

Effect on Muscle Cellularity of Diet Supplementation with Nannochloropsis gaditana Microalgae in the Final Fattening Phase of Gilthead Seabream Culture up to Commercial Size
 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rearing Conditions
2.2. Experimental Diets
2.3. Sampling
2.4. Analysis of Muscle Growth
2.5. Statistical Analysis
3. Results
3.1. Body Growth and Survival
- R2.5 versus R5 groups: When comparing raw diets of N. gaditana at different concentrations, the body values were similar in both groups (p > 0.05) (Table 2);
- R2.5 versus H2.5 groups: When comparing diets at 2.5% of raw versus hydrolyzed N. gaditana, the body weight showed similar values in both groups (p > 0.05) (Table 2);
- H2.5 versus H5 groups: When comparing hydrolyzed diets at different concentrations, the highest values of the body weight were reached at highest concentration, even though it was not significant (p > 0.05) (Table 2);
- R5 versus H5 groups: H5 showed the highest values of the body weight, even though it was not significant (p > 0.05) (Table 2);
- C group: The lowest body weight values were observed in this group, but it was not significant (p > 0.05) (Table 2).
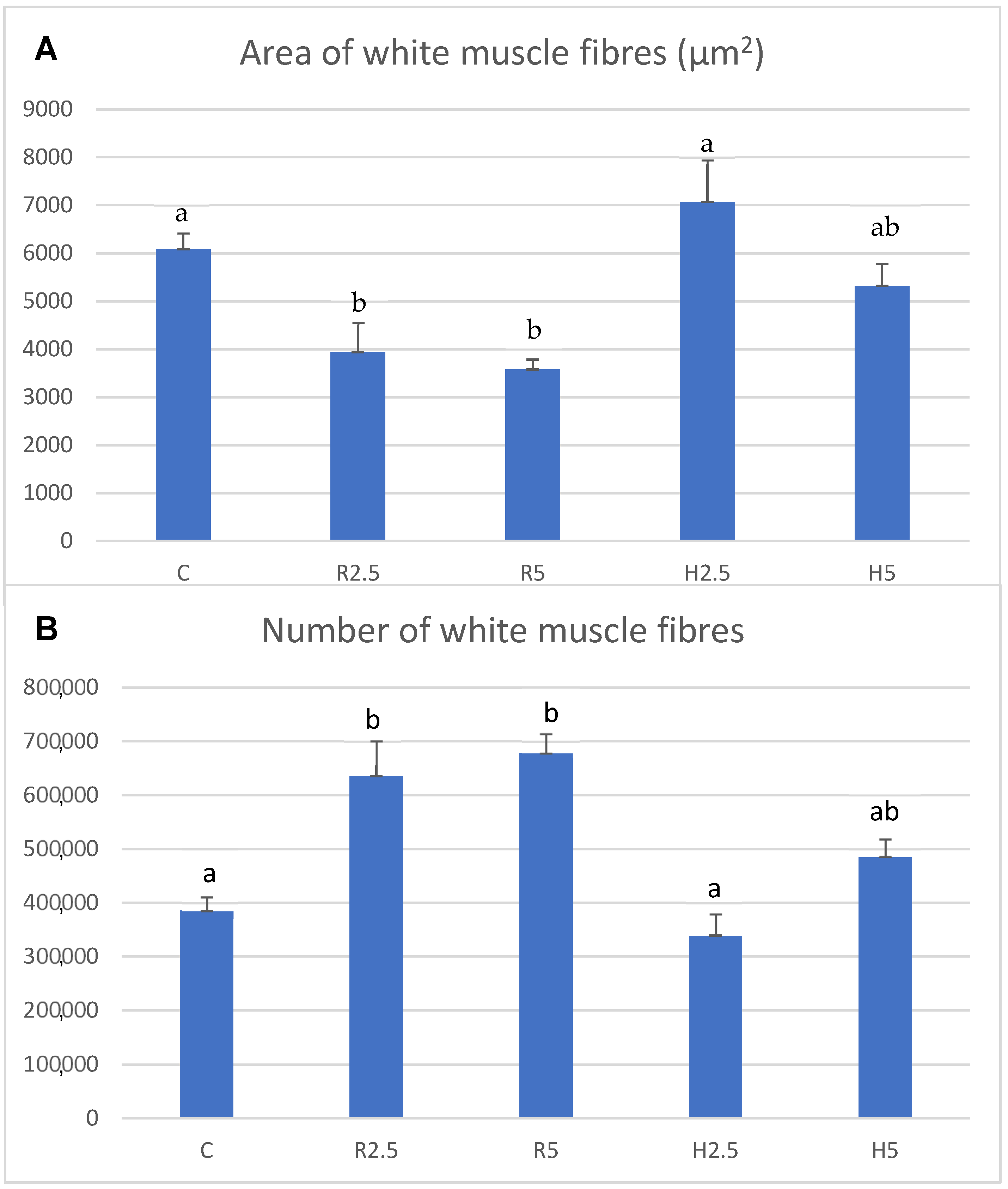
3.2. Muscle Growth
4. Discussion
5. Conclusions
- Enriched diets with N. gaditana did not significantly influence body growth (length and weight) or the transverse area of the white muscle in the final phase of fattening of gilthead seabream specimens.
- The form of N. gaditana (raw versus hydrolyzed) in the diet significantly influenced the muscle cellularity of gilthead seabream adults at commercial size, in such a way that the hypertrophy was higher in fish fed with hydrolyzed microalgae than in fish fed with raw microalgae.
- The enzymatic treatment of N. gaditana did not enhance the generation of new fibers, so that the greatest hyperplasia was reached by the fish fed with raw microalgae and it was correlated with the highest fillet hardness values previously found by other authors in these specimens [26].
- No significant differences attributable to the concentration levels of N. gaditana were observed. This result, together with the fact that the greatest generation of fibers was found in the diets of raw N. gaditana, allows us to conclude that the R2.5 diet could be the most optimal for this phase in sea bream.
- The short-term effect of the microalgae-enriched diets used in the final phase of fattening of sea breams of this study should be considered by farmers to optimize culture and to improve the quality of fillet meat.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, F.; Leng, Y.; Lu, Q. The application of microalgae biomass and bio-products as aquafeed for aquaculture. Algal Res. 2021, 60, 102541. [Google Scholar] [CrossRef]
- Krogdahl, Å.; Penn, M.; Thorsen, J.; Refstie, S.; Bakke, A.M. Important antinutrients in plant feedstuffs for aquaculture: An update on recent findings regarding responses in salmonids. Aquac. Res. 2010, 41, 333–344. [Google Scholar] [CrossRef]
- Ponis, E.; Robert, R.; Parisi, G. Nutritional value of fresh and concentrated algal diets for larval and juvenile Pacific oysters (Crassostrea gigas). Aquaculture 2003, 221, 491–505. [Google Scholar]
- Walker, A.; Berlinsky, D. Effects of partial replacement of fish meal protein by microalgae on growth, feed intake, and body composition of Atlantic cod. N. Am. J. Aquac. 2011, 73, 76–83. [Google Scholar]
- Tulli, F.; Chini Zittelli, G.; Giorgi, G.; Poli, B.M.; Tibaldi, E.; Tredici, M.R. Effect of the inclusion of dried Tetraselmis suecica on growth, feed utilization and fillet composition of European sea bass juveniles fed organic diets. J. Aquat. Food Prod. Technol. 2012, 21, 188–197. [Google Scholar] [CrossRef]
- Tibaldi, E.; ChiniZittelli, G.; Parisi, G.; Bruno, M.; Giorgi, G.; Tulli, F.; Venturini, S.; Tredici, M.R.; Poli, B.M. Growth performance and quality traits of European sea bass (D. labrax) fed diets including increasing levels of freez-dried Isochrysis sp. (T-ISO) biomass as a source protein and n-3 long chain PUFA in partial substitution of fish derivatives. Aquaculture 2015, 440, 60–66. [Google Scholar]
- Vizcaíno, A.; López, G.; Sáez, M.; Jiménez, J.; Barros, A.; Hidalgo, L.; Camacho-Rodríguez, J.; Martínez, T.; Cerón-García, M.; Alarcón, F. Effects of the microalga Scenedesmus almerienses as fishmeal alternative in diets for gilthead sea bream, Sparus aurata, juveniles. Aquaculture 2014, 431, 34–43. [Google Scholar] [CrossRef]
- Vizcaíno, A.J.; Saéz, M.I.; López, G.; Arizcun, M.; Abellán, E.; Martínez, T.F.; Cerón-García, M.C.; Alarcón, F.J. Tetraselmis suecia and Tisochrysis lutea meal as dietary ingredients for gilthead sea bream (Sparus aurata L.) fry. J. Appl. Phycol. 2016, 28, 2843–2855. [Google Scholar]
- Miller, M.R.; Nichols, P.D.; Carter, C.G. Replacement of fish oil with Thraustochytrid schizochytrium sp. L oil in Atlantic salmon parr (Salmo salar L.) diets. Comp. Biochem. Physiol. A 2007, 148, 382–392. [Google Scholar]
- Atalah, E.; Cruz, C.M.H.; Izquierdo, M.S.; Rosenlund, G.; Caballero, M.J.; Valencia, A.; Robaina, L. Two microalgae Crypthecodinium cohnii and Phaeodactylum tricornutum as alternative source of essential fatty acids in starter feeds for sea bream (Sparus aurata). Aquaculture 2007, 270, 178–185. [Google Scholar]
- Ganuza, E.; Benítez-Santana, T.; Atalah, E.; Vega-Orellana, O.; Ganga, R.; Izquierdo, M.S. Crypthecodinium cohnii and Schizochytrium sp. as a potential substitutes to fisheries-derived oils from seabream (Sparus aurata) microdiets. Aquaculture 2008, 277, 109–116. [Google Scholar]
- Eryalcin, K.M.; Roo, J.; Saleh, R.; Atalah, E.; Benítez, R.; Betancor, M.; Hernández-Cruz, C.M.; Izquierdo, M.S. Fish oil replacement by different microalgal products in microdiets for early weaning of gilthead sea bream (Sparus aurata, L.). Aquacult. Res. 2013, 44, 819–828. [Google Scholar] [CrossRef]
- Cahu, C.L.; Zambonino Infante, J.L.; Péres, A.; Quazuguel, P.; Le Gall, M.M. Algal addition in sea bass (Dicentrarchus labrax) larvae rearing: Effect on digestive enzymes. Aquaculture 1998, 161, 479–489. [Google Scholar]
- Karapanagiotidis, I.T.; Metsoviti, M.N.; Gkalogianni, E.Z.; Psofakis, P.; Asimaki, A.; Katsoulas, N.; Papapolymerou, G.; Zarkadas, I. The effects of replacing fishmeal by Chlorella vulgaris and fish oil by Schizochytrium sp. and Microchloropsis gaditana blend on growth performance, feed efficiency, muscle fatty acid composition and liver histology of gilthead seabream (Sparus aurata). Aquaculture 2022, 561, 738709. [Google Scholar]
- Seong, T.; Uno, Y.; Kitagima, R.; Kabeya, N.; Haga, Y.; Satoh, S. Microalgae as main ingredient for fish feed: Non-fish meal and non-fish oil diet development for red sea bream, Pagrus major, by blending of microalgae Nannochloropsis, Chlorella and Shizochytrium. Aquacult. Res. 2021, 52, 6025–6036. [Google Scholar] [CrossRef]
- Carvalho, M.; Izquierdo, M.; Valdés, M.; Montero, D.; Farías, A. Oils Combination with Microalgal Products as a Strategy for Increasing the N-3 Long-Chain Polyunsaturated Fatty Acid Content in Fish Oil-Free Diets for Meagre (Argyrosomus regius). Aquac. Nutr. 2022, 2022, 5275570. [Google Scholar] [CrossRef]
- Johnston, I.A.; Alderson, R.; Sandham, C.; Dingwall, A.; Mitchell, D.; Selkirk, C.; Nickell, D.; Baker, R.; Robertson, B.; Whyte, D.; et al. Patterns of muscle growth in early and late maturing populations of Atlantic salmon (Salmo salar L.). Aquaculture 2000, 189, 307–333. [Google Scholar]
- Johnston, I.A. Muscle development and growth: Potential implications for flesh quality in fish. Aquaculture 1999, 177, 99–115. [Google Scholar]
- Johnston, I.A.; Manthri, S.; Alderson, R.; Campbell, P.; Mitchell, D.; Whyte, D.; Dingwall, A.; Nickell, D.; Selkirk, C.; Robertson, B. Effects of dietary protein level on muscle cellularity and flesh quality in Atlantic salmon with particular reference to gaping. Aquaculture 2002, 210, 259–283. [Google Scholar] [CrossRef]
- Johnston, I.A.; Manthri, S.; Alderson, R.; Smart, A.; Campbell, P.; Nickell, D.; Robertson, B.; Paxton, C.G.; Burt, M.L. Freshwater environment affects growth rate and muscle fibre recruitment in seawater stages of Atlantic salmon (Salmo salar L.). J. Exp. Biol. 2003, 206, 1337–1351. [Google Scholar] [CrossRef]
- Johnston, I.A.; Manthri, S.; Smart, A.; Campbell, P.; Nickell, D.; Alderson, R. Plasticity of muscle fibre number in seawater stages of Atlantic salmon in response to photoperiod manipulation. J. Exp. Biol. 2003, 206, 3425–3435. [Google Scholar] [CrossRef]
- Fauconneau, B.; Andre, S.; Chmaitilly, P.Y.; Krieg, F.; Kaushik, S.J. Control of skeletal muscle fibres and adipose cells in the flesh of rainbow trout. J. Fish Biol. 1997, 50, 296–314. [Google Scholar] [CrossRef]
- Knutsen, H.R.; Johnsen, I.H.; Keizer, S.; Sorensen, M.; Roques, J.A.C.; Heden, I.; Sundell, K. Fish welfare, fast muscle cellularity, fatty acid and body-composition of juvenile spotted wolffish (Anarhichas minor) fed a combination of plant proteins and microalgae (Nannochloropsis oceanica). Aquaculture 2019, 506, 212–223. [Google Scholar] [CrossRef]
- Ayala, M.D.; Galián, C.; Fernández, V.; Chaves-Pozo, E.; García de la Serrana, D.; Sáez, M.I.; Galafat Díaz, A.; Alarcón, F.J.; Martínez, T.F.; Arizcun, M. Influence of Low Dietary Inclusion of the Microalga Nannochloropsis gaditana (Lubián 1982) on Performance, Fish Morphology, and Muscle Growth in Juvenile Gilthead Seabream (Sparus aurata). Animals 2020, 10, 2270. [Google Scholar] [CrossRef] [PubMed]
- Ayala, M.D.; Balsalobre, N.; Chaves-Pozo, E.; Sáez, M.I.; Galafat, A.; Alarcón, F.J.; Martínez, T.F.; Arizcun, M. Long-term Effects of a short juvenile feeding period with diets enriched with the microalgae Nannochloropsis gaditana on the subsequent body and muscle growth of gilthead seabream, Sparus aurata L. Animals 2023, 13, 482. [Google Scholar] [CrossRef]
- Sáez, M.J.; Galafat, A.; Suárez, M.D.; Chaves-Pozo, E.; Arizcun, M.; Ayala, M.D.; Alarcón, F.J.; Martínez, T.F. Effects of raw and hydrolysed Nannochloropsis gaditana biomass included at low level in finishing diets for gilthead seabream (Sparus aurata) on fillet quality and shelf life. J. Appl. Phycol. 2023, 35, 1163–1181. [Google Scholar] [CrossRef]
- Sáez, M.I.; Galafat, A.; Vizcaíno, A.J.; Chaves-Pozo, E.; Ayala, M.D.; Arizcun, M.; Alarcón, F.J.; Suárez, M.D.; Martínez, T.F. Evaluation of Nannochloropsis gaditana raw and hydrolysed biomass at low inclusion level as dietary functional additive for gilthead seabream (Sparus aurata) juveniles. Aquaculture 2022, 556, 738288. [Google Scholar] [CrossRef]
- Ayala, M.D.; Abellán, E.; Arizcun, M.; García-Alcázar, A.; Navarro, F.; Blanco, A.; López-Albors, A. Muscle development and body growth in larvae and early postlarvae of shi drum, Umbrina cirrosa L., reared under different larval photoperiod. Muscle structural and ultrastructural study. Fish Physiol. Biochem. 2013, 39, 807–827. [Google Scholar] [CrossRef]
- Ayala, M.D.; Arizcun, M.; García-Alcázar, A.; Santaella, M.; Abellán, E. Long-term effects of the larval photoperiod on the subsequent growth of shi drum Umbrina cirrosa L. specimens and the fillet texture at commercial size. Turk. J. Fish Aquat. 2015, 15, 93–101. [Google Scholar]
- Romanello, M.G.; Scapolo, P.A.; Luprano, S.; Mascarello, F. Post-larval growth in the lateral white muscle of the eel, Anguilla anguilla. J. Fish. Biol. 1987, 30, 161–172. [Google Scholar] [CrossRef]
- Scapolo, P.A.; Veggetti, A.; Mascarello, F.; Romanello, M.G. Developmental transitions of myosin isoforms and organisation of the lateral muscle in the teleost Dicentrarchus labrax (L.). Anat. Embryol. 1988, 178, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Veggetti, A.; Mascarello, F.; Scapolo, P.A.; Rowlerson, A. Hyperplastic and hypertrophic growth of lateral muscle in Dicentrarchus labrax (L.): An ultrastructural and morphometric study. Anat. Embryol. 1990, 182, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Higgins, P.J.; Thorpe, J.E. Hyperplasia and hypertrophy in the growth of skeletal muscle in juvenile Atlantic salmon (Salmo salar, L.). J. Fish Biol. 1990, 37, 505–519. [Google Scholar] [CrossRef]
- Johnston, I.A. Environment and plasticity of myogenesis in teleost fish. J. Exp. Biol. 2006, 209, 2249–2264. [Google Scholar] [CrossRef] [PubMed]
- Hatae, K.; Yoshimatsu, F.; Matsumoto, J.J. Discriminative characterization of different texture profiles of various cooked fish muscles. J. Food Sci. 1984, 49, 721–726. [Google Scholar] [CrossRef]
- Hatae, K.; Yoshimatsu, F.; Matsumoto, J.J. Role of muscle fibres in contributing firmness of cooked fish. J. Food Sci. 1990, 55, 693–696. [Google Scholar] [CrossRef]
- Periago, M.J.; Ayala, M.D.; López-Albors, O.; Abdel, I.; Martínez, C.; García-Alcázar, A.; Ros, G.; Gil, F. Muscle cellularity and flesh quality of wild and farmed sea bass, Dicentrarchus labrax L. Aquaculture 2005, 249, 175–188. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Diets | |||||
|---|---|---|---|---|---|
| Ingredient Composition (g kg−1 Dry Matter) | C | R2.5 | R5 | H2.5 | H5 |
| Fish meal LT94 1 | 150 | 150 | 150 | 150 | 150 |
| Raw N. gaditana 2 | - | 25 | 50 | - | - |
| Hydrolysed N. gaditana | - | - | - | 25 | 50 |
| Squid meal 3 | 20 | 20 | 20 | 20 | 20 |
| CPSP90 4 | 10 | 10 | 10 | 10 | 10 |
| Krill meal 5 | 20 | 20 | 20 | 20 | 20 |
| Gluten meal 6 | 150 | 150 | 150 | 150 | 150 |
| Soybean protein concentrate 7 | 400 | 388 | 373 | 388 | 373 |
| Fish oil 8 | 114 | 110 | 105 | 110 | 105 |
| Soybean lecithin 9 | 10 | 10 | 10 | 10 | 10 |
| Wheat meal 10 | 54 | 45 | 40 | 45 | 40 |
| Choline chloride 11 | 5 | 5 | 5 | 5 | 5 |
| Betain 12 | 5 | 5 | 5 | 5 | 5 |
| Lysine 13 | 15 | 15 | 15 | 15 | 15 |
| Methionine 14 | 6 | 6 | 6 | 6 | 6 |
| Vitamin and mineral premix 15 | 20 | 20 | 20 | 20 | 20 |
| Vitamin C 16 | 1 | 1 | 1 | 1 | 1 |
| Guar gum 17 | 20 | 20 | 20 | 20 | 20 |
| Crude protein | 452 | 461 | 464 | 454 | 459 |
| Crude lipid | 158 | 151 | 155 | 154 | 151 |
| Ash | 71 | 79 | 83 | 76 | 85 |
| Moisture | 61 | 58 | 53 | 59 | 49 |
| Groups | C | R2.5 | R5 | H2.5 | H5 |
|---|---|---|---|---|---|
| BL (cm) | 31.19 a ± 0.13 | 31.29 a ± 0.16 | 31.37 a ± 0.12 | 31.27 a ± 0.11 | 31.27 a ± 0.12 |
| BW (g) | 487.38 a ± 5.48 | 494.74 a ± 5.61 | 494.51 a ± 5.00 | 492.79 a ± 5.08 | 502.31 a ± 5.79 |
| Groups | C | R2.5 | R5 | H2.5 | H5 |
|---|---|---|---|---|---|
| W (mm2) | 2265.29 a ± 58.5 | 2247.61 a ± 39.4 | 2388.56 a ± 40.5 | 2347.68 a ± 87.1 | 2472.26 a ± 63.2 |
| D (µm) | 56.69 a ± 2.32 | 39.93 b ± 5.31 | 36.98 b ± 1.80 | 65.16 a ± 5.89 | 48.95 ab ± 2.32 |
| Dens | 167.14 a ± 8.95 | 285.35 b ± 31.21 | 286.04 b ± 15.84 | 151.14 a ± 20.59 | 199.31 ab ± 17.51 |
| G | C | R2.5 | R5 | H2.5 | H5 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| P | Area | D | Area | D | Area | D | Area | D | Area | D |
| 5 | 50.23 | 5.76 | 7.75 | 1.89 | 7.35 | 1.79 | 36.38 | 5.19 | 10.73 | 2.47 |
| 10 | 165.61 | 10.20 | 9.74 | 2.39 | 9.14 | 2.28 | 97.37 | 8.39 | 13.52 | 2.99 |
| 20 | 588.11 | 20.85 | 12.72 | 2.99 | 11.93 | 2.84 | 578.97 | 21.78 | 19.88 | 3.79 |
| 25 | 1011.98 | 26.46 | 14.91 | 3.31 | 13.32 | 3.08 | 1398.81 | 33.45 | 24.05 | 4.32 |
| 30 | 1437.19 | 32.90 | 17.29 | 3.64 | 14.71 | 3.42 | 2093.45 | 42.12 | 31.41 | 5.10 |
| 40 | 2607.08 | 43.89 | 29.82 | 4.96 | 20.67 | 4.19 | 3452.80 | 54.69 | 258.25 | 13.39 |
| 50 | 4167.69 | 54.85 | 395.43 | 17.42 | 367.19 | 15.73 | 5088.07 | 66.86 | 1680.52 | 36.08 |
| 60 | 6033.88 | 63.81 | 2089.30 | 41.73 | 1907.63 | 39.96 | 7759.92 | 77.72 | 3991.25 | 56.70 |
| 70 | 8076.40 | 72.49 | 4973.99 | 62.70 | 4207.55 | 58.41 | 10,267.34 | 91.19 | 6452.29 | 72.87 |
| 75 | 9388.52 | 78.17 | 6357.26 | 71.61 | 5634.99 | 68.02 | 11,428.18 | 98.88 | 7970.48 | 80.89 |
| 80 | 10,750.09 | 84.30 | 7886.20 | 79.99 | 7616.89 | 77.91 | 13,131.05 | 105.03 | 9763.42 | 89.24 |
| 90 | 14,134.55 | 101.16 | 11,873.16 | 99.40 | 11,908.83 | 99.30 | 17,302.84 | 123.35 | 13,648.51 | 108.61 |
| 95 | 17,056.17 | 116.78 | 15,153.39 | 114.80 | 14,877.44 | 114.95 | 21,566.41 | 139.69 | 17,056.76 | 122.93 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayala, M.D.; Chaves-Pozo, E.; Sáez, M.I.; Galafat, A.; Alarcón, F.J.; Martínez, T.F.; Arizcun, M. Effect on Muscle Cellularity of Diet Supplementation with Nannochloropsis gaditana Microalgae in the Final Fattening Phase of Gilthead Seabream Culture up to Commercial Size. Fishes 2023, 8, 532. https://doi.org/10.3390/fishes8110532
Ayala MD, Chaves-Pozo E, Sáez MI, Galafat A, Alarcón FJ, Martínez TF, Arizcun M. Effect on Muscle Cellularity of Diet Supplementation with Nannochloropsis gaditana Microalgae in the Final Fattening Phase of Gilthead Seabream Culture up to Commercial Size. Fishes. 2023; 8(11):532. https://doi.org/10.3390/fishes8110532
Chicago/Turabian StyleAyala, María Dolores, Elena Chaves-Pozo, María Isabel Sáez, Alba Galafat, Francisco Javier Alarcón, Tomás Francisco Martínez, and Marta Arizcun. 2023. "Effect on Muscle Cellularity of Diet Supplementation with Nannochloropsis gaditana Microalgae in the Final Fattening Phase of Gilthead Seabream Culture up to Commercial Size" Fishes 8, no. 11: 532. https://doi.org/10.3390/fishes8110532
APA StyleAyala, M. D., Chaves-Pozo, E., Sáez, M. I., Galafat, A., Alarcón, F. J., Martínez, T. F., & Arizcun, M. (2023). Effect on Muscle Cellularity of Diet Supplementation with Nannochloropsis gaditana Microalgae in the Final Fattening Phase of Gilthead Seabream Culture up to Commercial Size. Fishes, 8(11), 532. https://doi.org/10.3390/fishes8110532