1. Introduction
Enteromorpha prolifera is a type of green marine macroalgae that can cause green tides due to eutrophication [
1]. Considering the potential of a detrimental effect on the environment, this algal blooms in coastal areas of Qingdao of China in 2008 caused a great concern for the local government and environmentalists [
2]. After that, specialized commercial companies from Qingdao were established to clean up
E. prolifera. An important commercial use of
E. prolifera collected from the sea was to become feed ingredients [
3]. Traditionally, macroalgae including
E. prolifera were used as a seaweed meal in compound animal feed by drying and grinding to a fine powder [
4]. However, aquafeeds demand for
E. prolifera meal has gradually exceeded annual
E. prolifera production. Thus, except for the drying technique, other effective processing technologies should be used to obtain extracted bioactive compounds of
E. prolifera for high-value applications in aquaculture [
4].
E. prolifera contains sulfated polysaccharides mainly composed of rhamnose, glucuronic acid and xylose, which are regarded as the biologically active molecules [
5,
6]. Because of these,
E. prolifera polysaccharide has been used as the feed additive to regulate various biological activities, such as immunomodulatory, antioxidant, antimicrobial, and lipid metabolism [
7,
8]. These biological activities of
E. prolifera polysaccharide were not only confirmed in terrestrial animals [
5,
9], but also in fish and shrimp [
2,
10]. In addition, some studies found that seaweed polysaccharide could mitigate various stress responses, such as air exposure, heat stress and ammonia stress [
11,
12,
13,
14]. Except for the polysaccharide,
E. prolifera had been shown to enhance feed intake of fish due to its potential feeding attractants, such as betaine [
2,
15]. However, aforementioned studies with
E. prolifera meal or
E. prolifera polysaccharide were mainly based on the traditional drying process or extraction procedures on a laboratory scale. Novel extraction methods should be applied to increase the bioactive compounds of
E. prolifera at the level of large-scale industrialization to support the efficient utilization of
E. prolifera in aquafeeds.
Pacific white shrimp (
Litopenaeus vannamei) is an important cultured shrimp species, accounting for 70% of the total shrimp production in the world [
16]. As a feed ingredient,
E. prolifera meal was mainly used in shrimp feed.
E. prolifera inclusion level in aquaculture feed has to be decreased because its production is finite, but the aquaculture industry has still been expanding so far. Thus, the aim of this study was to evaluate three types of bio-products of
E. prolifera (
E. prolifera meal,
E. prolifera hydrolysate and
E. prolifera polysaccharide), which is supplied on a large scale by a commercial company. According to biological activities of
E. prolifera, the growth, feed intake, lipid metabolism, antioxidant, immune parameters and stress resistance were investigated in Pacific white shrimp.
4. Discussion
E. prolifera is a green algal species, which contains various feeding attractants such as dimethyl-
β-propiothetin (DMPT), dimethylsulfoniopropionate (DMSP), and betaine [
15,
23,
24,
25]. Asino [
2] reported that
E. prolifera showed a positive effect on the growth performance of the larger yellow croaker (
Pseudosciaena crocea), which was attributed, to some extent, to increasing feed intake. Thus, in the present study, the cumulative feed intake per shrimp for 14 days after the beginning of the feeding trial and after feed intake of the entire feeding period were determined to evaluate the attractant effects of
E. prolifera bio-products. Results of the cumulative feed intake per shrimp showed that statistical difference was observed 7 days after feeding. For aquatic animals, there were two main sensorial stimuli (olfaction and gustation) to distinguish the same type of feed [
23]. Thus, it could be speculated that bio-products of
E. prolifera mainly affected the gustation rather than the olfaction, thereby improving the feeding response of Pacific white shrimp [
26]. In addition, compared with the control diet, only the EPH0.4%, EPM3%, and DMPT0.1% groups showed significantly higher feed intake than the control group. It indicated that 3% of
E. prolifera meal and 0.4% of
E. prolifera hydrolysate in diets could achieve similar attractant effects as 0.1% of DMPT on Pacific white shrimp. Meanwhile, there was no significant difference in feed intake between the EPP0.03% and control groups. Considering that shrimp fed the EPP0.03% diet presented a numerically higher feed intake than the control group, it did not mean that 0.03% of
E. prolifera polysaccharide was not an attractant effect, but might be related to its volume in feed. The results of
E. prolifera hydrolysate supported this hypothesis, where feeding intake of the control group was not significantly different from the 0.2% of
E. prolifera hydrolysate group, but was numerically lower, and showed a significant decrease compared to the 0.4% of
E. prolifera hydrolysate group. Taken together, consistent with the larger yellow croaker, the results of growth including final body weight, weight gain, and specific growth rate were the highest in the 0.4% of
E. prolifera hydrolysate group, which further verified that bio-products of
E. prolifera promoted growth of shrimp partly due to the increased feeding intake [
2].
In rats, sulfated polysaccharides from
E. prolifera exhibited the hypolipidaemic activity by regulating the lipid metabolism [
27]. Oral administration of
E. prolifera polysaccharide at a dosage of 100, 200 and 300 mg/kg body weight successfully reduced plasma total cholesterol, TG and LDL-C in a dose-dependent manner in mice fed high-fat diet [
5]. Subsequent studies revealed the underlying mechanism that
E. prolifera reduced cholesterol content by inhibiting sterol regulatory element binding protein-2 (SREBP-2) and 3-hydroxy-3-methylglutaryl-CoA reductase [
28], while reducing TG content by regulating SREBP-1c and acetyl-CoA carboxylase [
29]. Similar to results in mice, the present study showed that either
E. prolifera hydrolysate,
E. prolifera polysaccharide or
E. prolifera meal decreased TG and LDL-C in plasma. It indicated that bio-products of
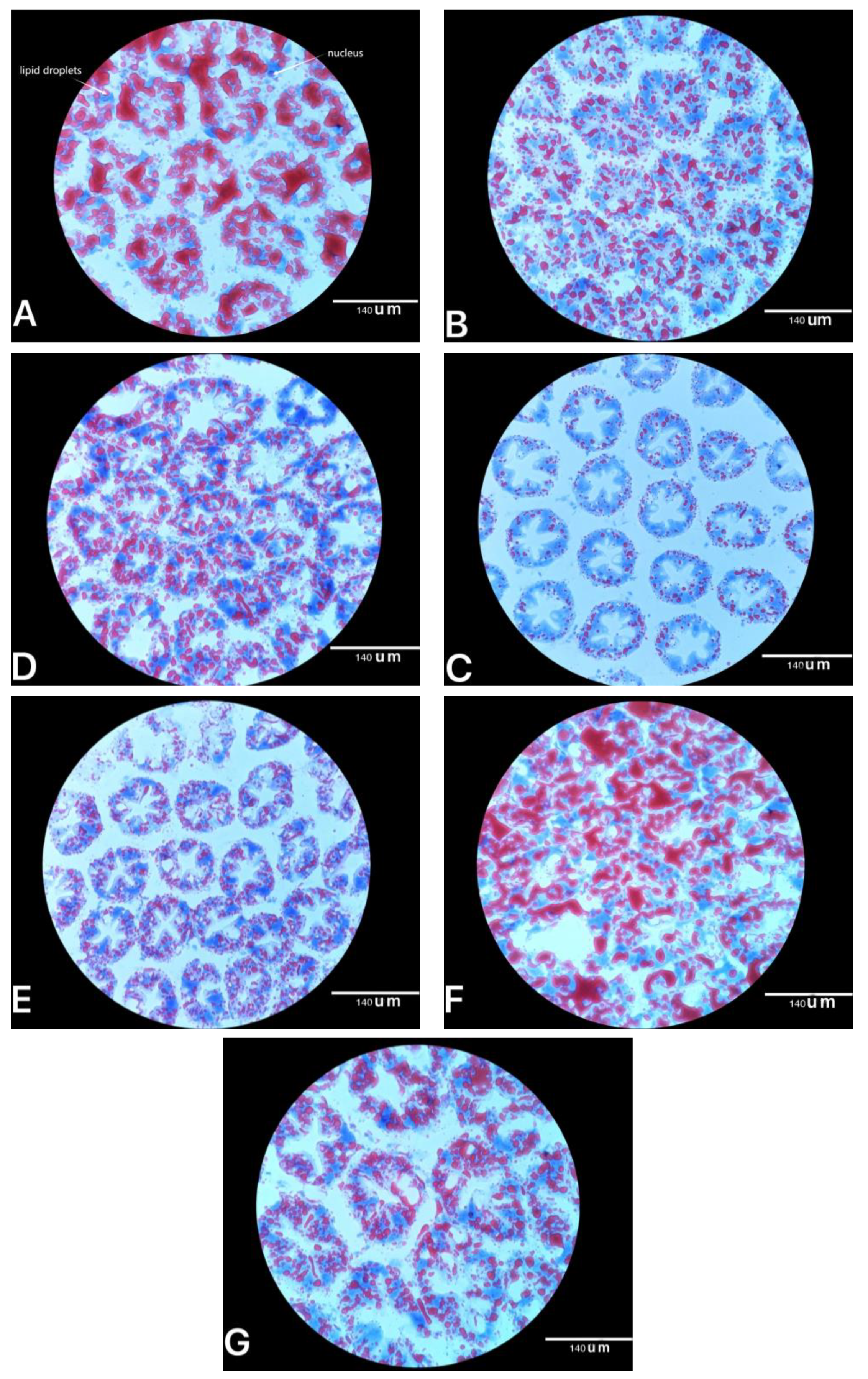
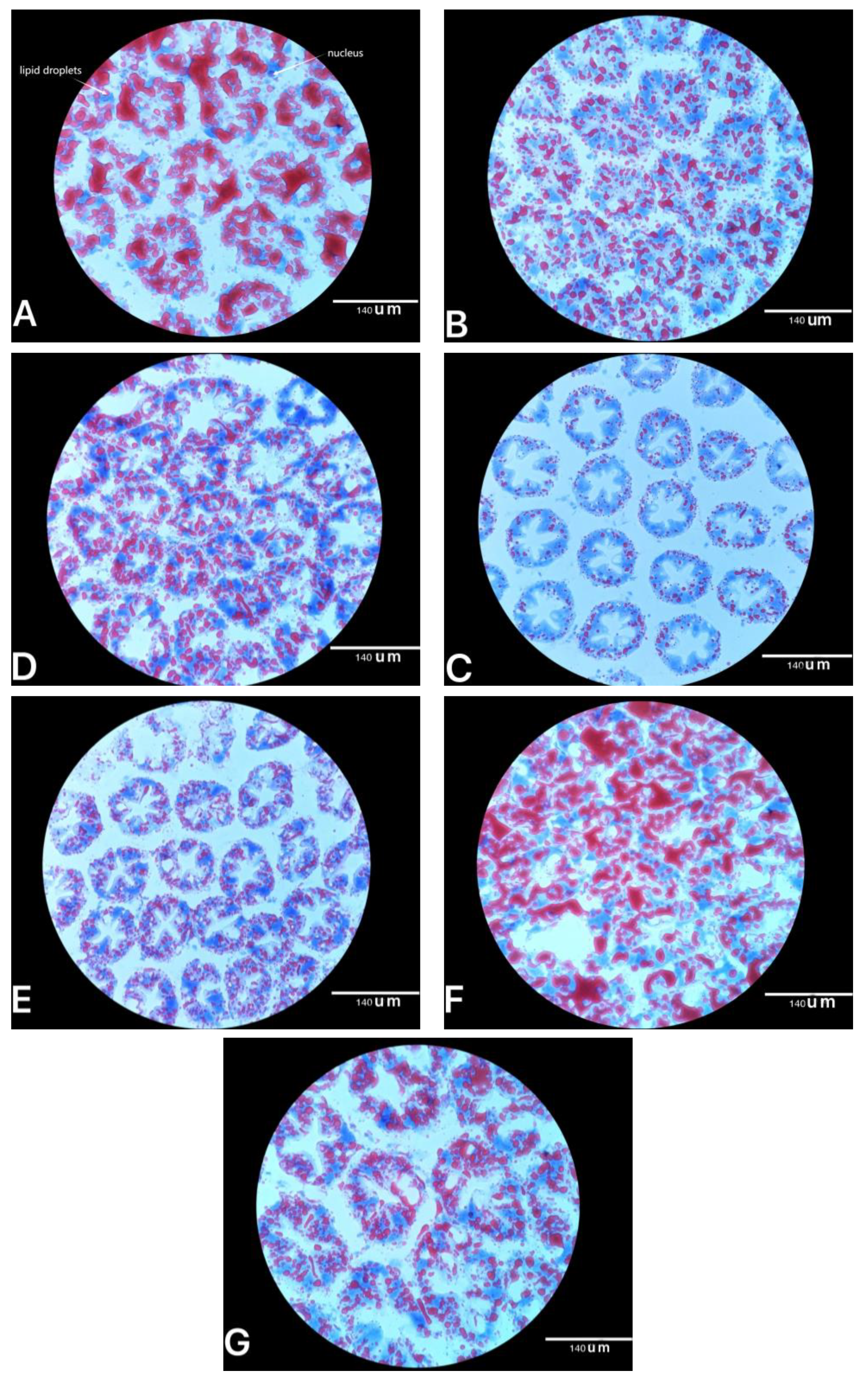
E. prolifera obtained from the three types of the drying process or extraction procedure had similar hypolipidaemic effects on Pacific white shrimp as those in mice. In addition, oil red O staining of the hepatopancreas showed that the abundance of lipid droplets in shrimp fed the bio-products of
E. prolifera, especially 0.4% of
E. prolifera hydrolysate and 3% of
E. prolifera meal, significantly decreased compared with the control group. Those results were supported by the hepatopancreas TG concentrations, which indicated that three types of
E. prolifera bio-products could effectively reduce the accumulation of lipids in hepatopancreas [
27].
It had been reported that
E. prolifera had antioxidant, immunomodulatory and anti-inflammatory properties in many aquatic animals, such as banana shrimp (
Fenneropenaeus merguiensis), red tilapia (
Oreochromis mossambicus ×
Oreochromis niloticus) and crucian carp (
Carassius auratus) [
6,
10,
30]. Reactive oxygen species (ROS) are by-products of aerobic metabolism, which can cause oxidative damage by increasing their intracellular levels [
31]. To avoid the damaging effects of naturally producing ROS, the living organisms including aquatic animals possess two distinct systems of antioxidant defense, the enzymatic antioxidant system such as SOD and GPx as well as the non-enzyme antioxidants such as GSH [
32,
33]. Many studies had demonstrated the involvement of antioxidant systems in repairing or preventing oxidative stress [
33]. In our study, three types of
E. prolifera bio-products increased the content of plasma GSH or the activity of plasma SOD, especially 0.03% of
E. prolifera polysaccharide for GSH or 0.4% of
E. prolifera hydrolysate and 3% of
E. prolifera meal for SOD. It indicated that bio-products of
E. prolifera based on different processing procedures could regulate some antioxidant enzymes and non-enzymatic antioxidant substances in Pacific white shrimp to some extent, which was able to eliminate the excess of superoxide or free radicals, thus resulting in improving the antioxidant capacity. The significantly lower level of plasma MDA (a product of lipid peroxidation) was observed in fish fed the EPM3% diet compared with the control diet, which supported the conclusion that
E. prolifera bio-products could improve antioxidant defense. As regards to the non-specific immunity of shrimp, most studies have evaluated the immunity by measuring the activities of immune-related enzymes in plasma, such as ACP, AKP, GPT GOP, LZM and PO [
34,
35,
36]. In the present study, however, the changes of most immune-related enzyme activities in the plasma of shrimp fed
E. prolifera bio-products were not found to improve the immunity of shrimp compared with the control group. Only the activity of PO in shrimp fed 0.2% of
E. prolifera hydrolysate was significantly higher than that of the control group. But, given that it had been well established that polysaccharides from
E. prolifera could enhance immunity [
7,
10], the results of our study could not negate this conclusion solely on the basis of the results of immune-related enzyme activities in plasma. Therefore, in future studies, a pathogen challenge test needs to be conducted to confirm whether polysaccharides from
E. prolifera could improve the immunity of Pacific white shrimp. In addition, in this study, the inflammatory status of shrimp was evaluated by analyzing the expression of a tumor necrosis factor-
α (
tnf-
α) in the hepatopancreas.
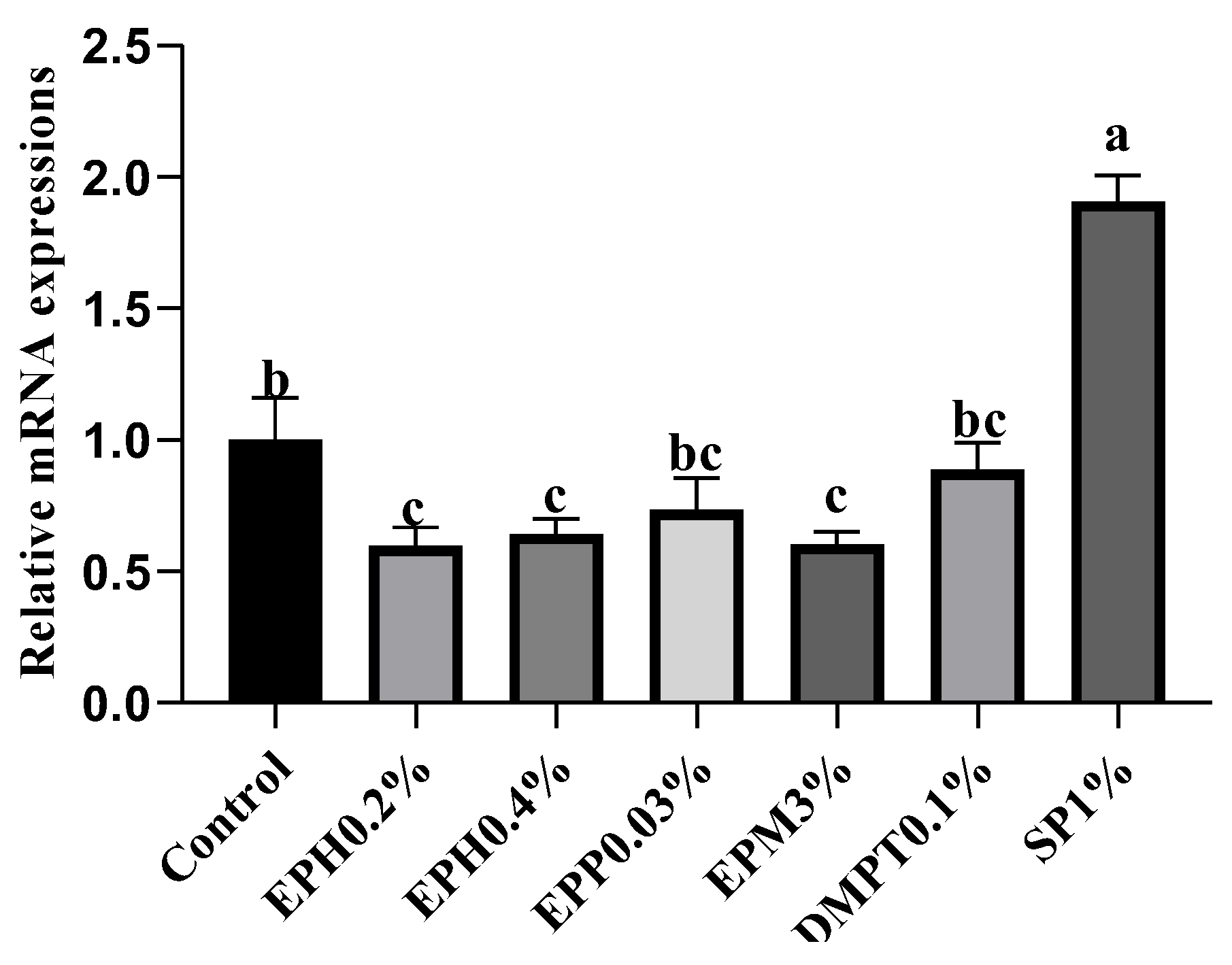
tnf-
α is a typical pro-inflammatory cytokine produced by immune cells [
37], and previous studies have found that polysaccharides from
Atractylodis macrocephalae (Chinese herbal medicine) could decrease its expression in the hepatopancreas of Pacific white shrimp [
38]. In broilers, heat stress caused a significant up-regulation of
tnf-α expression, indicating that it induced inflammatory responses. However, polysaccharides from
E. prolifera effectively decreased the level of
tnf-
α expression, which demonstrated that it might play an important role in suppressing inflammatory responses [
11]. In our study, although it could not be confirmed whether inflammation occurred in the control group during the feeding trial, combined with the above studies, it partially indicated that the down-regulation of
tnf-α expression by
E. prolifera bio-products (EPH0.2%, EPH0.4% and EPM3%) could reduce the risk of inflammation in shrimp.
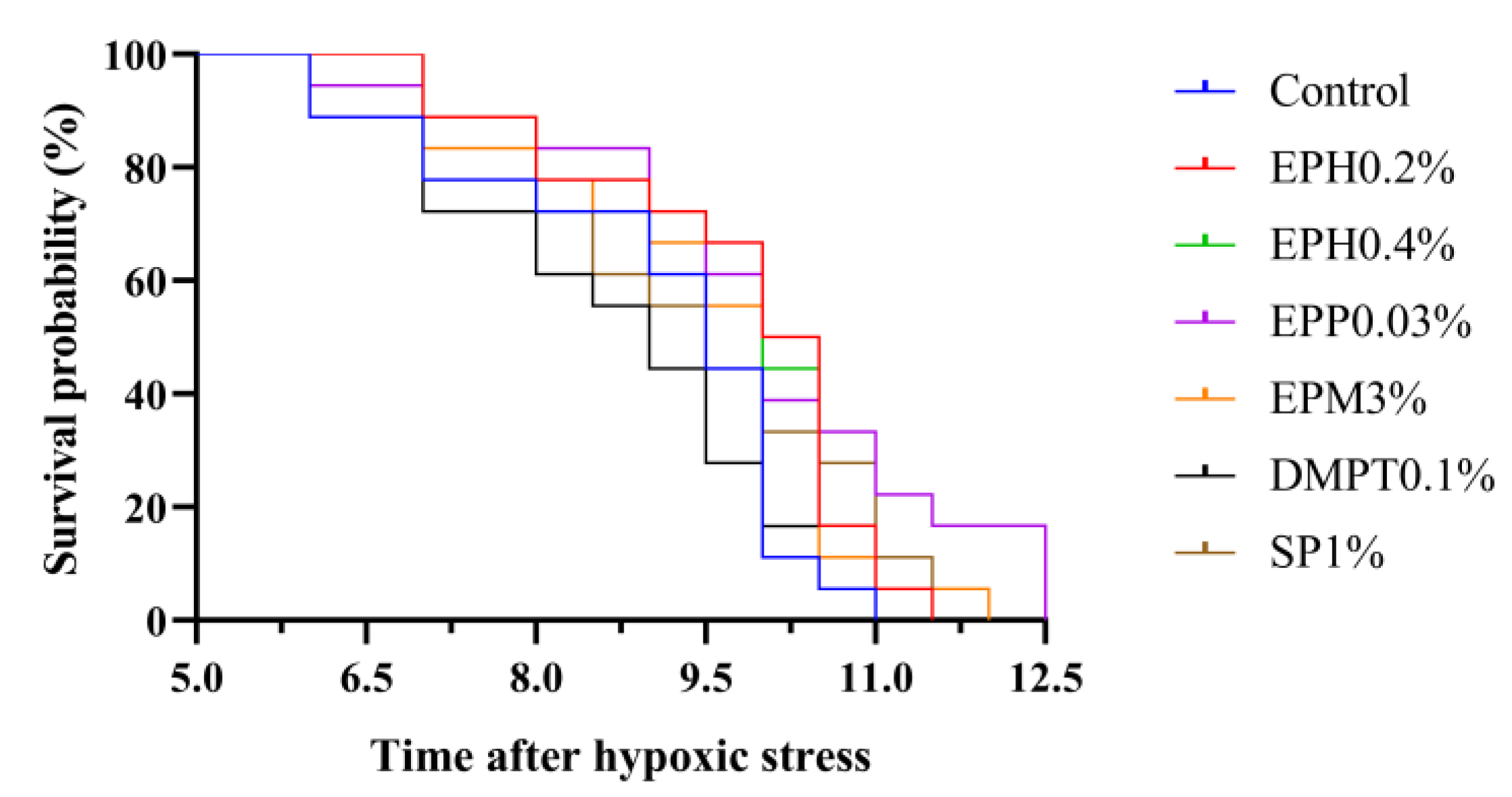
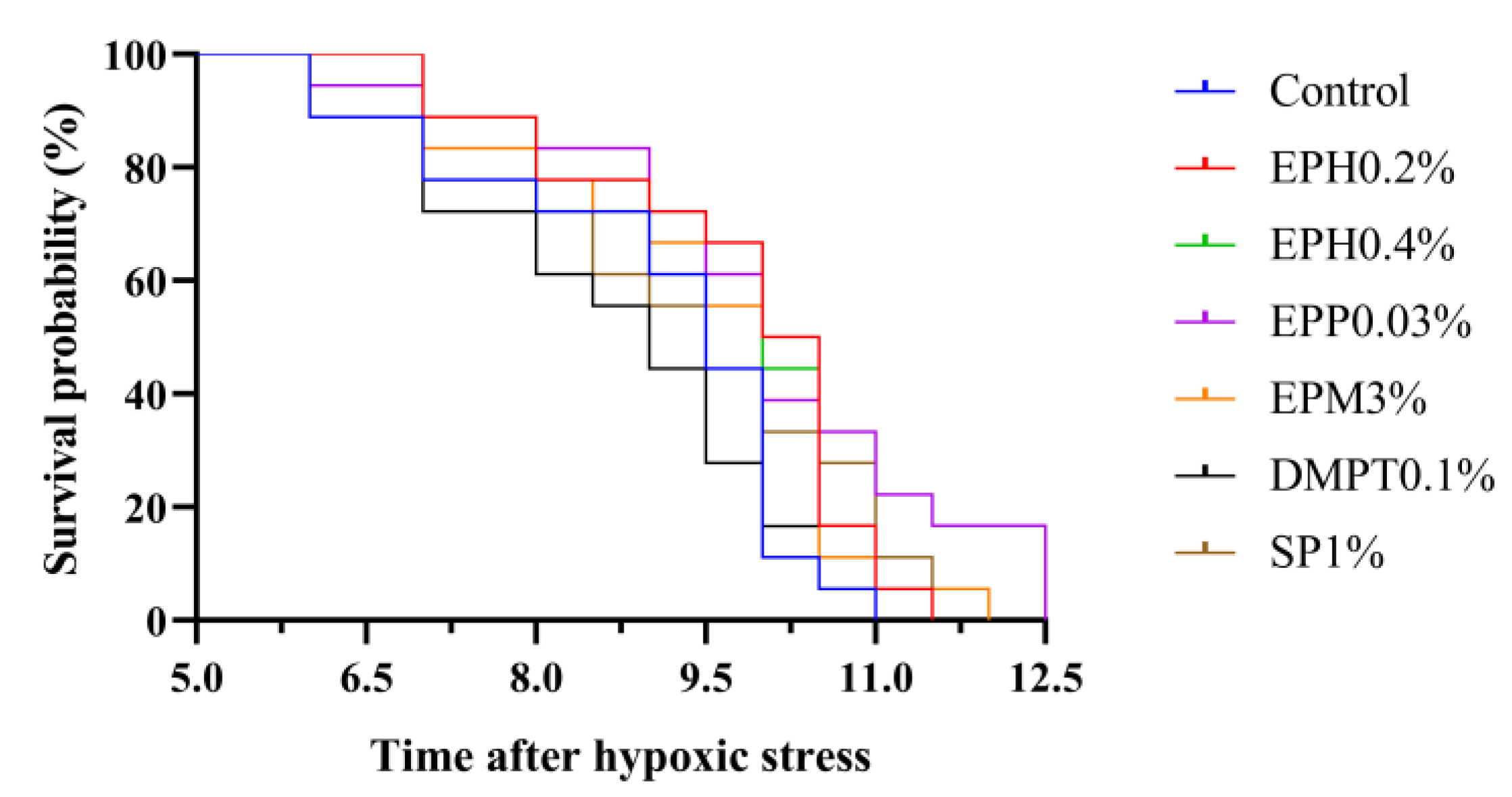
Apart from the lipid metabolism, antioxidant, and immune effects, the stress resistance of
E. prolifera bio-products was evaluated in this study. Polysaccharides from seaweed could improve survival rate of aquatic animals under various stress, such as air exposure, ammonia stress and heat stress [
12,
13,
14]. Survival rate was used as an important index for comprehensive evaluation of resistance to stress because its challenge tests reflected non-specific immune and anti-oxidation functions of the organisms [
39]. In the present study, based on Kaplan–Meier survival curve, survival probability in shrimp fed
E. prolifera bio-products (EPH0.2%: 89%; EPH0.4%: 89%; EPP0.03%: 89%; EPM3%: 83%) in 0.6 mg/L at 7 h after hypoxic stress was higher than that of the control diet (78%), and this trend persisted until the dissolved oxygen reached 0.2 mg/L at 10.5 h after hypoxic stress. It indicated that
E. prolifera bio-products could improve the resistance of shrimp to hypoxic stress.




 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}