


Effects of Seasonal Photoperiod on Growth, Lipid Metabolism, and Antioxidant Response in the Huanghe Carp (Cyprinus carpio haematopterus)

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Liver Tissue Biochemical Parameters
2.3. Transcriptome Assembly and Differential Expression Gene (DEGs) Analysis
2.4. RNA Extraction and Quantitative Real-Time PCR
2.5. Correlation Analysis
2.6. Statistical Analysis
3. Results
3.1. Effects of Seasonal Photoperiod on the Growth Performance of Huanghe Carp
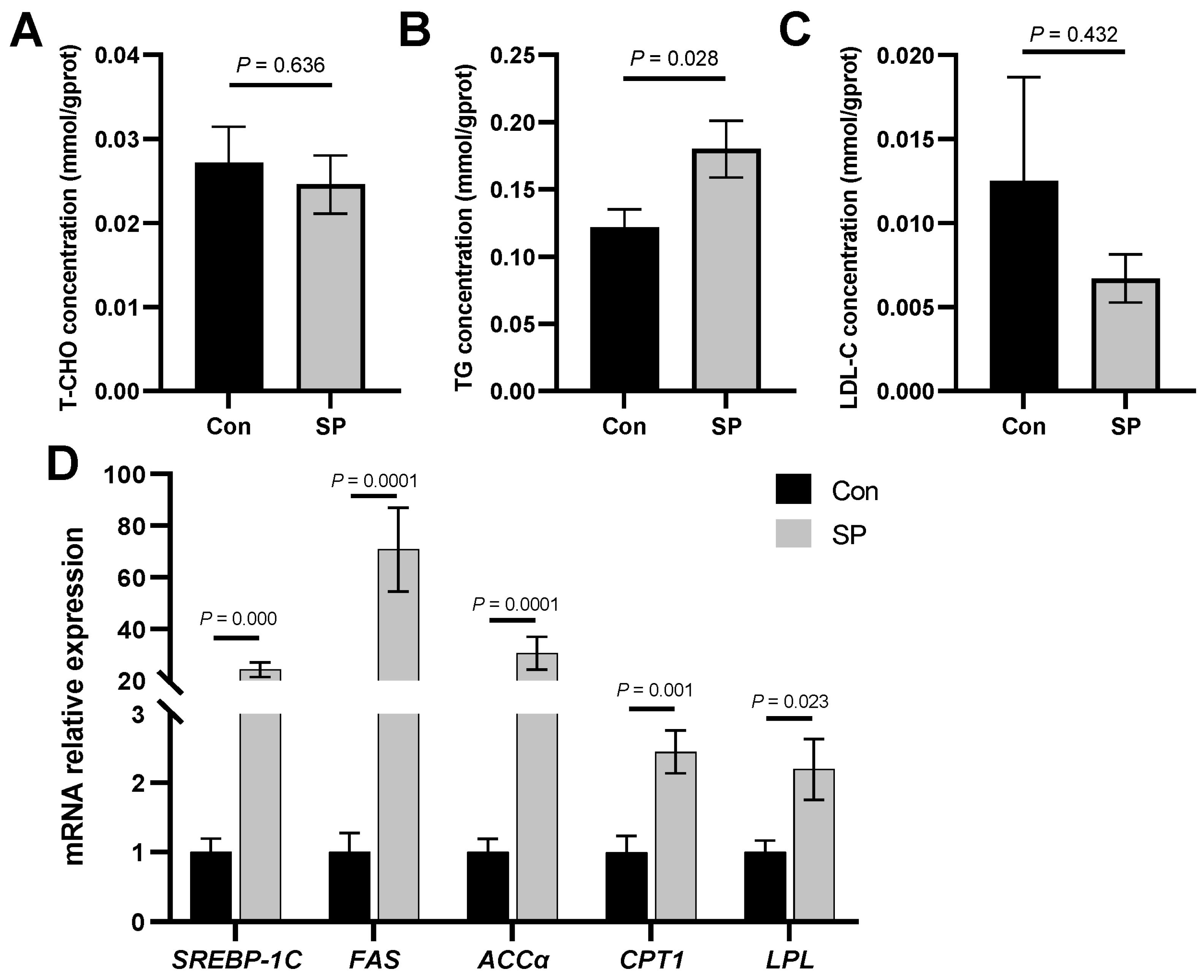
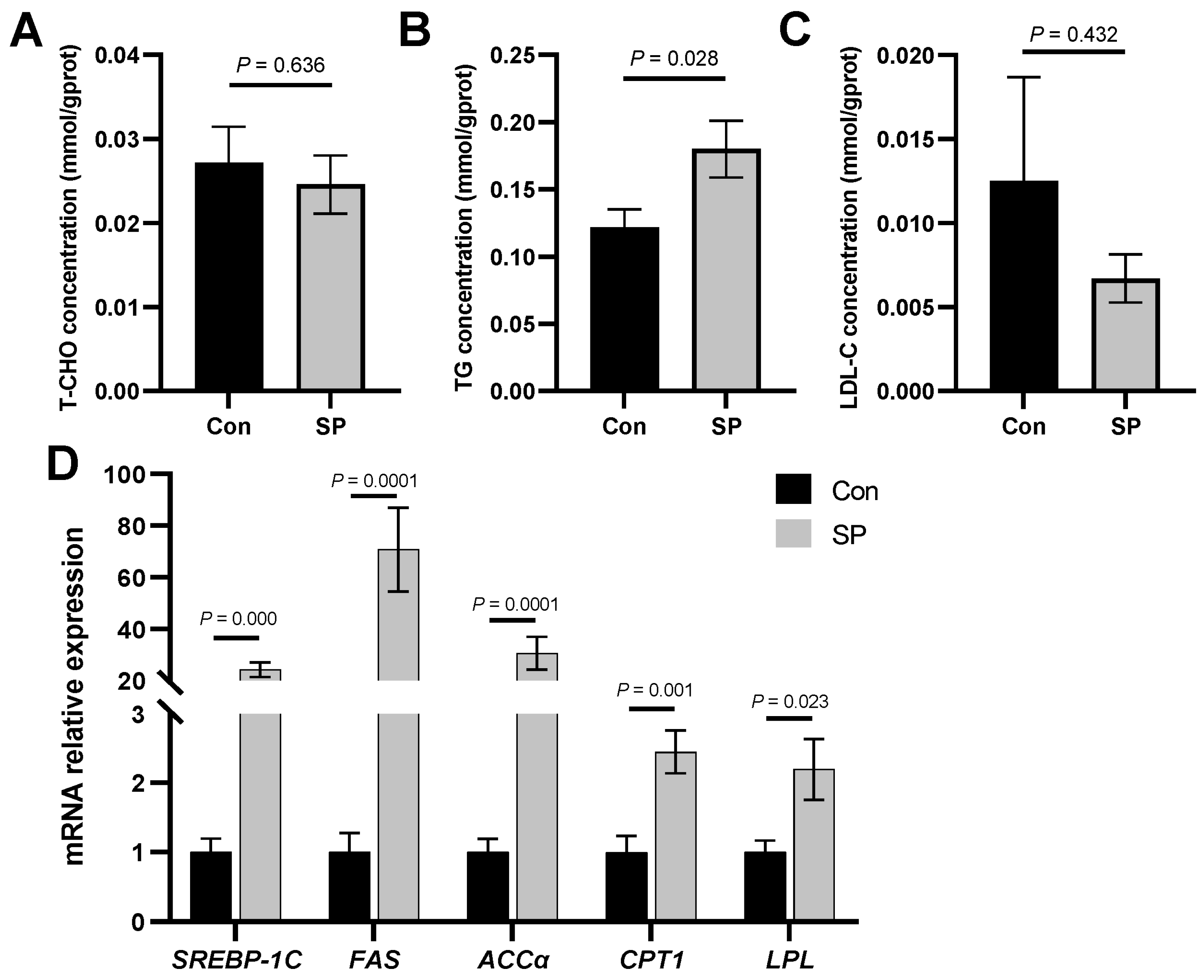
3.2. Alterations in Lipid Metabolism during Seasonal Photoperiods
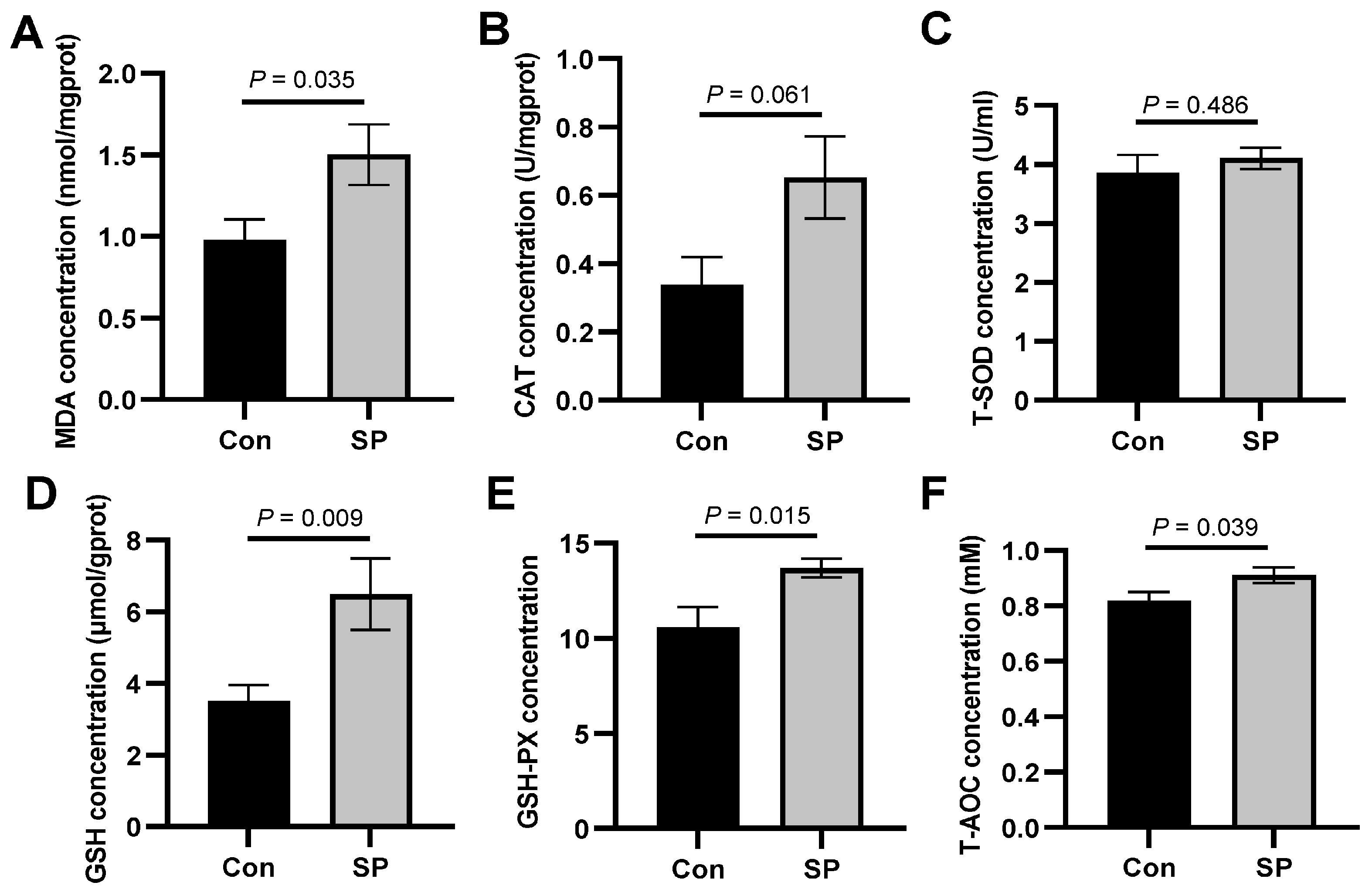
3.3. Influence of Photoperiod on Oxidative Stress in the Liver of the Huanghe Carp
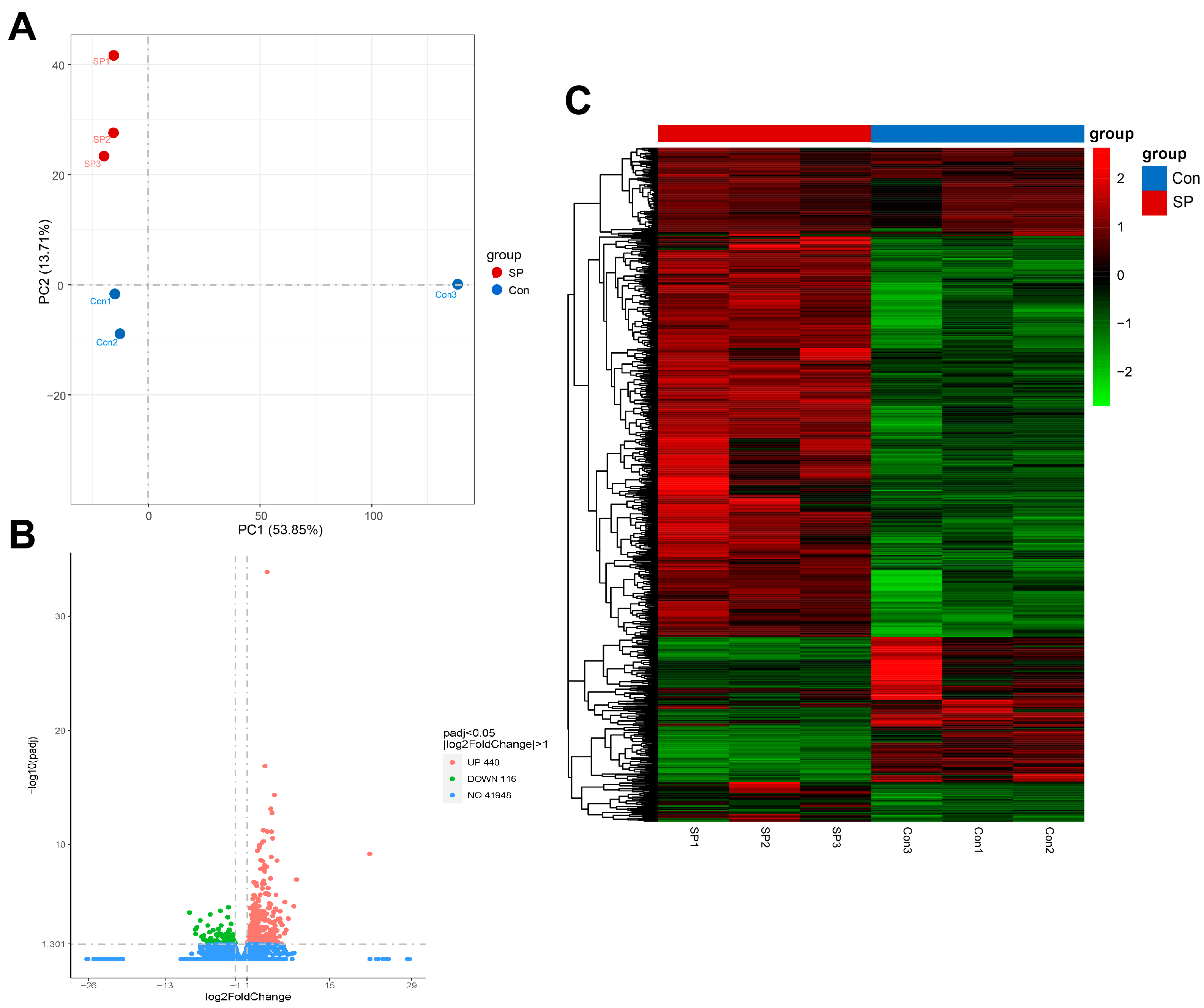
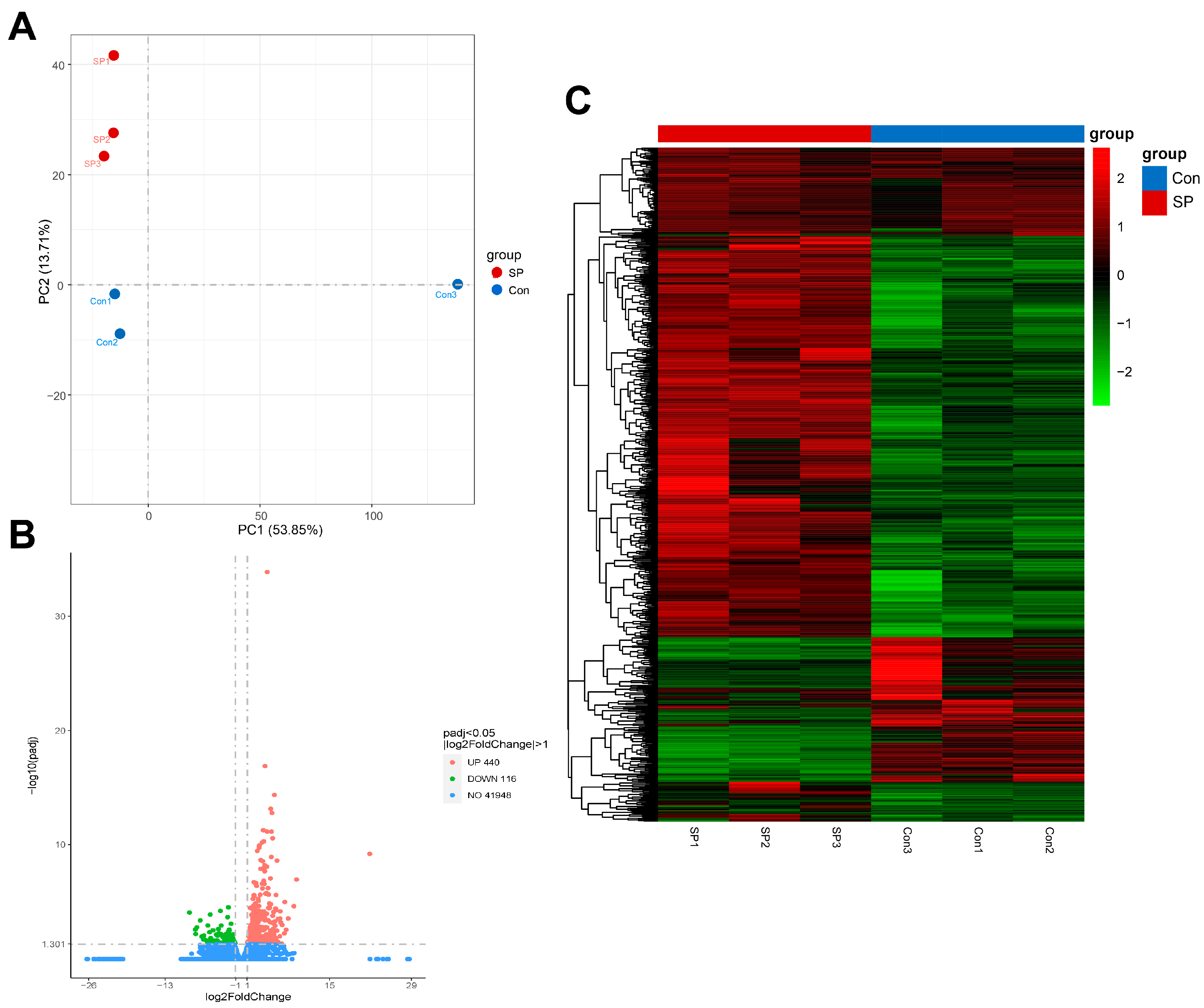
3.4. Quality of Library Sequencing and Differential Gene Expression in Liver
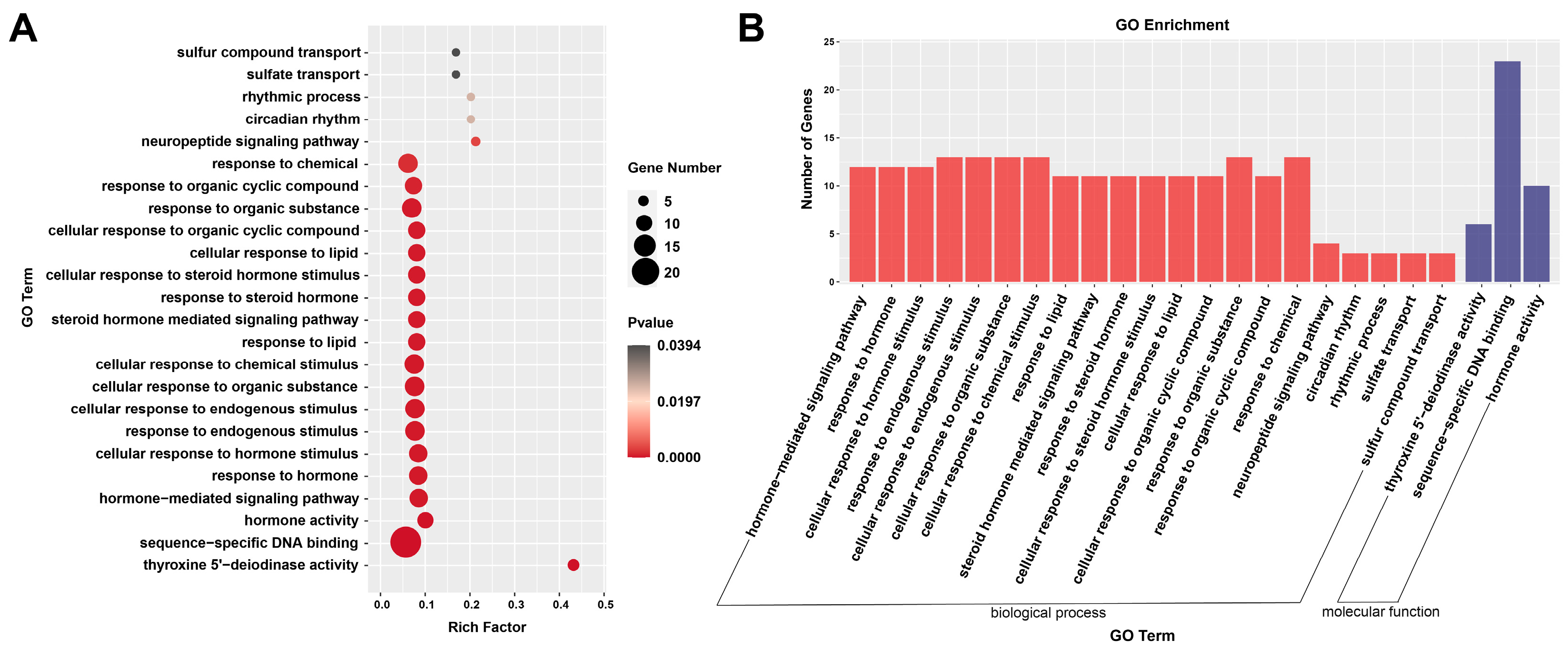
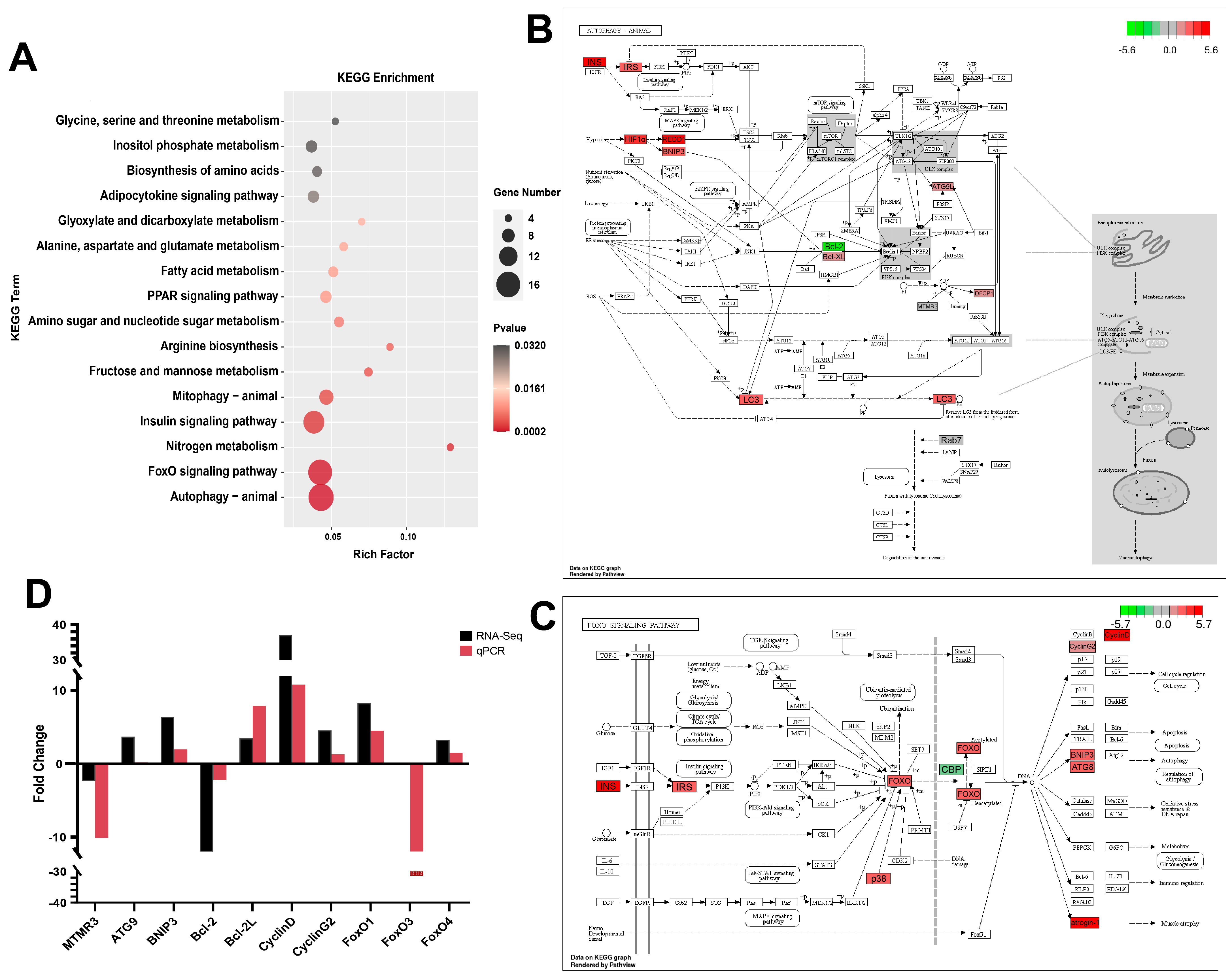
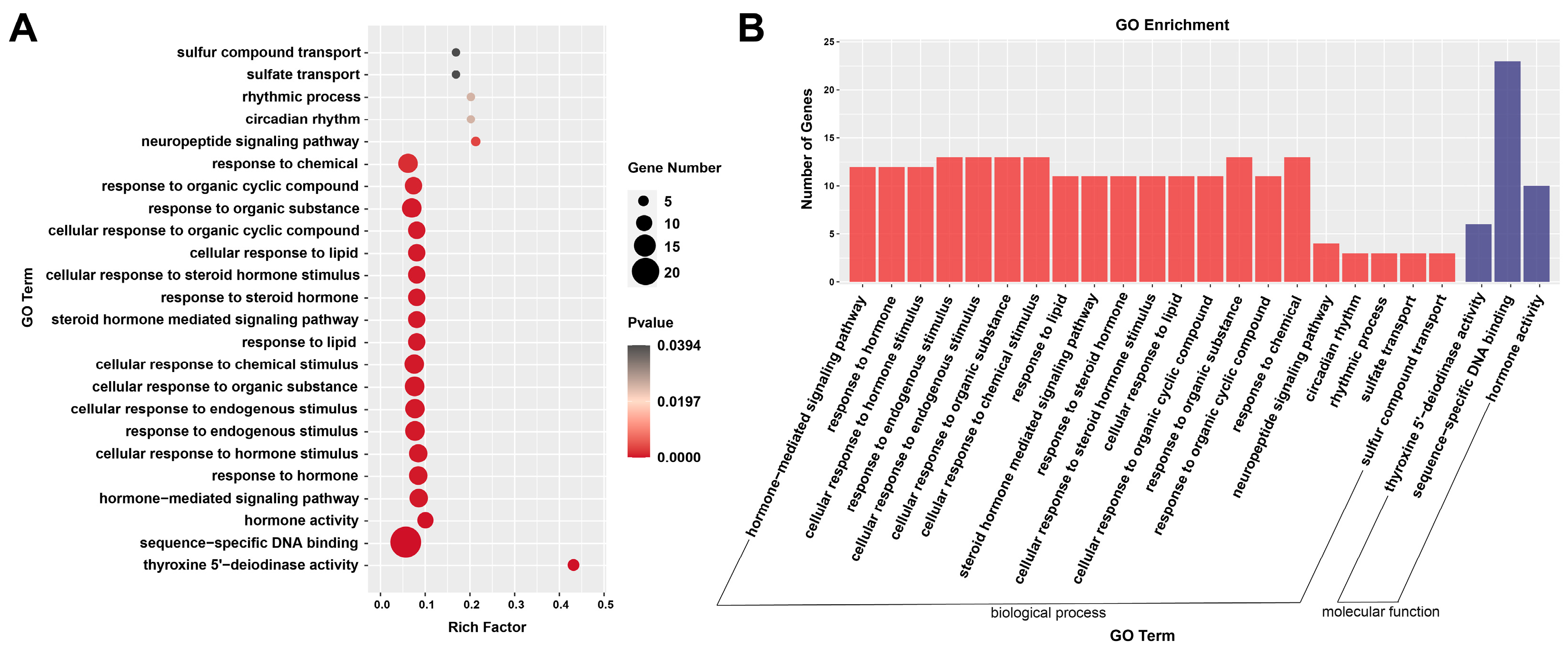
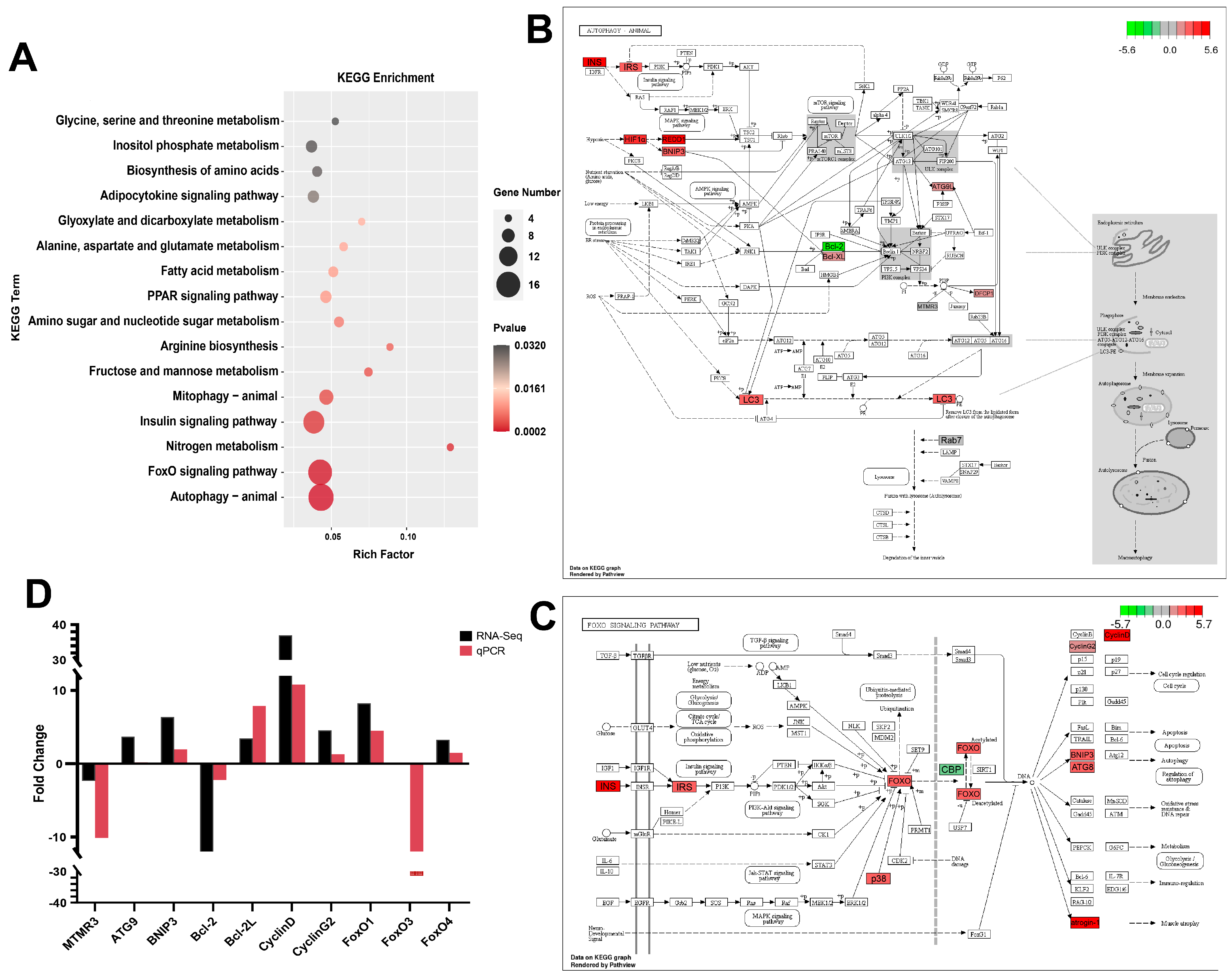
3.5. Functional Analysis by GO Enrichment and KEGG Enrichment
3.6. Data Validation by qPCR
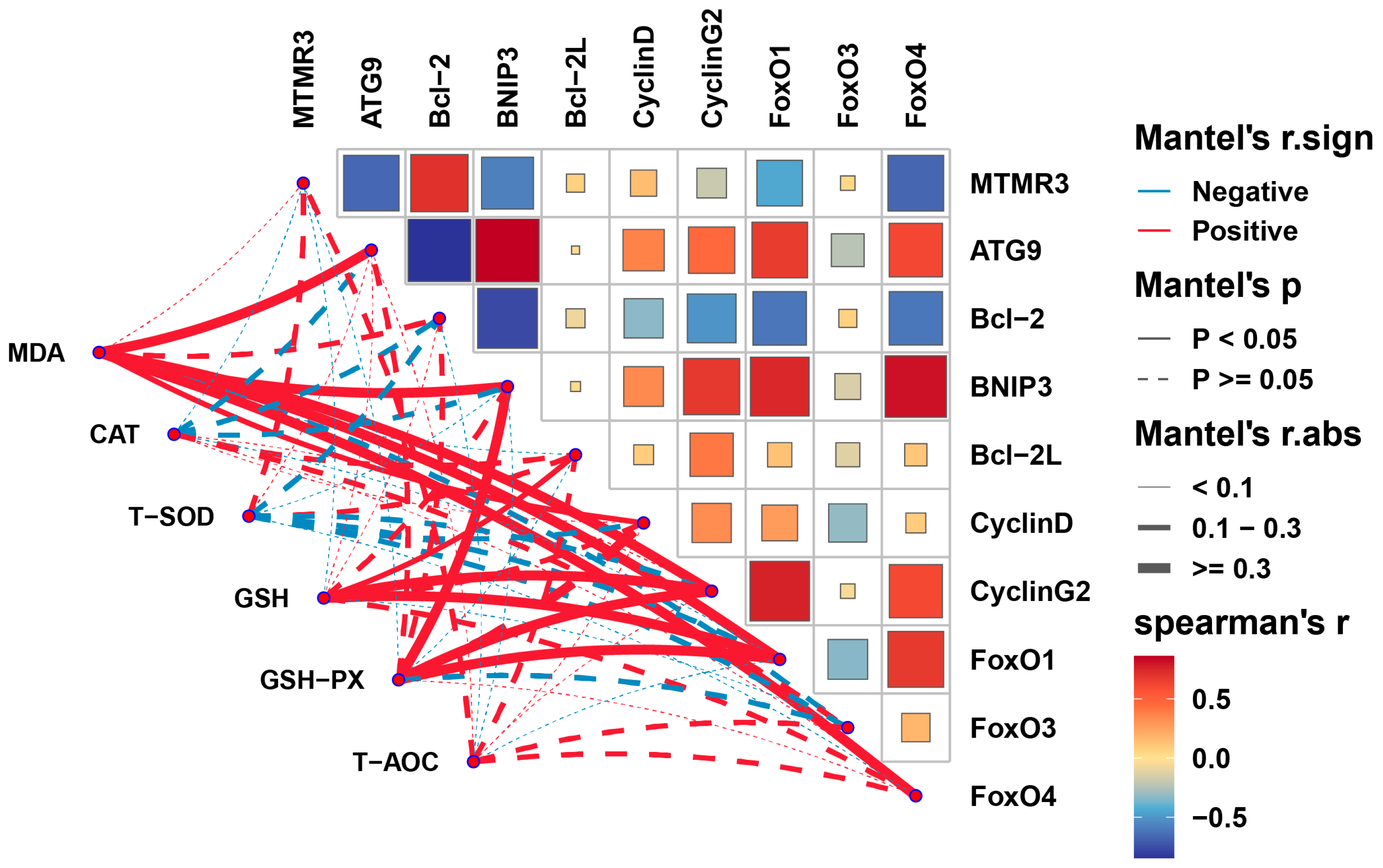
3.7. Correlation Analysis between Antioxidant Parameters and Key Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Villamizar, N.; Blanco-Vives, B.; Migaud, H.; Davie, A.; Carboni, S.; Sánchez-Vázquez, F.J. Effects of light during early larval development of some aquacultured teleosts: A review. Aquaculture 2011, 315, 86–94. [Google Scholar] [CrossRef]
- Reebs, S.G. Sensory Systems, Perception, and Learning| Circadian Rhythms in Fish. In Encyclopedia of Fish Physiology; Farrell, A.P., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 736–743. [Google Scholar]
- Bromage, N.; Porter, M.; Randall, C. The environmental regulation of maturation in farmed finfish with special reference to the role of photoperiod and melatonin. Aquaculture 2001, 197, 63–98. [Google Scholar] [CrossRef]
- Chen, S.; Liu, J.; Shi, C.; Migaud, H.; Ye, Y.; Song, C.; Mu, C.; Ren, Z.; Wang, C. Effect of photoperiod on growth, survival, and lipid metabolism of mud crab Scylla paramamosain juveniles. Aquaculture 2023, 567, 739279. [Google Scholar] [CrossRef]
- Fu, X.; Zou, Z.; Zhu, J.; Xiao, W.; Li, D.; Yu, J.; Chen, B.; Yang, H. Effects of different photoperiods on growth performance, daily rhythm of growth axis-related genes, and hormones in Nile tilapia (Oreochromis niloticus). Aquaculture 2022, 553, 738071. [Google Scholar] [CrossRef]
- Wei, H.; Cai, W.J.; Liu, H.K.; Han, D.; Zhu, X.M.; Yang, Y.X.; Jin, J.Y.; Xie, S.Q. Effects of photoperiod on growth, lipid metabolism and oxidative stress of juvenile gibel carp (Carassius auratus). J. Photochem. Photobiol. B Biol. 2019, 198, 111552. [Google Scholar] [CrossRef] [PubMed]
- Fukada, H.; Yabuki, H.; Miura, C.; Miura, T.; Kato, K. Regulation of lipid metabolism by water temperature and photoperiod in yellowtail Seriola quinqueradiata. Fish Sci. 2023, 89, 1–12. [Google Scholar] [CrossRef]
- Malinovskyi, O.; Rahimnejad, S.; Stejskal, V.; Boňko, D.; Stara, A.; Velíšek, J.; Policar, T. Effects of different photoperiods on growth performance and health status of largemouth bass (Micropterus salmoides) juveniles. Aquaculture 2021, 548, 737631. [Google Scholar] [CrossRef]
- Hosaka, T.; Biggs, W.H.; Tieu, D.; Boyer, A.D.; Varki, N.M.; Cavenee, W.K.; Arden, K.C. Disruption of forkhead transcription factor (FOXO) family members in mice reveals their functional diversification. Proc. Natl. Acad. Sci. USA 2004, 101, 2975–2980. [Google Scholar] [CrossRef]
- Barthel, A.; Schmoll, D.; Unterman, T.G. FoxO proteins in insulin action and metabolism. Trends Endocrinol. Metab. 2005, 16, 183–189. [Google Scholar] [CrossRef]
- Chen, G.; Pang, M.; Yu, X.; Wang, J.; Tong, J. Transcriptome sequencing provides insights into the mechanism of hypoxia adaption in bighead carp (Hypophthalmichthys nobilis). Comp. Biochem. Phys. D Genom. Proteom. 2021, 40, 100891. [Google Scholar] [CrossRef]
- Wu, S.; Huang, J.; Li, Y.; Pan, Y. Dynamic and systemic regulatory mechanisms in rainbow trout (Oncorhynchus mykiss) in response to acute hypoxia and reoxygenation stress. Aquaculture 2023, 572, 739540. [Google Scholar] [CrossRef]
- Blödorn, E.B.; Domingues, W.B.; Nunes, L.S.; Komninou, E.R.; Pinhal, D.; Campos, V.F. MicroRNA roles and their potential use as selection tool to cold tolerance of domesticated teleostean species: A systematic review. Aquaculture 2021, 540, 736747. [Google Scholar] [CrossRef]
- Shang, X.; Xu, W.; Zhang, Y.; Sun, Q.; Li, Z.; Geng, L.; Teng, X. Transcriptome analysis revealed the mechanism of Luciobarbus capito (L. capito) adapting high salinity: Antioxidant capacity, heat shock proteins, immunity. Mar. Pollut. Bull. 2023, 192, 115017. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Zhao, L.; Ma, R.; Wang, J.; Du, L. FoxO signaling and mitochondria-related apoptosis pathways mediate tsinling lenok trout (Brachymystax lenok tsinlingensis) liver injury under high temperature stress. Int. J. Biol. Macromol. 2023, 251, 126404. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Cao, Y.; Jiang, Y.; Wei, Y.; Liu, H. Melatonin protects mouse granulosa cells against oxidative damage by inhibiting FOXO1-mediated autophagy: Implication of an antioxidation-independent mechanism. Redox. Biol. 2018, 18, 138–157. [Google Scholar] [CrossRef]
- Pal, S.; Haldar, C.; Verma, R. Impact of photoperiod on uterine redox/inflammatory and metabolic status of golden hamster, Mesocricetus auratus. J. Exp. Zool. A Ecol. Integr. Physiol. 2022, 337, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M. Role of common carp (Cyprinus carpio) in aquaculture production systems. Front. Life Sci. 2015, 8, 399–410. [Google Scholar] [CrossRef]
- Su, S.; Li, H.; Du, F.; Zhang, C.; Li, X.; Jing, X.; Liu, L.; Li, Z.; Yang, X.; Xu, P.; et al. Combined QTL and Genome Scan Analyses with the Help of 2b-RAD Identify Growth-Associated Genetic Markers in a New Fast-Growing Carp Strain. Front. Genet. 2018, 9, 592. [Google Scholar] [CrossRef]
- Zhang, C.; Su, S.; Li, X.; Li, B.; Yang, B.; Zhu, J.; Wang, W. Comparative transcriptomics identifies genes differentially expressed in the intestine of a new fast-growing strain of common carp with higher unsaturated fatty acid content in muscle. PLoS ONE 2018, 13, e0206615. [Google Scholar] [CrossRef]
- Bradshaw, W.E.; Holzapfel, C.M. Light, time, and the physiology of biotic response to rapid climate change in animals. Annu. Rev. Physiol. 2010, 72, 147–166. [Google Scholar] [CrossRef]
- Karatsoreos, I.N.; Bhagat, S.M.; Bloss, E.B.; Morrison, J.H.; McEwen, B.S. Disruption of circadian clocks has ramifications for metabolism, brain, and behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 1657–1662. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bowden, T.J.; Thompson, K.D.; Morgan, A.L.; Gratacap, R.M.; Nikoskelainen, S. Seasonal variation and the immune response: A fish perspective. Fish Shellfish Immunol. 2007, 22, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Nelson, R.; Demas, G.; Klein, S.; Kriegsfeld, L.; Bronson, F. Seasonal Patterns of Stress, Immune Function, and Disease; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Huber, M.; Bengtson, D.A. Effects of photoperiod and temperature on the regulation of the onset of maturation in the estuarine fish Menidia beryllina (Cope) (Atherinidae). J. Exp. Mar. Biol. Ecol. 1999, 240, 285–302. [Google Scholar] [CrossRef]
- Deng, X.; Dong, Q.; Bridges, D.; Raghow, R.; Park, E.A.; Elam, M.B. Docosahexaenoic acid inhibits proteolytic processing of sterol regulatory element-binding protein-1c (SREBP-1c) via activation of AMP-activated kinase. Biochim. Biophys. Acta 2015, 1851, 1521–1529. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Guo, L.; Deng, B.; Li, M.; Yang, T.; Yang, F.; Yang, Z. Long non-coding RNA HR1 participates in the expression of SREBP-1c through phosphorylation of the PDK1/AKT/FoxO1 pathway. Mol. Med. Rep. 2018, 18, 2850–2856. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wu, X.; Lai, J.; Liu, Y.; Song, M.; Li, F.; Gong, Q. Characterization of two lipid metabolism-associated genes and their expression profiles under different feeding conditions in Acipenser dabryanus. Aquac. Rep. 2021, 21, 100780. [Google Scholar] [CrossRef]
- Liu, K.; Liu, H.; Chi, S.; Dong, X.; Yang, Q.; Tan, B. Effects of different dietary lipid sources on growth performance, body composition and lipid metabolism-related enzymes and genes of juvenile golden pompano. Trachinotus ovatus. Aquac. Res. 2018, 49, 717–725. [Google Scholar] [CrossRef]
- Van der Oost, R.; Beyer, J.; Vermeulen, N.P. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Olsvik, P.A.; Kristensen, T.; Waagbø, R.; Rosseland, B.O.; Tollefsen, K.E.; Baeverfjord, G.; Berntssen, M.H. mRNA expression of antioxidant enzymes (SOD, CAT and GSH-Px) and lipid peroxidative stress in liver of Atlantic salmon (Salmo salar) exposed to hyperoxic water during smoltification. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2005, 141, 314–323. [Google Scholar] [CrossRef]
- Singhal, S.S.; Singh, S.P.; Singhal, P.; Horne, D.; Singhal, J.; Awasthi, S. Antioxidant role of glutathione S-transferases: 4-Hydroxynonenal, a key molecule in stress-mediated signaling. Toxicol. Appl. Pharmacol. 2015, 289, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xu, P.; Wen, H.; Xue, M.; Wang, Q.; He, J.; He, C.; Su, S.; Li, J.; Yu, F.; et al. Hypothermia-mediated oxidative stress induces immunosuppression, morphological impairment and cell fate disorder in the intestine of freshwater drum, Aplodinotus grunniens. Aquaculture 2023, 575, 739805. [Google Scholar] [CrossRef]
- Ma, H.; Wei, P.; Li, X.; Liu, S.; Tian, Y.; Zhang, Q.; Liu, Y. Effects of photoperiod on growth, digestive, metabolic and non-special immunity enzymes of Takifugu rubripes larvae. Aquaculture 2021, 542, 736840. [Google Scholar] [CrossRef]
- Kiffin, R.; Bandyopadhyay, U.; Cuervo, A.M. Oxidative stress and autophagy. Antioxid. Redox Signal. 2006, 8, 152–162. [Google Scholar] [CrossRef]
- Zhou, Z.; He, Y.; Wang, S.; Wang, Y.; Shan, P.; Li, P. Autophagy regulation in teleost fish: A double-edged sword. Aquaculture 2022, 558, 738369. [Google Scholar] [CrossRef]
- Jia, R.; Du, J.; Cao, L.; Feng, W.; He, Q.; Xu, P.; Yin, G. Immune, inflammatory, autophagic and DNA damage responses to long-term H2O2 exposure in different tissues of common carp (Cyprinus carpio). Sci. Total Environ. 2020, 757, 143831. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Shi, L.; Liu, H.; Wang, H. Hypoxia activates autophagy by Akt/FoxO1 pathway in fish cells. Aquac. Fish. 2023. [Google Scholar] [CrossRef]
- Mou, J.; Xu, J.; Wang, Z.; Wang, C.; Yang, X.; Wang, X.; Xue, H.; Wu, M.; Xu, L. Effects of photoperiod on morphology and function in testis and epididymis of Cricetulus barabensis. J. Cell. Physiol. 2021, 236, 2109–2125. [Google Scholar] [CrossRef]
- Gao, L.; Yuan, Z.; Zhou, T.; Yang, Y.; Gao, D.; Dunham, R.; Liu, Z. FOXO genes in channel catfish and their response after bacterial infection. Dev. Comp. Immunol. 2019, 97, 38–44. [Google Scholar] [CrossRef]
- Duan, P.; Tian, Y.; Li, Z.; Chen, S.; Li, L.; Wang, X.; Wang, L.; Liu, Y.; Zhai, J.; Li, W.; et al. Comparative transcriptome analysis of hybrid Jinhu grouper (Epinephelus fuscoguttatus ♀ × Epinephelus tukula ♂) and Epinephelus fuscoguttatus under temperature stress. Aquaculture 2024, 578, 740037. [Google Scholar] [CrossRef]
- Cheng, Z.; Guo, S.; Copps, K.; Dong, X.; Kollipara, R.; Rodgers, J.T.; Depinho, R.A.; Puigserver, P.; White, M.F. Foxo1 integrates insulin signaling with mitochondrial function in the liver. Nat. Med. 2009, 15, 1307–1311. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; White, M.F. Targeting Forkhead box O1 from the concept to metabolic diseases: Lessons from mouse models. Antioxid. Redox Signal. 2011, 14, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, Y.I.; Kitamura, T.; Kruse, J.P.; Raum, J.C.; Stein, R.; Gu, W.; Accili, D. FoxO1 protects against pancreatic beta cell failure through NeuroD and MafA induction. Cell Metab. 2005, 2, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z. The FoxO-Autophagy Axis in Health and Disease. Trends Endocrinol. Metab. 2019, 30, 658–671. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Wang, X.; Qin, W.; Jiang, J.; Cheng, L. Emerging regulatory mechanisms and functions of autophagy in fish. Aquaculture 2019, 11, 734212. [Google Scholar] [CrossRef]
- Xiong, X.; Tao, R.; DePinho, R.A.; Dong, X.C. The autophagy-related gene 14 (Atg14) is regulated by forkhead box O transcription factors and circadian rhythms and plays a critical role in hepatic autophagy and lipid metabolism. J. Biol. Chem. 2012, 287, 39107–39114. [Google Scholar] [CrossRef]
- Ma, D.; Panda, S.; Lin, J.D. Temporal orchestration of circadian autophagy rhythm by C/EBPβ. EMBO J. 2011, 30, 4642–4651. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′−3′) | Fragment Length (bp) |
|---|---|---|
| SREBP1C | F: GCCTGCTTCACTTCACTACT | 137 |
| R: CCAGTCCTCATCCACAAA | ||
| FAS | F: GTGTACGCCACCGCCTATTA | 104 |
| R: ATAGCAATAGCGGCCTGTCC | ||
| ACCα | F: AGACCGTATCTACAGGCACT | 107 |
| R: GCATCTTATGGTTGGCAC | ||
| CPT1 | F: TGACCTACAGTTGAGCCG | 196 |
| R: AATCATGCCCATAGAGGG | ||
| LPL | F: GACAATGGCACAGAATGG | 190 |
| R: ACATACCCGTAACCGTCC | ||
| ATG9 | F: ACAAGCGTGGAGGGAACCGT | 184 |
| R: AGGGACCAACATCGAGCA | ||
| BNIP3 | F: ACGGGAATCCAGCAGTAG | 162 |
| R: ATCCTTGCGACAGCCTCAG | ||
| BCL2 | F: AGCGGCTTTATCAGTCGG | 162 |
| R: CACAAACGGTCCCTCCAA | ||
| BCL2L1 | F: CGCATCGTGGGACTGTTT | 241 |
| R: TCATTCCAGCCAGCAACC | ||
| CYCLIND | F: GCGGCTACACTGAACTCT | 210 |
| R: GCTGGCTCTTTCCTCTTCAA | ||
| CYCLING2 | F: TCTTCGGTTACAGCACTCAG | 142 |
| R: TCTTCTTTGGTCACTCGG | ||
| MTMR3 | F: GGAGGGCACTAAATGGTT | 178 |
| R: GATGGTCCTGTAGAAGGGA | ||
| FOXO1 | F: ACGAACTTGGCGACTCTG | 183 |
| R: CCACTGATGGGTTCTTAGG | ||
| FOXO3 | F: ACGCCTGGGGAAACTATT | 200 |
| R: GACAAAGCGACTGTGGAG | ||
| FOXO4 | F: CCACCGAGGAAGATAAACAC | 161 |
| R: GATCAGGTCTGCGTAGGA | ||
| GAPDH | F: CCGTTCATGCTATCACAGCTACACA | 310 |
| R: GTGGATACCACCTGGTCCTCTG |
| Con | SP | |
|---|---|---|
| Final body weight (g) | 55.44 ± 2.09 | 62.39 ± 2.75 * |
| Body length (cm) | 13.08 ± 0.21 | 13.55 ± 0.20 |
| Body width (mm) | 42.24 ± 0.60 | 42.97 ± 0.74 |
| Body thickness (mm) | 22.84 ± 0.35 | 23.77 ± 0.45 |
| Hepatosomatic index (HSI) (%) | 1.53 ± 0.04 | 1.68 ± 0.05 * |
| Condition factor (CF) (g/cm3) | 0.026 ± 0.00 | 0.025 ± 0.00 |
| Con-1 | Con-2 | Con-3 | SP-1 | SP-2 | SP-3 | |
|---|---|---|---|---|---|---|
| Raw reads | 46,576,536 | 41,233,316 | 45,346,460 | 44,230,214 | 45,064,924 | 43,351,336 |
| Clean reads | 44,365,184 | 39,283,252 | 43,104,336 | 43,551,724 | 44,496,136 | 42,217,336 |
| Q20 (%) | 97.13 | 97.07 | 97.45 | 97.94 | 97.93 | 97.56 |
| Q30 (%) | 92.21 | 92.14 | 93.01 | 93.93 | 94.04 | 93.07 |
| GC content (%) | 48.19 | 47.61 | 47.33 | 48.09 | 47.48 | 47.75 |
| Total mapped | 37,300,129 | 32,077,643 | 36,404,503 | 38,222,064 | 38,312,667 | 37,036,832 |
| Mapping rate (%) | 84.08 | 81.66 | 84.46 | 87.76 | 86.1 | 87.73 |
| Uniquely mapped | 34,006,887 | 29,918,503 | 34,285,329 | 34,530,992 | 35,449,121 | 33,857,504 |
| Uniquely mapped rate (%) | 76.65 | 76.16 | 79.54 | 79.29 | 79.67 | 80.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Su, S.; Dong, P.; Feng, W.; Li, J.; Zhang, C.; Tang, Y. Effects of Seasonal Photoperiod on Growth, Lipid Metabolism, and Antioxidant Response in the Huanghe Carp (Cyprinus carpio haematopterus). Fishes 2023, 8, 595. https://doi.org/10.3390/fishes8120595
Wang W, Su S, Dong P, Feng W, Li J, Zhang C, Tang Y. Effects of Seasonal Photoperiod on Growth, Lipid Metabolism, and Antioxidant Response in the Huanghe Carp (Cyprinus carpio haematopterus). Fishes. 2023; 8(12):595. https://doi.org/10.3390/fishes8120595
Chicago/Turabian StyleWang, Wenqian, Shengyan Su, Ping Dong, Wenrong Feng, Jianlin Li, Chengfeng Zhang, and Yongkai Tang. 2023. "Effects of Seasonal Photoperiod on Growth, Lipid Metabolism, and Antioxidant Response in the Huanghe Carp (Cyprinus carpio haematopterus)" Fishes 8, no. 12: 595. https://doi.org/10.3390/fishes8120595
APA StyleWang, W., Su, S., Dong, P., Feng, W., Li, J., Zhang, C., & Tang, Y. (2023). Effects of Seasonal Photoperiod on Growth, Lipid Metabolism, and Antioxidant Response in the Huanghe Carp (Cyprinus carpio haematopterus). Fishes, 8(12), 595. https://doi.org/10.3390/fishes8120595