Comparison of infectious agents detected from hatchery and wild juvenile Coho salmon in British Columbia, 2008–2018. PLoS ONE 2019, 14, e0221956. [Google Scholar] [CrossRef]
- Marty, G.D.; Bidulka, J.; Joseph, T. Cross-sectional study of histopathology and piscine orthoreovirus during a marine production cycle of farmed Atlantic salmon (Salmo salar L.) in British Columbia, Canada. J. Fish Dis. 2020, 43, 1019–1028. [Google Scholar] [CrossRef] [PubMed]
- Mimeault, C.; Polinski, M.; Garver, K.A.; Jones, S.R.M.; Johnson, S.; Boily, F.; Malcolm, G.; Holt, K.; Burgetz, I.J.; Parsons, G.J. Assessment of the Risk to Fraser River Sockeye Salmon Due to Piscine Orthoreovirus (PRV) Transfer From Atlantic Salmon Farms in the Discovery Islands Area, British Columbia; Government of Canada: Ottawa, ON, Canada, 2019; p. 45.
- Polinski, M.P.; Haddad, C.A.; Siah, A.; Fuller, C.; Higgins, M.; Parsons, J. British Columbia freshwater salmon hatcheries demonstrate minimal contribution to piscine orthoreovirus (PRV) regional occurrence with no evidence for non-endemic strain introductions. Facets, in press.
- Hrushowy, S. A Molecular Investigation of the Dynamics of Piscine Orthoreovirus in a Wild Sockeye Salmon Community on the Central Coast of British Columbia; Simon Fraser University: Burnaby, BC, USA, 2018. [Google Scholar]
- Wilson, E.B. Probable inference, the law of succession, and statistical inference. J. Am. Stat. Assoc. 1927, 22, 209–212. [Google Scholar] [CrossRef]
- Neville, C.E.M.; Johnson, S.C.; Beacham, T.D.; Whitehouse, T.; Tadey, J.; Trudel, M. Initial estimates from an integrated study examining the residence period and migration timing of juvenile Sockeye salmon from the Fraser River through coastal water of British Columbia. NPAFC Bull 2016, 6, 45–60. [Google Scholar] [CrossRef]
- Laurin, E.; Bradshaw, J.; Hawley, L.; Gardner, I.A.; Garver, K.; Johnson, S.C.; Thakur, K.K. Importance of sample size for estimating prevalence: A case example of Infectious Hematopoietic Necrosis viral RNA detection in mixed-stock Fraser River Sockeye salmon (Oncorhynchus nerka), British Columbia, Canada. Can. J. Fish. Aquat. Sci. 2021, 78, 589–598. [Google Scholar] [CrossRef]
- Beamish, R.J.; Sweeting, R.M.; Lange, K.L.; Noakes, D.J.; Preikshot, D.; Neville, C.M. Early marine survival of Coho salmon in the Strait of Georgia declines to very low levels. Mar. Coast. Fish. 2010, 2, 424–439. [Google Scholar] [CrossRef]
- Morris, J.F.T.; Trudel, M.; Thiess, M.E.; Sweeting, R.M.; Fisher, J.; Hinton, S.A.; Fergusson, E.A.; Orsi, J.A.; Farley, E.V.; Welch, D.W. (Eds.) Stock-Specific Migrations of Juvenile Coho Salmon Derived from Coded-Wire Tag Recoveries on the Continental Shelf of Western North America; American Fisheries Society: Bethesda, MD, USA, 2007; Volume Symposium 57, p. 247. [Google Scholar]
- Preikshot, D.; Beamish, R.J.; Sweeting, R.M.; Neville, C.M.; Beacham, T.D. The residence time of Jjuvenile Fraser River Sockeye salmon in the Strait of Georgia. Mar. Coast. Fish. 2012, 4, 438–449. [Google Scholar] [CrossRef]
- Quinn, T.P.; Myers, K.W. Anadromy and the marine migrations of Pacific salmon and trout: Rounsefell revisited. Rev. Fish Biol. Fish. 2004, 14, 421–442. [Google Scholar] [CrossRef]
- Trudel, M.; Fisher, J.; Orsi, J.A.; Morris, J.F.T.; Thiess, M.E.; Sweeting, R.M.; Hinton, S.; Fergusson, E.A.; Welch, D.W. Distribution and migration of juvenile Chinook salmon derived from coded wire tag recoveries along the continental shelf of Western North America. Trans. Am. Fish. Soc. 2009, 138, 1369–1391. [Google Scholar] [CrossRef]
- Tucker, S.; Trudel, M.; Welch, D.W.; Candy, J.R.; Morris, J.F.T.; Thiess, M.E.; Wallace, C.; Teel, D.J.; Crawford, W.; Farley, E.V.; et al. Seasonal Stock-Specific Migrations of juvenile Sockeye salmon along the west coast of North America: Implications for growth. Trans. Am. Fish. Soc. 2009, 138, 1458–1480. [Google Scholar] [CrossRef]
- Zhao, J.; Vendramin, N.; Cuenca, A.; Polinski, M.; Hawley, L.M.; Garver, K.A. Pan-Piscine orthoreovirus (PRV) detection using reverse transcription quantitative PCR. Pathogens 2021, 10, 1548. [Google Scholar] [CrossRef]
- Polinski, M.P.; Marty, G.D.; Snyman, H.N.; Garver, K.A. Piscine orthoreovirus demonstrates high infectivity but low virulence in Atlantic salmon of Pacific Canada. Sci. Rep. 2019, 9, 3297. [Google Scholar] [CrossRef] [PubMed]
- Garver, K.A.; Johnson, S.C.; Polinski, M.P.; Bradshaw, J.C.; Marty, G.D.; Snyman, H.N.; Morrison, D.B.; Richard, J. Piscine Orthoreovirus from western North America is transmissible to Atlantic salmon and Sockeye salmon but fails to cause Heart and Skeletal Muscle Inflammation. PLoS ONE 2016, 11, e0146229. [Google Scholar] [CrossRef] [PubMed]
- Beacham, T.D.; Candy, J.R.; Jonsen, K.L.; Supernault, J.; Wetklo, M.; Deng, L.T.; Miller, K.M.; Withler, R.E.; Varnavskaya, N. Estimation of stock composition and individual identification of Chinook salmon across the Pacific Rim by use of microsatellite variation. Trans. Am. Fish. Soc. 2006, 135, 861–888. [Google Scholar] [CrossRef]
- Tucker, S.; Trudel, M.; Welch, D.W.; Candy, J.R.; Morris, J.F.T.; Thiess, M.E.; Wallace, C.; Beacham, T.D. Annual coastal migration of juvenile Chinook salmon: Static stock-specific patterns in a highly dynamic ocean. Mar. Ecol. Prog. Ser. 2012, 449, 245–262. [Google Scholar] [CrossRef]
- Purcell, M.K.; Powers, R.L.; Evered, J.; Kerwin, J.; Meyers, T.R.; Stewart, B.; Winton, J.R. Molecular testing of adult Pacific salmon and trout (Oncorhynchus spp.) for several RNA viruses demonstrates widespread distribution of piscine orthoreovirus in Alaska and Washington. J. Fish Dis. 2018, 41, 347–355. [Google Scholar] [CrossRef]
- Morton, A.; Routledge, R.; Hrushowy, S.; Kibenge, M.; Kibenge, F. The effect of exposure to farmed salmon on piscine orthoreovirus infection and fitness in wild Pacific salmon in British Columbia, Canada. PLoS ONE 2017, 12, e0188793, Erratum in PLoS ONE 2021, 16, e0248912; PLoS ONE 2023, 18, e0282687. [Google Scholar] [CrossRef] [PubMed]
- Deeg, C.M.; Kanzeparova, A.N.; Somov, A.A.; Esenkulova, S.; Di Cicco, E.; Kaukinen, K.H.; Tabata, A.; Ming, T.J.; Li, S.R.; Mordecai, G.; et al. Way out there: Pathogens, health, and condition of overwintering salmon in the Gulf of Alaska. Facets 2022, 7, 247–285. [Google Scholar] [CrossRef]
- Marty, G.D.; Morrison, D.B.; Bidulka, J.; Joseph, T.; Siah, A. Piscine reovirus in wild and farmed salmonids in British Columbia, Canada: 1974–2013. J. Fish Dis. 2015, 38, 713–728. [Google Scholar] [CrossRef]
- Saksida, S.M.; Marty, G.D.; Jones, S.R.M.; Manchester, H.A.; Diamond, C.L.; Bidulka, J.; St-Hilaire, S. Parasites and hepatic lesions among pink salmon, Oncorhynchus gorbuscha (Walbaum), during early seawater residence. J. Fish Dis. 2012, 35, 137–151. [Google Scholar] [CrossRef]
- Jeffries, K.M.; Hinch, S.G.; Gale, M.K.; Clark, T.D.; Lotto, A.G.; Casselman, M.T.; Li, S.R.; Rechisky, E.L.; Porter, A.D.; Welch, D.W.; et al. Immune response genes and pathogen presence predict migration survival in wild salmon smolts. Mol. Ecol. 2014, 23, 5803–5815. [Google Scholar] [CrossRef]
- Stevenson, C.F.; Bass, A.L.; Furey, N.; Miller, K.M.; Li, S.R.; Rechisky, E.L.; Porter, A.D.; Welch, D.W.; Hinch, S.G. Infectious agents and gene expression differ between Sockeye salmon (Oncorhynchus nerka) smolt age classes but do not predict migration survival. Can. J. Fish. Aquat. Sci. 2020, 77, 484–495. [Google Scholar] [CrossRef]
- Nekouei, O.; Vanderstichel, R.; Ming, T.B.; Kaukinen, K.H.; Thakur, K.; Tabata, A.; Laurin, E.; Tucker, S.; Beacham, T.D.; Miller, K.M. Detection and assessment of the distribution of infectious agents in juvenile Fraser River Sockeye salmon, Canada, in 2012 and 2013. Front. Microbiol. 2018, 9, 3221. [Google Scholar] [CrossRef]
- Rechisky, E.L.; Porter, A.D.; Johnston, S.D.; Stevenson, C.F.; Hinch, S.G.; Hunt, B.P.V.; Welch, D.W. Exposure time of wild, juvenile Sockeye salmon to open-net-pen Atlantic Salmon farms in British Columbia, Canada. North Am. J. Fish. Manag. 2021, 41, 650–660. [Google Scholar] [CrossRef]
- Thakur, K.K.; Vanderstichel, R.; Li, S.R.; Laurin, E.; Tucker, S.; Neville, C.; Tabata, A.; Miller, K.M. A comparison of infectious agents between hatchery-enhanced and wild out-migrating juvenile Chinook salmon (Oncorhynchus tshawytscha) from Cowichan River, British Columbia. Facets 2018, 3, 695–721. [Google Scholar] [CrossRef]
- Tucker, S.; Li, S.R.; Kaukinen, K.H.; Patterson, D.A.; Miller, K.M. Distinct seasonal infectious agent profiles in life-history variants of juvenile Fraser River Chinook salmon: An application of high-throughput genomic screening. PLoS ONE 2018, 13, e0195472. [Google Scholar] [CrossRef] [PubMed]
- Laurin, E.; Jaramillo, D.; Vanderstichel, R.; Ferguson, H.; Kaukinen, K.H.; Schulze, A.D.; Keith, I.R.; Gardner, I.A.; Miller, K.M. Histopathological and novel high-throughput molecular monitoring data from farmed salmon (Salmo salar and Oncorhynchus spp.) in British Columbia, Canada, from 2011-2013. Aquaculture 2019, 499, 220–234. [Google Scholar] [CrossRef]
- Bass, A.L.; Bateman, A.W.; Connors, B.M.; Staton, B.A.; Rondeau, E.B.; Mordecai, G.J.; Teffer, A.K.; Kaukinen, K.H.; Li, S.R.; Tabata, A.M.; et al. Identification of infectious agents in early marine Chinook and Coho salmon associated with cohort survival. Facets 2022, 7, 742–773. [Google Scholar] [CrossRef]
- Garver, K.A.; Marty, G.D.; Cockburn, S.N.; Richard, J.; Hawley, L.M.; Muller, A.; Thompson, R.L.; Purcell, M.K.; Saksida, S. Piscine reovirus, but not Jaundice Syndrome, was transmissible to Chinook salmon, Oncorhynchus tshawytscha (Walbaum), Sockeye salmon, Oncorhynchus nerka (Walbaum), and Atlantic salmon, Salmo salar L. J. Fish Dis. 2016, 39, 117–128. [Google Scholar] [CrossRef]
- Purcell, M.K.; Powers, R.L.; Taksdal, T.; McKenney, D.; Conway, C.M.; Bilott, D.G.; Polinski, M.; Garver, K.; Winton, J. Consequences of Piscine orthoreovirus genotype 1 (PRV-1) infections in Chinook salmon (Oncorhynchus tshawytscha), Coho salmon (O. kisutch) and Rainbow trout (O. mykiss). J. Fish Dis. 2020, 43, 719–728. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Polinski, M.P.; Morrison, P.R.; Brauner, C.J.; Farrell, A.P.; Garver, K.A. High-load reovirus infections do not imply physiological impairment in salmon. Front. Physiol. 2019, 10, 114. [Google Scholar] [CrossRef]
- Polinski, M.P.; Zhang, Y.F.; Morrison, P.R.; Marty, G.D.; Brauner, C.J.; Farrell, A.P.; Garver, K.A. Innate antiviral defense demonstrates high energetic efficiency in a bony fish. BMC Biol. 2021, 19, 138. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Life Stage | Survey # | Sample Month | Sample Region/Description | Sample Size (N) | No. of Individuals Testing Positive | Prevalence (%) | Lower 95% CI | Upper 95% CI |
|---|---|---|---|---|---|---|---|---|---|
| Chinook | Freshwater pre-release | 2015-02 | June | Quinsam | 25 | 0 | 0.0 | 0.00 | 13.32 |
| 2016-02 | June | Quinsam | 15 | 0 | 0.0 | 0.00 | 20.39 | ||
| 2018-B | March | Eby St-Terrace | 25 | 0 | 0.0 | 0.00 | 13.32 | ||
| 2018-D | March | Spius creek, Coldwater | 25 | 0 | 0.0 | 0.00 | 13.32 | ||
| 2018-E | March | Spius creek | 24 | 0 | 0.0 | 0.00 | 13.80 | ||
| 2018-F | April | Inch creek, Chilko | 25 | 0 | 0.0 | 0.00 | 13.32 | ||
| 2018-G | April | Chehalis, Chilko | 25 | 0 | 0.0 | 0.00 | 13.32 | ||
| 2018-H | April | Chehalis-Summer red | 25 | 0 | 0.0 | 0.00 | 13.32 | ||
| 2018-J | April | Tenderfoot, Namquam | 25 | 0 | 0.0 | 0.00 | 13.32 | ||
| 2018-K | April | Tenderfoot, Ashley | 25 | 0 | 0.0 | 0.00 | 13.32 | ||
| 2018-O | April | Nitnat | 24 | 0 | 0.0 | 0.00 | 13.80 | ||
| Total: | 263 | 0 | 0.0 | - | - | ||||
| Juvenile | 2012-01 | May | SOG | 50 | 0 | 0.0 | 0.00 | 7.13 | |
| 2012-02 | June | SOG | 42 | 0 | 0.0 | 0.00 | 8.38 | ||
| 2015-05 | June | SOG | 11 | 0 | 0.0 | 0.00 | 25.88 | ||
| 2015-16 | May–June | SOG | 218 | 4 | 1.8 | 0.72 | 4.62 | ||
| 2015-19 | October | WCVI | 87 | 11 | 12.6 | 7.21 | 21.24 | ||
| 2016-07 | May | SOG | 78 | 1 | 1.3 | 0.23 | 6.91 | ||
| 2016-08 | June | SOG | 45 | 2 | 4.4 | 1.23 | 14.83 | ||
| 2016-12 | July | SOG | 25 | 0 | 0.0 | 0.00 | 13.32 | ||
| 2018-04 | September–October | SOG | 318 | 26 | 8.2 | 5.64 | 11.71 | ||
| 2019-02 | June | SOG | 179 | 1 | 0.6 | 0.10 | 3.10 | ||
| 2019-03 | September–October | SOG | 281 | 51 | 18.1 | 14.08 | 23.08 | ||
| 2020-02 | September–October | SOG | 260 | 22 | 8.5 | 5.65 | 1.48 | ||
| Total: | 1594 | 118 | 7.4 | - | - | ||||
| Adult Returns | 2016-161 | November | Puntledge | 25 | 1 | 4.0 | 0.71 | 19.54 | |
| 2016-170 | November | Omega, Robertson | 40 | 3 | 7.5 | 2.58 | 19.86 | ||
| 2017-110 | August | Tenderfoot, Chekamus | 26 | 0 | 0.0 | 0.00 | 12.87 | ||
| 2017-121 | August | Tenderfoot | 8 | 0 | 0.0 | 0.00 | 32.44 | ||
| 2017-143 | September | Chilko | 25 | 0 | 0.0 | 0.00 | 13.32 | ||
| 2017-163 | October | Puntledge | 24 | 0 | 0.0 | 0.00 | 13.80 | ||
| 2017-167 | October | Robertson | 36 | 2 | 5.6 | 1.54 | 18.14 | ||
| 2017-181 | November | Puntledge | 58 | 1 | 1.7 | 0.31 | 9.14 | ||
| Total: | 242 | 7 | 2.9 | - | - | ||||
| Total Chinook: | 2099 | 125 | 6.0 | 5.02 | 7.05 | ||||
| Chum | Juvenile | 2011-07 | March–May | WCVI | 60 | 0 | 0.0 | 0.00 | 6.02 |
| 2012-01 | May | SOG | 25 | 0 | 0.0 | 0.00 | 13.32 | ||
| 2012-02 | June | SOG | 50 | 0 | 0.0 | 0.00 | 7.13 | ||
| Total Chum: | 135 | 0 | 0.0 | 0.00 | 2.77 | ||||
| Coho | Freshwater | 2018-A | March | Kitimat | 25 | 0 | 0.0 | 0.00 | 13.32 |
| 2018-C | March | Oilfield, Prince Rupert | 25 | 0 | 0.0 | 0.00 | 13.32 | ||
| 2018-I | April | Inch creek, Stave | 25 | 0 | 0.0 | 0.00 | 13.32 | ||
| 2018-L | April | Quinsam | 25 | 0 | 0.0 | 0.00 | 13.32 | ||
| 2018-M | April | Fanny Bay | 25 | 0 | 0.0 | 0.00 | 13.32 | ||
| 2018-N | April | Nitnat | 25 | 0 | 0.0 | 0.00 | 13.32 | ||
| 2018-Q | April | Chilliwack, Coldwater | 25 | 0 | 0.0 | 0.00 | 13.32 | ||
| 2018-R | April | Quatse | 25 | 0 | 0.0 | 0.00 | 13.32 | ||
| Total: | 200 | 0 | 0.0 | - | - | ||||
| Juvenile | 2015-16 | May | SOG | 1 | 0 | 0.0 | 0.00 | 79.35 | |
| 2016-07 | July | SOG | 1 | 1 | 100 | 20.65 | 100.00 | ||
| 2016-08 | June | SOG | 2 | 0 | 0.0 | 0.00 | 65.76 | ||
| 2018-04 | September | SOG | 152 | 1 | 0.7 | 0.12 | 3.63 | ||
| 2019-02 | June | SOG | 142 | 4 | 2.8 | 1.10 | 7.02 | ||
| 2019-03 | September | SOG | 128 | 3 | 2.3 | 0.80 | 6.66 | ||
| 2020-02 | September | SOG | 169 | 1 | 0.6 | 0.10 | 3.28 | ||
| Total: | 595 | 10 | 1.7 | - | - | ||||
| Adult Returns | 2017-195 | August | Oilfield | 15 | 0 | 0.0 | 0.00 | 20.39 | |
| 2017-191 | September | Kitimat | 50 | 2 | 4.0 | 1.10 | 13.46 | ||
| 2017-192 | September | Nitnat | 25 | 0 | 0.0 | 0.00 | 13.32 | ||
| 2017-212 | September | Tenderfoot | 50 | 1 | 2.0 | 0.35 | 10.50 | ||
| 2018-203 | September | Kitimat | 50 | 4 | 8.0 | 3.15 | 18.84 | ||
| 2018-209 | September | Nitnat | 25 | 0 | 0.0 | 0.00 | 13.32 | ||
| Total: | 215 | 7 | 3.3 | - | - | ||||
| Total Coho: | 1010 | 17 | 1.7 | 1.05 | 2.68 | ||||
| Pink | Juvenile | 2012-01 | May | SOG | 20 | 0 | 0.0 | 0.00 | 16.11 |
| 2012-02 | June | SOG | 50 | 0 | 0.0 | 0.00 | 7.13 | ||
| Total Pink: | 70 | 0 | 0.0 | 0.00 | 5.20 | ||||
| Sockeye | Juvenile | 2014-RST | April–May | LFR | 112 | 0 | 0.0 | 0.00 | 3.32 |
| 2016-RST | April–May | LFR | 232 | 0 | 0.0 | 0.00 | 1.63 | ||
| 2012-01 | May | DISC | 135 | 0 | 0.0 | 0.00 | 2.77 | ||
| 2014-04 | May–June | DISC | 127 | 0 | 0.0 | 0.00 | 2.94 | ||
| 2015-05 | May–June | DISC | 43 | 0 | 0.0 | 0.00 | 8.20 | ||
| 2016-07 | May–June | DISC | 68 | 0 | 0.0 | 0.00 | 5.35 | ||
| Total Sockeye: | 717 | 0 | 0.0 | 0.00 | 0.53 | ||||
| GRAND TOTAL: | 4031 | 142 | 3.5 | 3.00 | 4.14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turcotte, L.D.M.; Bradshaw, J.C.; Polinski, M.P.; Johnson, S.C. Piscine orthoreovirus Genotype-1 (PRV-1) in Wild Pacific Salmon of British Columbia, Canada: 2011–2020. Fishes 2023, 8, 252. https://doi.org/10.3390/fishes8050252
Turcotte LDM, Bradshaw JC, Polinski MP, Johnson SC. Piscine orthoreovirus Genotype-1 (PRV-1) in Wild Pacific Salmon of British Columbia, Canada: 2011–2020. Fishes. 2023; 8(5):252. https://doi.org/10.3390/fishes8050252
Chicago/Turabian StyleTurcotte, Lenora D. M., Julia C. Bradshaw, Mark P. Polinski, and Stewart C. Johnson. 2023. "Piscine orthoreovirus Genotype-1 (PRV-1) in Wild Pacific Salmon of British Columbia, Canada: 2011–2020" Fishes 8, no. 5: 252. https://doi.org/10.3390/fishes8050252
APA StyleTurcotte, L. D. M., Bradshaw, J. C., Polinski, M. P., & Johnson, S. C. (2023). Piscine orthoreovirus Genotype-1 (PRV-1) in Wild Pacific Salmon of British Columbia, Canada: 2011–2020. Fishes, 8(5), 252. https://doi.org/10.3390/fishes8050252