
Proposal of Brotulella n. gen. for Monogeneans from the Gills of the Pacific Bearded Brotula Brotula clarkae Hubbs, 1944 (Ophidiiformes: Ophidiidae) Based on Morphological and Molecular Evidence

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimen Collection and Morphological Analyses
2.2. Molecular Characterization and Phylogenetic Analyses
3. Results
3.1. Order Dactylogyridea Bychowsky, 1937
- Dactylogyridae Bychowsky, 1933
- Brotulella n. gen. Cruces, Chero & Luque
- Diagnosis
- Body divided into cephalic region, trunk, peduncle, and haptor. Tegument thin, smooth. Head organs, cephalic lobes present; cephalic glands unicellular, anterior and posterolateral to pharynx. Eyespots absent; accessory chromatic granules absent. Mouth subterminal, midventral; pharynx muscular, glandular; oesophagus short, intestine bifurcated; intestinal caeca confluent posteriorly to testis, without diverticula. Common genital pore midventral, near level of intestinal bifurcation. Haptor armed with 14 hooks with ancyrocephaline distribution sensu Mizelle [15], 2 pairs of anchors, 2 haptoral bars, lacking haptoral reservoirs; anchors dissimilar, distal end of superficial and deep roots with stocking-shaped sheath; ventral bar bowed; dorsal bar with anterior and posterior broad process; hooks with undilated shanks and upright blunt thumb. Gonads tandem, intercaecal. Testis posterior to ovary. Vas deferens looping left intestinal caecum. Seminal vesicle almost sigmoid. Prostatic reservoirs two, with thick muscular walls. Male copulatory organ (MCO) tubular, sclerotized, with twisted distal end; base cylindrical; accessory piece delicate, membranous, articulated to shaft of MCO. Ovary U-shaped; oviduct, uterus not observed. Vagina sclerotized, emptying to seminal receptacle, vaginal aperture dextrolateral. Vitelline follicles dense, coextensive with intestinal caeca. Parasites of gills of marine fishes.
- Type species by original designation: Brotulella laurafernandae n. sp. from the gills of the Pacific bearded brotula Brotula clarkae Hubbs, 1944 (Ophidiiformes: Ophidiidae). Other species: Brotulella luisahelenae n. sp. from B. clarkae.
- Etymology: The genus name refers to the genus name of the fish host (Brotula). The diminutive -ella is appended to the genus name and should be treated as female.
- Remarks: Brotulella n. g. is characterized by the combination of the following features: (1) anchors with a stocking-shaped sheath associated with the distal end of superficial and deep roots, (2) tandem gonads, (3) a distally twisted MCO with a delicate membranous accessory piece articulated to the shaft of the MCO, (4) a U-shaped ovary, (5) an almost sigmoid seminal vesicle, and (6) two prostatic reservoirs with thick muscular walls. Brotulella n. g. most closely resembles species of the genus Platycephalotrema Kritsky & Nitta, 2019, which comprises nine species that infect the gill lamellae of the flathead fishes (Platycephalidae). Species of both genera share (1) tandem gonads, (2) a vas deferens looping left intestinal caecum, (3) two prostatic reservoirs, (4) a sclerotized vagina, (5) dextral vaginal aperture, (6) seven pairs of hooks, (7) and tubular MCO with complex distal end. However, Brotulella n. g. differs from Platycephalotrema by its species having anchors unequal (anchors equal in Platycephalotrema spp.); a stocking-shaped sheath associated with the distal end of superficial and deep roots of the anchors (absent in Platycephalotrema spp.); a MCO with accessory piece (absent in Platycephalotrema spp.); hooks with upright blunt thumb (protruding blunt thumb in Platycephalotrema spp.); a U-shaped ovary (ovary entire in Platycephalotrema spp.); a short vaginal vestibule (large vaginal vestibule in Platycephalotrema spp.); prostatic reservoirs with thick muscular walls (prostatic reservoirs without thick muscular walls in Platycephalotrema spp.); and absence of eyespots (four eyespots in Platycephalotrema spp.).
- Species of Brotulella n. g. resembles species of Ligophorus Euzet & Suriano, 1977, which includes species parasitizing mullets (Mugilidae). Members of both genera share the following features: (1) gonads in tandem, (2) a tubular sclerotized and uncoiled MCO, (3) an ovary with U-shaped, and (4) a sclerotized vagina. However, species of these genera differ by having anchors with a stocking-shaped sheath associated with the distal end of superficial and deep roots (absent in Ligophorus spp.), a vas deferens looping the left intestinal caecum (vas deferens not looping the intestinal caecum in Ligophorus spp.), two prostatic reservoirs with thick muscular walls (one pyriform prostatic reservoir without thick muscular wall in Ligophorus spp.) and by having a ventral bar bowed (V-shaped ventral bar in Ligophorus spp.).
- Species of the new genus slightly resemble those of Bravohollisia Bychowsky & Nagibina, 1970 and Caballeria Bychowsky & Nagibina, 1970 by having tandem gonads, a tubular sclerotized and uncoiled MCO, and a bowed ventral bar. However, species of Brotulella n. g. differ from species of these genera by having a MCO with accessory piece (absent in Bravohollisia and Caballeria species) and by the absence of haptoral glands (present in Bravohollisia and Caballeria species). Species of the new genus slightly resembles species of Mexicana Caballero & Bravo-Hollis, 1959, parasitic on haemulid fishes, by having tandem gonads and an almost U-shaped ovary but differ from these species by having two prostatic reservoirs (one prostatic reservoir in Mexicana spp.), a testis not bipartite (principally bipartite posteriorly in Mexicana spp.) and absence of eyespots (four eyespots in Mexicana spp.).
- Finally, species of Brotulella n. gen. resemble species of the Boegeriella Mendoza-Palmero & Hsiao, 2020 and Nanayella Acosta, Mendoza-Palmero, da Silva & Scholz, 2019 by the shape of the ovary (U-shaped). However, species of Brotulella n. gen. differ from Boegeriella spp. by the morphology of the MCO (tubular with twisted distal end in Brotulella n. gen. vs coiled in Boegeriella spp.). Species of the new genus can be distinguished from species of Nanayella by having hooks of similar size (hook of dissimilar size in Nanayella).
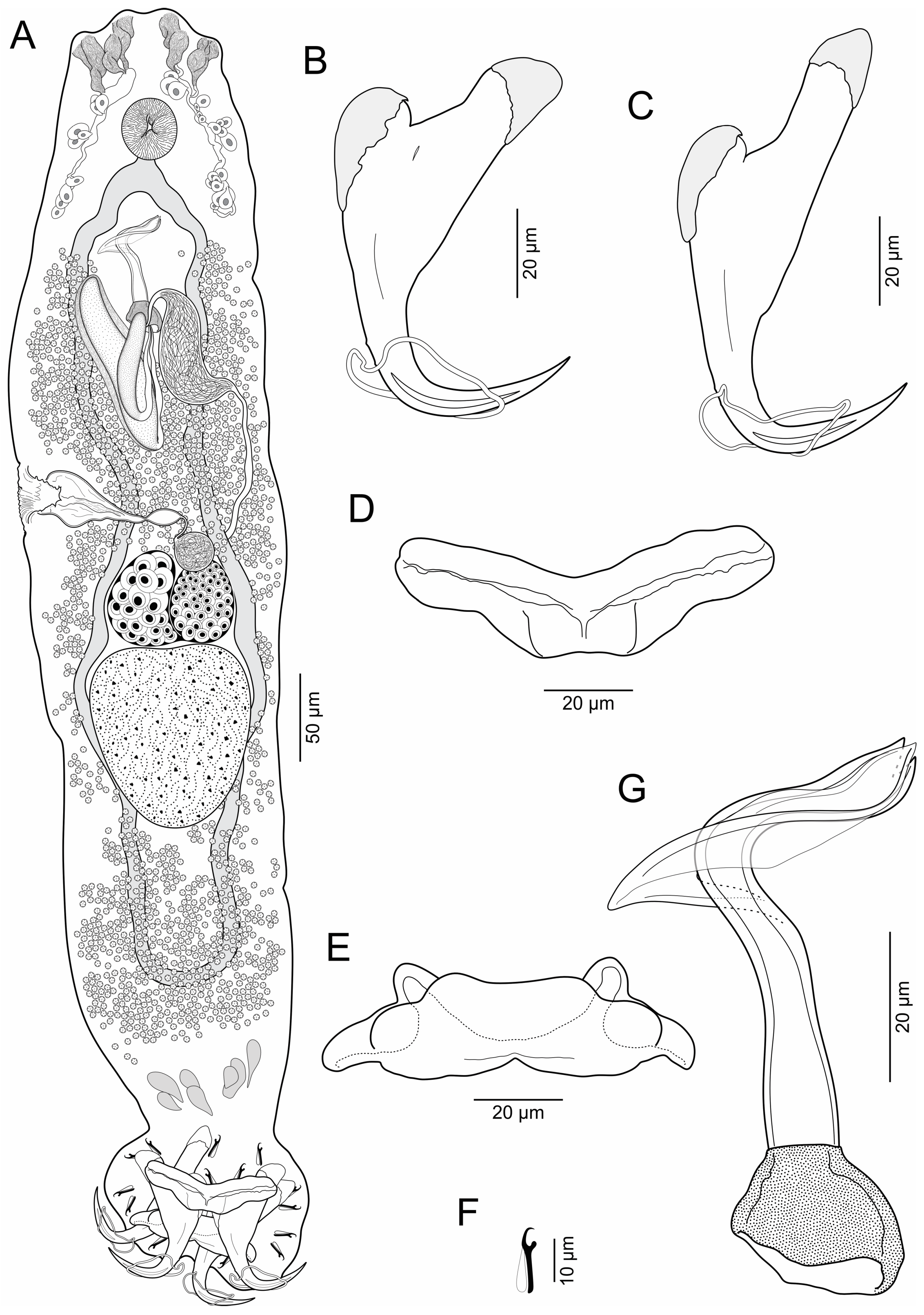
3.1.1. Brotulella laurafernandae n. sp. Cruces, Chero & Luque
- Type host: Brotula clarkae Hubbs, 1944 (Ophidiiformes: Ophidiidae), Pacific bearded brotula.
- Type locality: Puerto Pizarro resort (3°29′ S, 80°24′ W), Tumbes, Peru.
- Site in host: Gills.
- Type specimens: Holotype (MUSM-HEL 5132), 12 paratypes (MUSM-HEL 5133a–l), 1 hologenophore (MUSM 5133m).
- ZooBank registration: The Life Science Identifier (LSID) for Brotulella laurafernandae n. sp. is urn:lsid:zoobank.org:act:A1C5B22A-E8FF-4A70-B574-1B9A8B8082CC.
- Representative DNA sequence: Sequence was deposited in GenBank under the accession number OR860318 for the 28S rDNA with 720 bp.
- Etymology: The new species is named in honor of Laura Fernanda do Amarante Luque, daughter of the senior author.
- Remarks: Brotulella laurafernandae n. sp. is the type species of the genus.
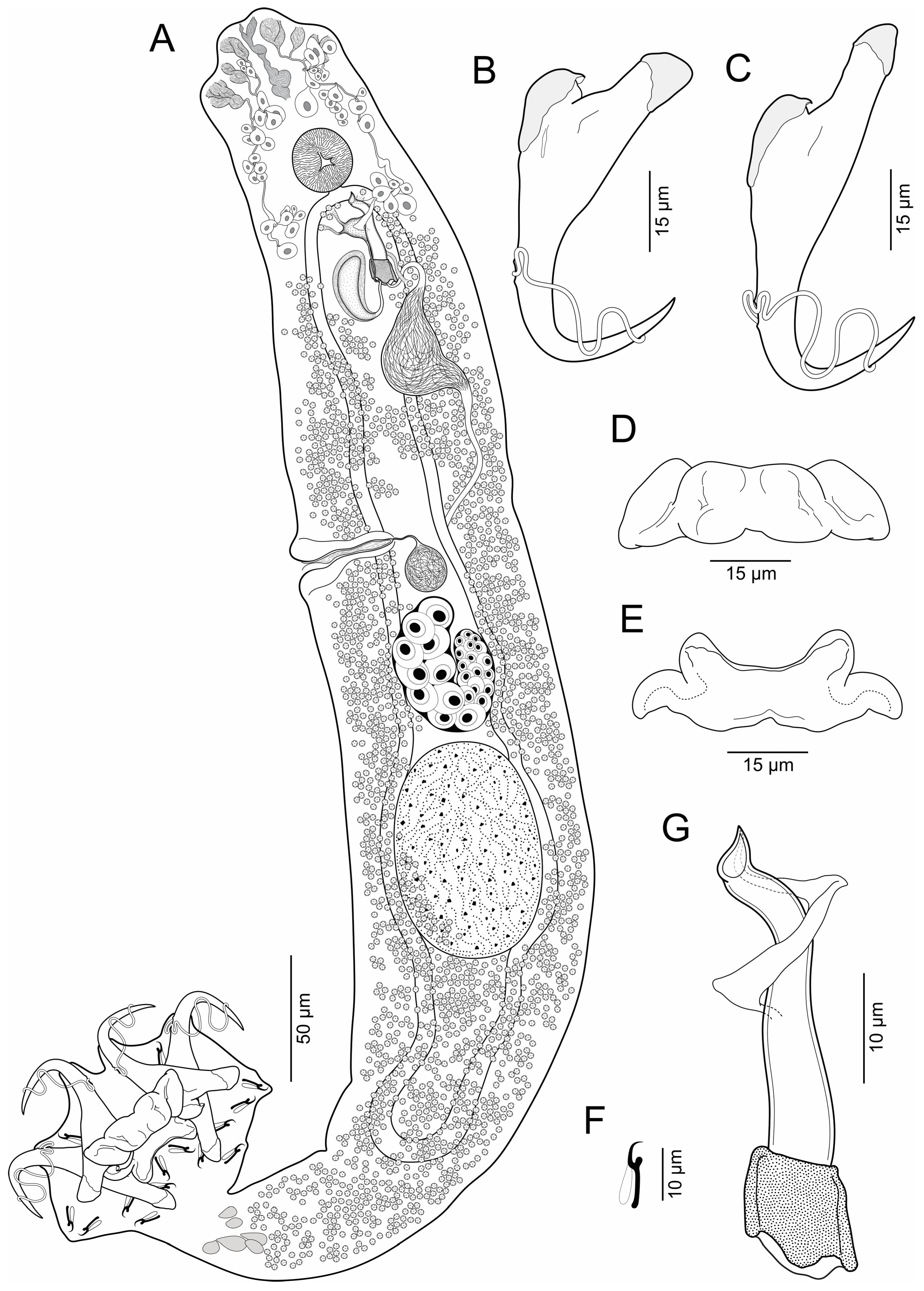
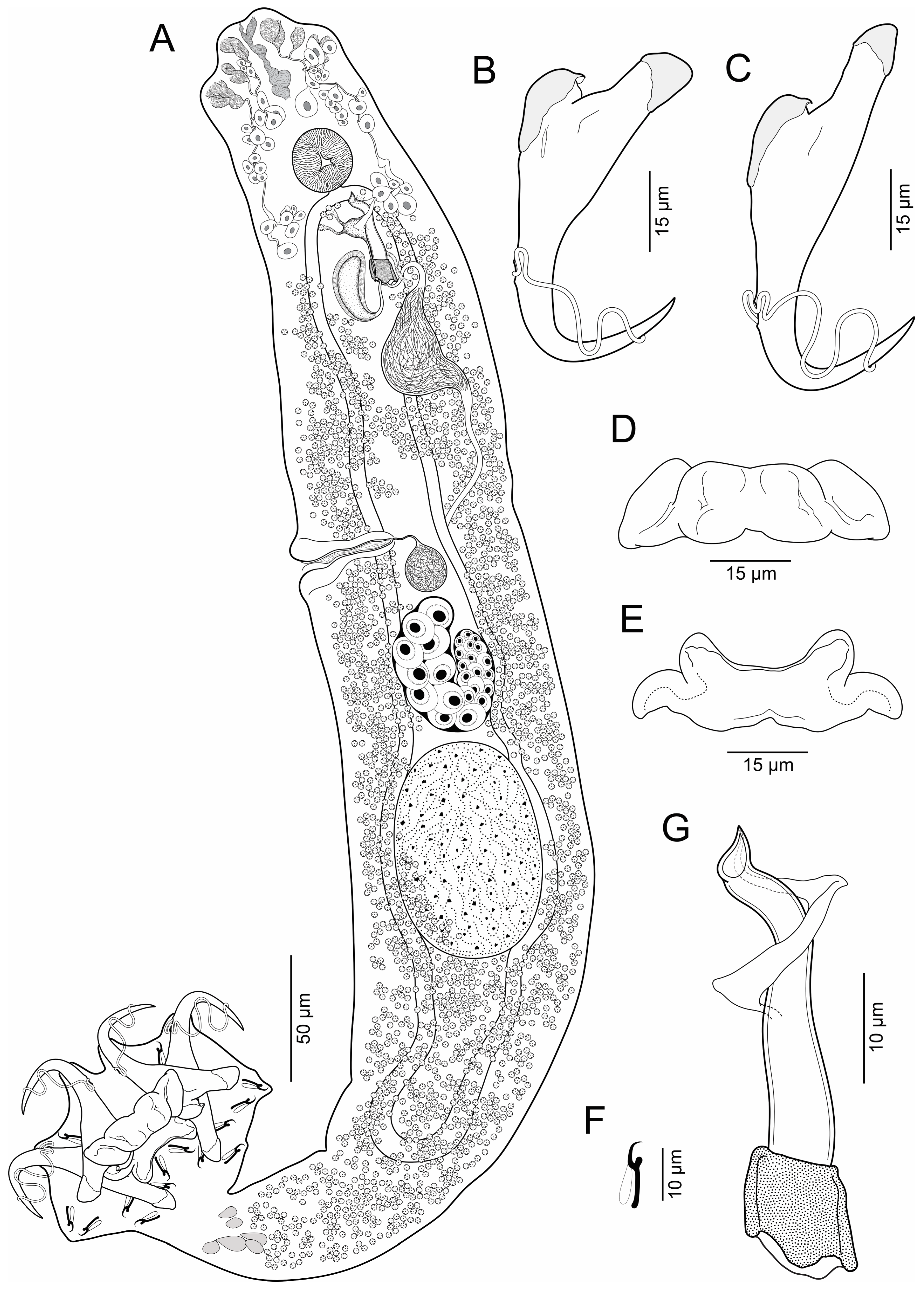
3.1.2. Brotulella luisahelenae n. sp. Cruces, Chero & Luque
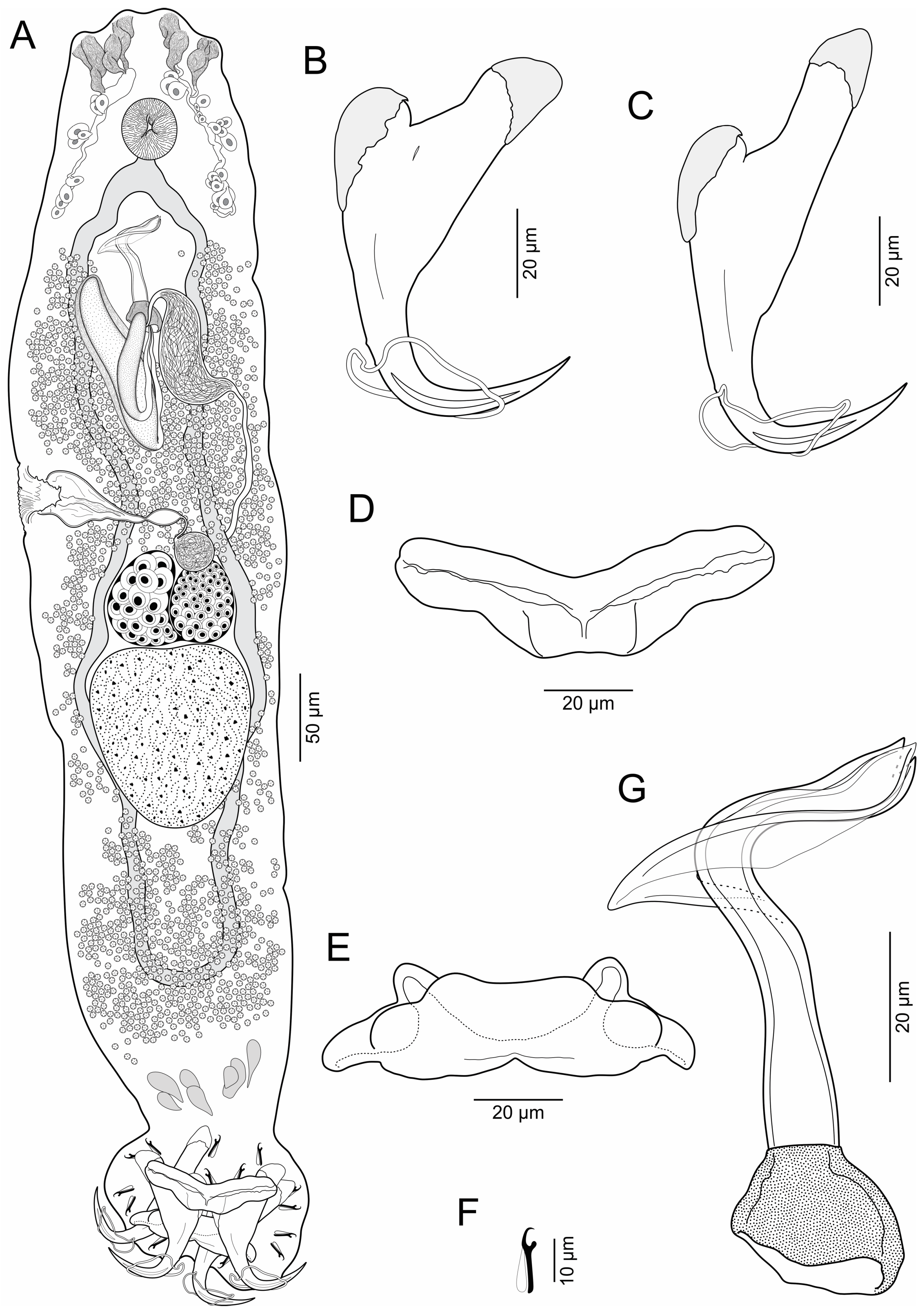
- Body elongates (Figure 2), 625–930 (769, n = 12) long; greatest width 111–153 (132, n = 12) usually at level of seminal vesicle. Cephalic region broad; cephalic lobes poorly developed; cephalic glands bilateral, paired at pre and postpharyngeal level. Pharynx spherical, in greatest width 29–44 (35, n = 8). Peduncle short to elongated. Haptor subquadrangular, 78–109 (94, n = 12) long, 120–165 (139, n = 12) wide; group of inconspicuous secretory gland-cells lying on peduncle. Anchors with fine conspicuous alae; ventral anchor 67–76 (70, n = 7) long, with rounded deep root, robust superficial root, slightly arced shaft, elongated point with furrow on external surface, base 22–26 (24, n = 7) wide; dorsal anchor 80–89 (88; n = 7) long, with almost rounded deep root, elongated superficial root, slightly arced shaft, elongated point with furrow on external surface, base 20–24 (22, n = 7) wide. Ventral bar 60–78 (69, n = 8) long, broadly V-shaped, with moderately enlarged lateral ends. Dorsal bar 56–73 (66, n = 8) long, with almost cat face-shaped, short anterior processes, oblique posterior processes. Hooks 14, similar, 12–15 (14, n = 7) long, each with protruded obtuse thumb, uniform shank, and delicate point; filamentous hook (FH) loop around shank length. MCO 88–107 (96, n = 12) long, tapered, accessory piece with proximal expansion; base of MCO with almost trapezium-shaped. Testis large, intercaecal, subtriangular, not lobulated, 91–129 (108, n = 5) long, 75–97 (88, n = 5) wide; vas deferens dilating to form big seminal vesicle in left side of trunk, posterolateral to MCO; ventral prostatic reservoir elongated, ventral to MCO, dorsal prostatic reservoir well-developed, dextrolateral to MCO. Ovary 42–61 (54, n = 5) long, 58–73 (65, n = 5) wide; oviduct, oötype and uterus not observed. Vaginal vestibule infundibuliform, sclerotized, lying horizontal on right side of body anterior to ovary; vaginal duct running posteriorly to join big subspherical seminal receptacle. Vitelline follicles throughout trunk, lateral fields of follicles confluent posterior to MCO and posterior to testis. Eggs not observed.
- Type host: Brotula clarkae Hubbs, 1944 (Ophidiiformes: Ophidiidae), Pacific bearded brotula.
- Type locality: Puerto Pizarro resort (3°29′ S, 80°24′ W), Tumbes, Peru.
- Site in host: Gills.
- Type specimens: Holotype (MUSM-HEL 5134), 12 paratypes (MUSM-HEL 5135a–k), 1 hologenophore (MUSM 5135l).
- ZooBank registration: The Life Science Identifier (LSID) for Brotulella luisahelenae n. sp. is urn:lsid:zoobank.org:act:6E75215A-471D-490E-809C-4968C7901A34.
- Representative DNA sequence: Sequence was deposited in GenBank under the accession number OR860321 for the 28S rRNA with 720 bp.
- Etymology: The new species is named in honor of Luisa Helena do Amarante Luque, daughter of the senior author.
- Remarks: Brotulella luisahelenae n. sp. can be distinguished from Brotulella laurafernandae n. sp. by its MCO, which have an almost trapezium-shaped base and a blanket-shaped membrane with proximal expansion. In addition, B. luisahelenae n. sp. is typified by having the points of the ventral anchors with a furrow on external surface.
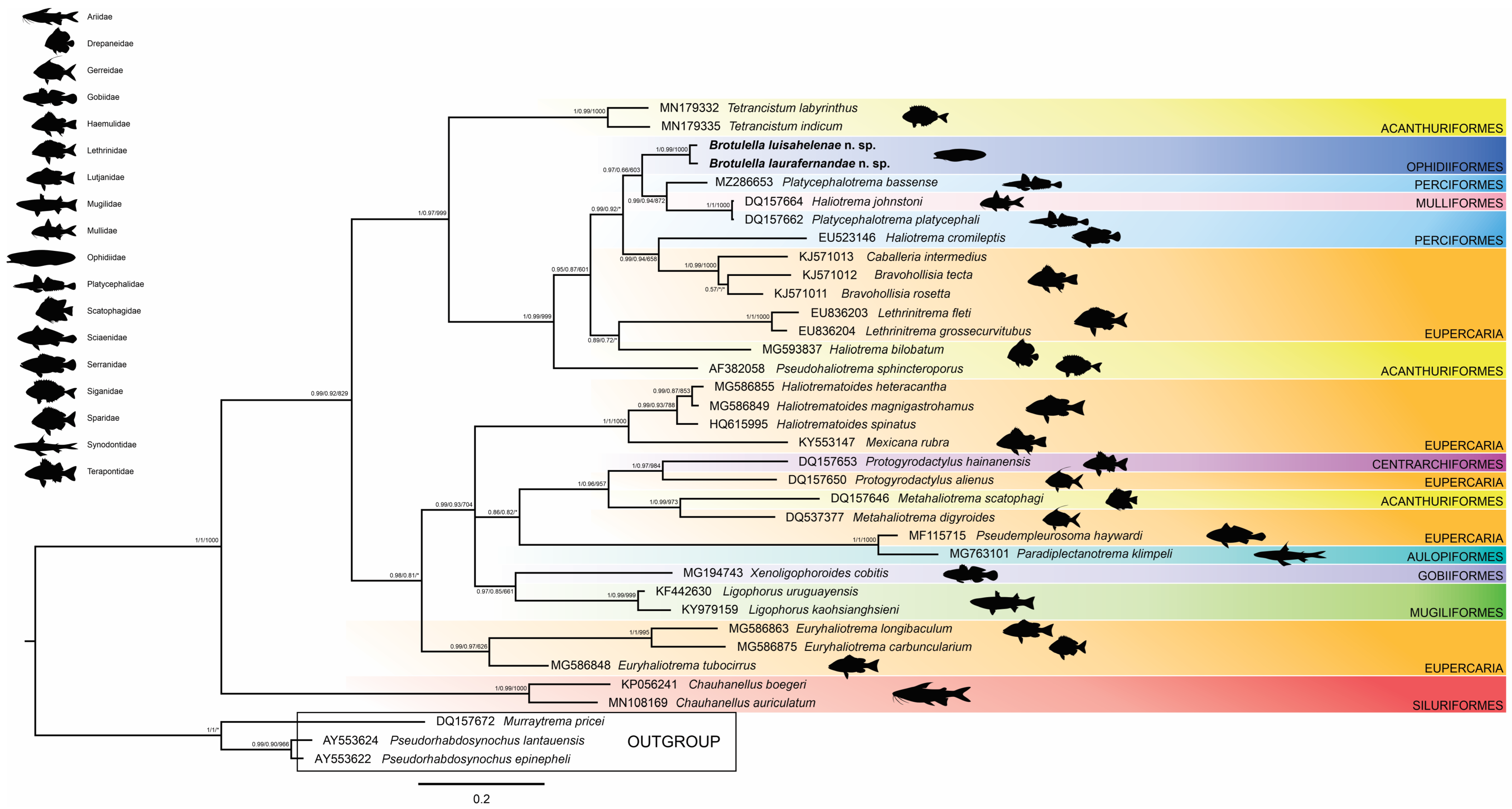
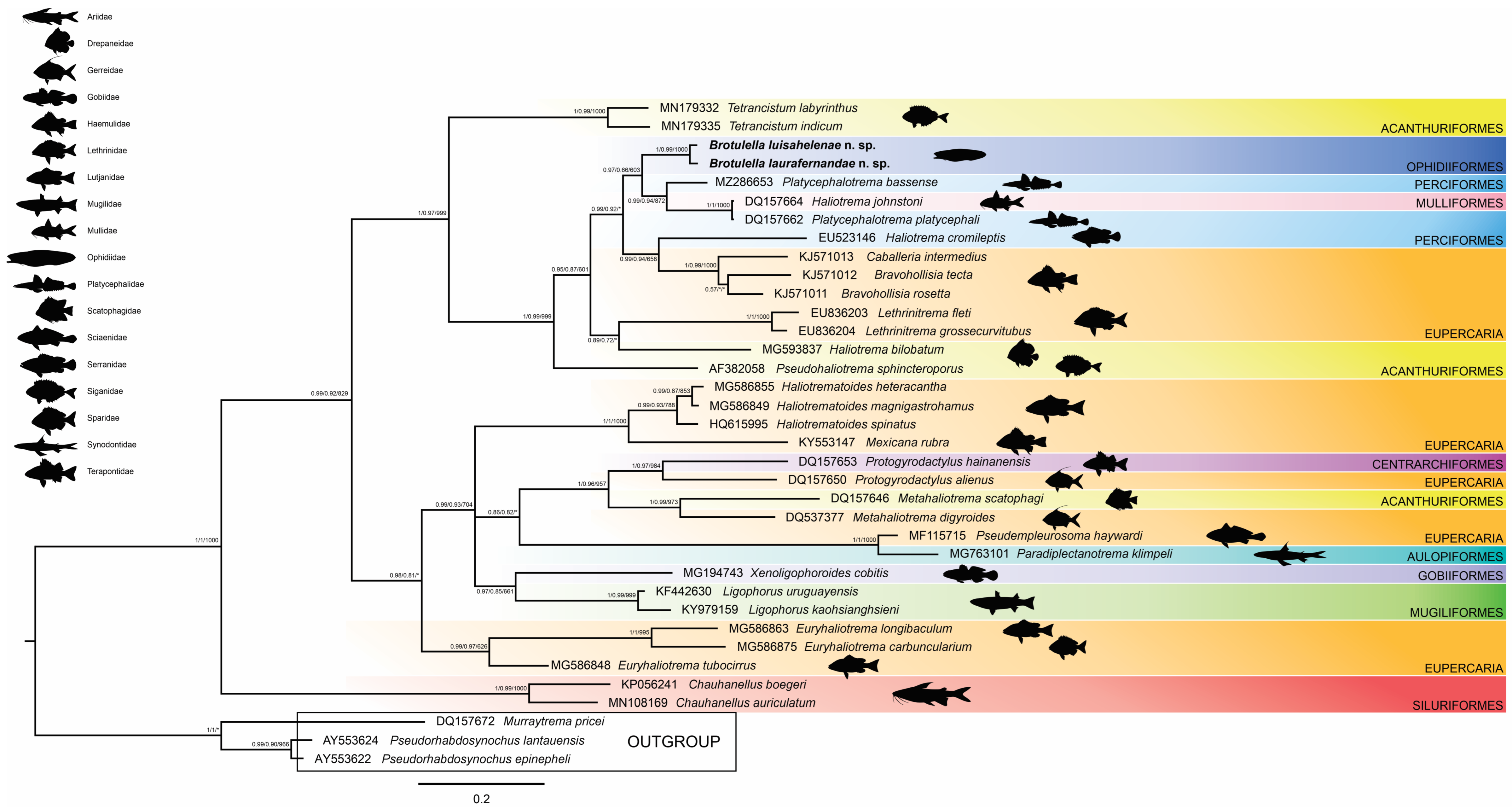
3.2. Phylogenetic Relationships
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marín, A.; Gozzer-Wuest, R.; Grillo-Nuñez, J.; Alavarez-Jaque, I.B.; Riveros, J.C. DNA barcoding reveals overlooked shark and bony fish species in landing reports of small-scale fisheries from northern Peru. Mar. Fish. Sci. 2022, 35, 307–314. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. FishBase. Available online: https://www.fishbase.se/search.php (accessed on 24 May 2023).
- Nature & Culture International. Available online: https://www.natureandculture.org/directory/marine-reserve-peru/ (accessed on 24 May 2023).
- Chero, J.D.; Cruces, C.L.; Sáez, G.; Luque, J.L. Proposal of Cynoscionella n. g. (Monogenea: Diplectanidae), with description of a new species from the gills of Cynoscion phoxocephalus (Actinopterygii: Sciaenidae) in Peru and reassignment of two species of Diplectanum Monticelli, 1903. Syst. Paraitology 2022, 100, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Cruces, C.L.; Chero, J.D.; Sáez, G.; Luque, J.L. Redescription of Haliotrematoides mediohamides (Monogenea: Dactylogyridae), a gill parasite of the Pacific porgy Calamus brachysomus (Perciformes: Sparidae) from the Eastern Pacific Ocean. Syst. Parasitol. 2022, 93, e933960. [Google Scholar]
- Thatcher, V.E. Aquatic Biodiversity in Latin America: Amazon Fish Parasites, 2nd ed.; Pensoft: Sofia, Bulgaria, 2006; 508p. [Google Scholar]
- Tancredo, K.R.; Martins, M.L. Three previous recorded species of Dactylogyrus Diesing, 1850 (Monogenea: Dactylogyridae) infecting cultured Carassius auratus in southern Brazil. J. Parasit. Dis. 2019, 43, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Mbokane, E.M.; Matla, M.M.; Theron, J.; Luus-Powell, W.J. Seasonal dynamics and occurrences of three Dactylogyrus species on the gills of three cyprinids at Nwanedi–Luphephe dams in Limpopo province, South Africa. Afr. Zool. 2015, 50, 119–125. [Google Scholar] [CrossRef]
- Allen, G.R.; Robertson, D.R. Fishes of the Tropical Eastern; Pacific University of Hawaii Press: Honolulu, HI, USA, 1994; 332p. [Google Scholar]
- Nielsen, J.G.; Cohen, D.M.; Markle, D.F.; Robins, C.R. Ophidiiform fishes of the world (Order Ophidiiformes). An annotated and illustrated catalogue of pearlfishes, cusk-eels, brotulas and other ophidiiform fishes known to date. FAO Fish. Synop. 1999, 125, 178. [Google Scholar]
- Herrera, M.; Clarke, T.; Naranjo-Elizondo, B.; Espinoza, M.; Wehrtmann, I.S. Size at maturity of the Pacific bearded brotula (Ophidiidae: Brotula clarkae): A commercially exploited species in the Pacific of Costa Rica. Lat. Am. J. Aquat. Res. 2016, 44, 657–661. [Google Scholar] [CrossRef]
- Ambrose, D.A. Ophidiidae: Cusk-eels. In The Early Stages of Fishes in the California Current Region; Moser, H.G., Ed.; California Cooperative Oceanic Fisheries Investigations (CalCOFI): San Diego, CA, USA, 1996; 1505p. [Google Scholar]
- Chirichigno, N.; Cornejo, R.M. Catálogo Comentado de los Peces Marinos del Perú; Publicación Especial del Instituto del Mar, Instituto del Mar del Perú: Callao, Peru, 2001; 314p. [Google Scholar]
- Lea, R.N.; Allen, M.J.; Power, W. Records of the Pacific bearded brotula, Brotula clarkae, fromSouthern California. Bull. South. Calif. Acad. Sci. 2009, 108, 163–167. [Google Scholar]
- Acevedo, J.; Angulo, W.; Ramírez, M.; Zapata, L.A. Reproduction of the fish Brotula clarkae (Pisces: Ophidiidae) in the Colombian Pacific. Rev. Biol. Trop. 2007, 55, 957–967. [Google Scholar] [CrossRef]
- Chávez-Cevallos, J.M.; Caballero-Vergara, J.A. Análisis del contenido gastrointestinal de la corvina deroca (Brotula clarkae, Hubbs, 1994) desembarcadosen la Playa de Tarqui, Cantón Manta, Provincia de Manabí. Ph.D. Thesis, Universidad Laica Eloy Alfaro de Manabí, Manta, Ecuador, 2008; 84p. [Google Scholar]
- Humason, G.L. Animal Tissue Techniques, 4th ed.; W. H. Freeman and Co.: San Francisco, CA, USA, 1997; 661p. [Google Scholar]
- Mizelle, J.D. New species of trematodes from the gills of Illinois fishes. Am. Midl. Nat. 1936, 17, 785–806. [Google Scholar] [CrossRef]
- Mizelle, J.D.; Price, C.E. Additional haptoral hooks in the genus Dactylogyrus. J. Parasitol. 1963, 19, 785–806. [Google Scholar] [CrossRef]
- Chirichigno, N.; Vélez, M. Clave Para Identificar los peces Marinos del Perú, 2nd ed.; Publicación Especial del Instituto del Mar, Instituto del Mar del Perú: Callao, Peru, 1998; 500p. [Google Scholar]
- Littlewood, D.T.J.; Curini-Galletti, M.; Herniou, E.A. The interrelationships of Proseriata (Platyhelminthes: Seriata) tested with molecules and morphology. Mol. Phylogenet. Evol. 2000, 16, 449–466. [Google Scholar] [CrossRef] [PubMed]
- Lockyer, A.E.; Olson, P.D.; Littlewood, D.T.J. Utility of complete large and small subunit rRNA genes in resolving the phylogeny of the Neodermata (Platyhelminthes): Implications and a review of the cercomer theory. Biol. J. Linn. Soc. 2003, 78, 155–171. [Google Scholar] [CrossRef]
- Olson, P.D.; Cribb, T.H.; Tkach, V.V.; Bray, R.A.; Littlewood, D.T.J. Phylogeny and classification of the Digenea (Platyhelminthes: Trematoda). Int. J. Parasitol. 2003, 33, 733–755. [Google Scholar] [CrossRef]
- Mendoza-Palmero, C.A.; Blasco-Costa, I.; Scholz, T. Molecular phylogeny of Neotropical monogeneans (Platyhelminthes: Monogenea) from catfishes (Siluriformes). Parasites Vectors 2015, 8, 164. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum likelihood phylogenies: Assessing the performance ofPhyML3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Lefort, V.; Longueville, J.E.; Gascuel, O. SMS: Smart Model Selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef]
- Anisimova, M.; Gascuel, O. Approximate likelihood-ratio test for branches: A fast, accurate, and powerful alternative. Syst. Biol. 2006, 55, 539–552. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.; Teslenko, M. Draft MrBayes Version 3.2 Manual: Tutorials and Model Summaries. Available online: https://nbisweden.github.io/MrBayes/manual.html (accessed on 6 April 2023).
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES science gateway: A community resource for phylogenetic analyses. In Proceedings of the 2011 TeraGrid Conference: Extreme Digital Discovery, Salt Lake City, UT, USA, 18–21 July 2011; pp. 1–8. [Google Scholar]
- Kmentová, N.; Cruz-Laufer, A.J.; Pariselle, A.; Smeets, K.; Artois, T.; Vanhove, M.P.M. Dactylogyridae 2022: A meta-analysis of phylogenetic studies and generic diagnoses of parasitic flatworms using published genetic and morphological data. Int. J. Parasitol. 2022, 52, 427–457. [Google Scholar] [CrossRef] [PubMed]
- Yamaguti, S. Monogenetic Trematodes of Hawaiian Fishes; University of Hawaii Press: Honolulu, HI, USA, 1968; 287p. [Google Scholar]
- Justine, J.-L.; Lambert, A.; Mattei, X. Spermatozoon ultrastructure and phylogenetic relationships in the monogeneans (Platyhelminthes). Int. J. Parasitol. 1985, 15, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Perkins, E.M.; Donnellan, S.C.; Bertozzi, T.; Chisholm, L.A.; Whittington, I.D. Looks can deceive: Molecular phylogeny of a family of flatworm ectoparasites (Monogenea: Capsalidae) does not reflect current morphological classification. Mol. Phylogenet. Evol. 2009, 52, 705–714. [Google Scholar] [CrossRef]
- Justine, J.-L.; Poddubnaya, L.G. Spermiogenesis and spermatozoon ultrastructure in basal polyopisthocotylean monogeneans, Hexabothriidae and Chimaericolidae, and their significance for the phylogeny of the Monogenea. Parasite 2018, 25, 7. [Google Scholar] [CrossRef]
- Mollaret, I.; Jamieson, B.G.M.; Adlard, R.D.; Hugall, A.; Lecointre, G.; Chombard, C.; Justine, J.-L. Phylogenetic analysis of the Monogenea and their relationships with Digenea and Eucestoda inferred from 28S rDNA sequences. Mol. Biochem. Parasitol. 1997, 90, 433–438. [Google Scholar] [CrossRef]
- Brabec, J.; Salomaki, E.D.; Kolísko, M.; Scholz, T.; Kutchta, R. The evolution of endoparasitism and complex life cycles in parasitic platyhelminths. Curr. Biol. 2023, 33, 4269–4275. [Google Scholar] [CrossRef]
- Mendoza-Palmero, C.A.; Blasco-Costa, I.; Hernández-Mena, D.; de León, G.P. Parasciadicleithrum octofasciatum n. gen., n. sp. (Monogenoidea: Dactylogyridae), parasite of Rocio octofasciata (Regan) (Cichlidae: Perciformes) from Mexico characterised by morphological and molecular evidence. Parasitol. Int. 2017, 66, 152–162. [Google Scholar] [CrossRef]
- Franceschini, L.; Acosta, A.A.; Zago, A.C.; Müller, M.I.; da Silva, R.J. Trinigyrus spp. (Monogenea: Dactylogyridae) from Brazilian catfishes: New species, molecular data and new morphological contributions to the genus. J. Helminthol. 2020, 94, e126. [Google Scholar] [CrossRef]
- Mendoza-Palmero, C.; Acosta, A.; Scholz, T. Molecular phylogeny of Cosmetocleithrum Kritsky, Thatcher & Boeger, 1986 (Monogenoidea: Dactylogyridae), gill parasites of Neotropical catfishes (Siluriformes). J. Helminthol. 2022, 96, E56. [Google Scholar]
- Ciccheto, J.R.M.; Razzolini, E.L.; de Buron, I.; Boeger, W.A. Position of Polyclithrum within Gyrodactylidae (Monogenoidea): Incongruences between morphological and molecular phylogenies. Syst. Parasitol. 2023, 100, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Oliva, M.E.; Sepulveda, F.A.; González, M. Parapedocotyle prolatili gen. n. et sp. n., a representative of a new subfamily of the Diclidophoridae (Monogenea), a gill parasite of Prolatilus jugularis (Teleostei: Pinguipedidae) from Chile. Folia Parasitol. 2014, 61, 543–548. [Google Scholar] [CrossRef]
- Cohen, S.C.; Justo, M.C.N.; Kohn, A. South American Monogenoidea Parasites of Fishes, Amphibians and Reptiles; Oficina de Livros: Rio de Janeiro, Brazil, 2013; 663p. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Species | Host Species | Locality | GenBank ID |
|---|---|---|---|
| Dactylogyridae | |||
| Bravohollisia roseta | Pomadasys argenteus | Zhanjiang, Guangdong, China | KJ571011 |
| Bravohollisia tecta | Pomadasys maculatus | Zhanjiang, Guangdong, China | KJ571012 |
| Brotulella laurafernandae n. sp. | Brotula clarkae | Off Puerto Pizarro resort, Peru | OR860318 |
| Brotulella luisahelenae n. sp. | Brotula clarkae | Off Puerto Pizarro resort, Peru | OR860321 |
| Caballeria intermedius | Pomadasys argenteus | Zhanjiang, Guangdong, China | KJ571013 |
| Chauhanellus auriculatum | Arius maculatus | Malaysia | MN108169 |
| Chauhanellus boegeri | Genidens genidens | Baia de Antonia, Brazil | KP056241 |
| Euryhaliotrema longibaculum | Lutjanus synagris | Campeche Bank, Mexico | MG586863 |
| Euryhaliotrema carbuncularium | Archosargus rhomboidalis | Campeche Bank, Mexico | MG586875 |
| Euryhaliotrema tubocirrus | Lutjanus griseus | Campeche Bank, Mexico | MG586848 |
| Haliotrema bilobatum | Drepane punctata | Malaysia | MG593837 |
| Haliotrema cromileptis | Epinephelus coioides | Nha Trang Bay, Vietnam | EU523146 |
| Haliotrema johnstoni | Upeneus luzonius | Haikou, Hainan Province, China | DQ157664 |
| Haliotrematoides spinatus | Lutjanus guttatus | Pacific Ocean, off Mexico | HQ615995 |
| Haliotrematoides heteracantha | Lutjanus synagris | Campeche Bank, Mexico | MG586855 |
| Haliotrematoides magnigastrohamus | Lutjanus synagris | Campeche Bank, Mexico | MG586849 |
| Lethrinitrema fleti | Lethrinus nebulosus | China | EU836203 |
| Lethrinitrema grossecurvitubus | Lethrinus nebulosus | China | EU836204 |
| Ligophorus uruguayensis | Mugil liza | Brazil | KF442630 |
| Ligophorus kaohsianghsieni | Planiliza haematocheilus | Russia | KY979159 |
| Metahaliotrema scatophogi | Scatophagus argus | Panyu, Guangdong, China | DQ157646 |
| Metahaliotrema digyroides | Gerres macrosoma | China | DQ537377 |
| Mexicana rubra | Orthopristis ruber | Coast of Rio de Janeiro, Brazil | KY553147 |
| Paradiplectanotrema klimpeli | Saurida tumbil | Indonesia | MG763101 |
| Platycephalotrema bassense | Platycephalus caeruleopunctatus | Australia | MZ286653 |
| Platycephalotrema platycephali | Platycephalus indicus | Weihai, Shangdong, China | DQ157662 |
| Protogyrodactylus alienus | Gerrres filamentosus | China | DQ157650 |
| Protogyrodactylus hainanensis | Therapon jarbua | Yangjiang, Guangdong, China | DQ157653 |
| Pseudohaliotrema sphincteroporus | Siganus doliatus | Australia | AF382058 |
| Pseudempleurosoma haywardi | Johnius amblycephalus | Indonesia | MF115715 |
| Tetrancistrum labyrinthus | Siganus canaliculatus | Oman | MN179332 |
| Tetrancistrum indicum | Siganus canaliculatus | Oman | MN179335 |
| Xenoligophoroides cobitis | Gobius cobitis | Black Sea, off Russia | MG194743 |
| Diplectanidae | |||
| Murraytrema pricei * | Nibea albiflora | Panyu, China | DQ157672 |
| Pseudorhabdosynochus lantauensis * | Epinephelus bruneus | Huidong, China | AY553624 |
| Pseudorhabdosynochus epinepheli * | Epinephelus bruneus | Huidong, China | AY553622 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruces, C.L.; Simões, R.; Maldonado Júnior, A.; Severino, R.; Chero, J.D.; Luque, J.L. Proposal of Brotulella n. gen. for Monogeneans from the Gills of the Pacific Bearded Brotula Brotula clarkae Hubbs, 1944 (Ophidiiformes: Ophidiidae) Based on Morphological and Molecular Evidence. Fishes 2023, 8, 588. https://doi.org/10.3390/fishes8120588
Cruces CL, Simões R, Maldonado Júnior A, Severino R, Chero JD, Luque JL. Proposal of Brotulella n. gen. for Monogeneans from the Gills of the Pacific Bearded Brotula Brotula clarkae Hubbs, 1944 (Ophidiiformes: Ophidiidae) Based on Morphological and Molecular Evidence. Fishes. 2023; 8(12):588. https://doi.org/10.3390/fishes8120588
Chicago/Turabian StyleCruces, Celso Luis, Raquel Simões, Arnaldo Maldonado Júnior, Ruperto Severino, Jhon Darly Chero, and José Luis Luque. 2023. "Proposal of Brotulella n. gen. for Monogeneans from the Gills of the Pacific Bearded Brotula Brotula clarkae Hubbs, 1944 (Ophidiiformes: Ophidiidae) Based on Morphological and Molecular Evidence" Fishes 8, no. 12: 588. https://doi.org/10.3390/fishes8120588
APA StyleCruces, C. L., Simões, R., Maldonado Júnior, A., Severino, R., Chero, J. D., & Luque, J. L. (2023). Proposal of Brotulella n. gen. for Monogeneans from the Gills of the Pacific Bearded Brotula Brotula clarkae Hubbs, 1944 (Ophidiiformes: Ophidiidae) Based on Morphological and Molecular Evidence. Fishes, 8(12), 588. https://doi.org/10.3390/fishes8120588