1. Introduction
The Chinese mitten crab (
Eriocheir sinensis) is an important freshwater cultured crustacean in China [
1].
E. sinensis has excellent taste and good nutritional properties. The hepatopancreas, gonads, and muscles are the main edible parts of
E. sinensis, which are rich in a variety of essential amino acids (EAA) and polyunsaturated fatty acids (PUFA) and have positive pharmacological and physiological effects on human health [
2,
3]. Therefore, the analysis of muscle and hepatopancreas components is important to assess the edible quality and nutritional value of
E. sinensis.
Although numerous studies have described the culture technology of
E. sinensis at home and abroad, the appropriate breeding mode of
E. sinensis often varies substantially due to the influence of different water environments [
4,
5,
6]. Therefore, it is necessary to research the key factors in the culture process. It is known that in fish culture, an increase in the stocking density will intensify the intraspecific struggle of cultured organisms; for example, by increasing the competition pressure for food and space, which will affect their survival, growth, and physiology, and will also aggravate the deterioration of the quality of cultured water [
7,
8,
9]. Previous studies have reported that stocking density affects the survival rate, growth, breeding quality, and aquaculture water environment of shrimp and crab culture, and this effect is negatively correlated [
10,
11,
12,
13]. The generally accepted explanation is that the high-density living environment will intensify the competition for water space and food resources, thereby affecting its feeding, growth, and energy metabolism, such as
Mithraculus forceps [
14] and
Palaemonetes sinensis [
15]. In addition, intensive farming at high density can also cause stress responses in shrimp and crabs [
16,
17]. Considering the above-mentioned reasons, the importance of studying the appropriate stocking density of
E. sinensis becomes evident.
In recent years, the aquaculture area and output of E. sinensis in Jiangxi Province have increased annually, and aquaculture methods have diversified. However, few systematic reports on the healthy breeding of E. sinensis in Jiangxi have been reported. Therefore, Poyang Lake was selected as the focus of research in the current study, and the influence of stocking density on growth performance, nutritional quality, and immune-related gene expression of E. sinensis in the pond ecosystem was analyzed. According to the actual production of E. sinensis cultured in the Poyang Lake area, five density groups were set up, namely, G1-G5(G1: 1.2 inds/m2; G2: 1.5 inds/m2; G3: 1.8 inds/m2; G4: 2.1 inds/m2; G5: 2.4 inds/m2). This study provides basic scientific data for the healthy culture of the Chinese mitten crab in the Poyang Lake area and promotes the development of E. sinensis farming in the area.
2. Materials and Methods
2.1. Experimental Protocol and Sample Collection
This study followed the guidelines and was approved by the Animal Care and Use Committee of Nanchang University, and was performed in a crab breeding base of Jiangxi province. The experimental ponds (1000–1333 m
2) are earth ponds with sufficient water sources, and the water quality complies with the requirements of water quality standards for fisheries and aquaculture (
Table 1). The crabs (
E. sinensis, a novel variety “Yangtze River No. 2”) were obtained from Ludong, Jiangsu Province, in early February, and healthy
E. sinensis (6.32 ± 1.19 g) with robust physique, agile crawling, healthy appendages, no external injuries, and no parasite attachment were selected as the breeding test crab species. Basic information on the
E. sinensis species and ponds is provided in
Table 2. The
E. sinensis were randomly divided into five groups (G1–G5) based on the breeding densities of 1.2, 1.5, 1.8, 2.1, and 2.4 inds/m
2, respectively. Three parallel ponds of similar area were set for each density group, and the breeding trial period was 169 d.
2.2. Aquaculture Water Quality Measurements
On days 0, 30, 60, 120, and 150 of the breeding experiment, the water quality of the ponds was analyzed. With the water inlet, outlet, and central location of each test pond as fixed monitoring points (sampling points), three samples were taken from each pond, and nine samples were taken from three ponds of each density group for measurement. Water samples were taken directly from the preset position in 250 mL brown glass bottles, sealed, and stored away from light for nitrite nitrogen (NO2-N, mg/L) analysis, 1-L water collectors were used to take the water sample at the preset position, and concentrated sulfuric acid was added at a ratio of 1:500, sealed, and stored away from light for ammonia nitrogen (NH3-N, mg/L) analysis. Water temperature (WT, °C), dissolved oxygen (DO, mg/L), pH, and electrical conductivity (EC, μs/cm) were measured using a YSI portable multifunctional water quality analyzer (Professional Plus, USA). Ammonia nitrogen (NH3-N, mg/L), nitrite nitrogen (NO2-N, mg/L), and other indicators were analyzed and determined using a Hach DR3900 multifunctional water quality analyzer and its supporting kits.
2.3. Growth Performance Measurements
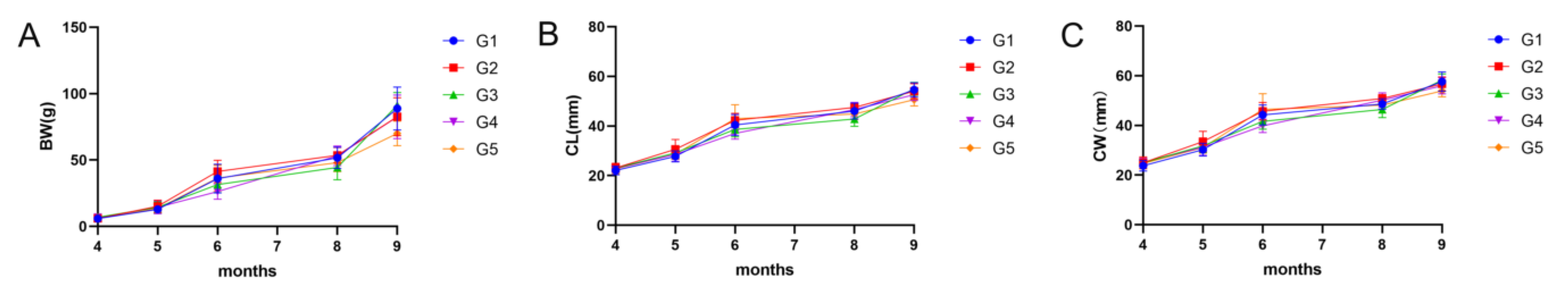
On days 0, 30, 60, 120, and 150 of the breeding experiment, 30
E. sinensis were randomly selected, respectively, from each group to record basic growth data. Body weight (BW) was determined using electronic scales (with an accuracy of 0.01 g), and carapace length (CL) and width (CW) were measured using electronic Vernier calipers (accurate to 0.001 cm). The average value of each index at day 0 and d 150 was used to calculate the weight gain rate (WGR), specific growth values (SGR), carapace length growth rate (GR
CL) and width (GR
CW), the mean BW and CL at day 150 of each group were used to calculate condition factors (CF), and the average value and standard deviation of each index at five sampling times were used to calculate standard deviation coefficient of variance (CV) as in the following formulas:
where, W
i is the average value of the initial wet weight (g); W
f, the average value of the final wet weight (g); T, the time interval in d; CL
i and CL
f, the average value of the initial and final carapace length, respectively; and CW
i and CW
f, the average value of the initial and final carapace width. SD is standard deviation, MN is the mean of BW, CL, and CW of each test pool crab, respectively.
2.4. Nutritional Composition Measurements
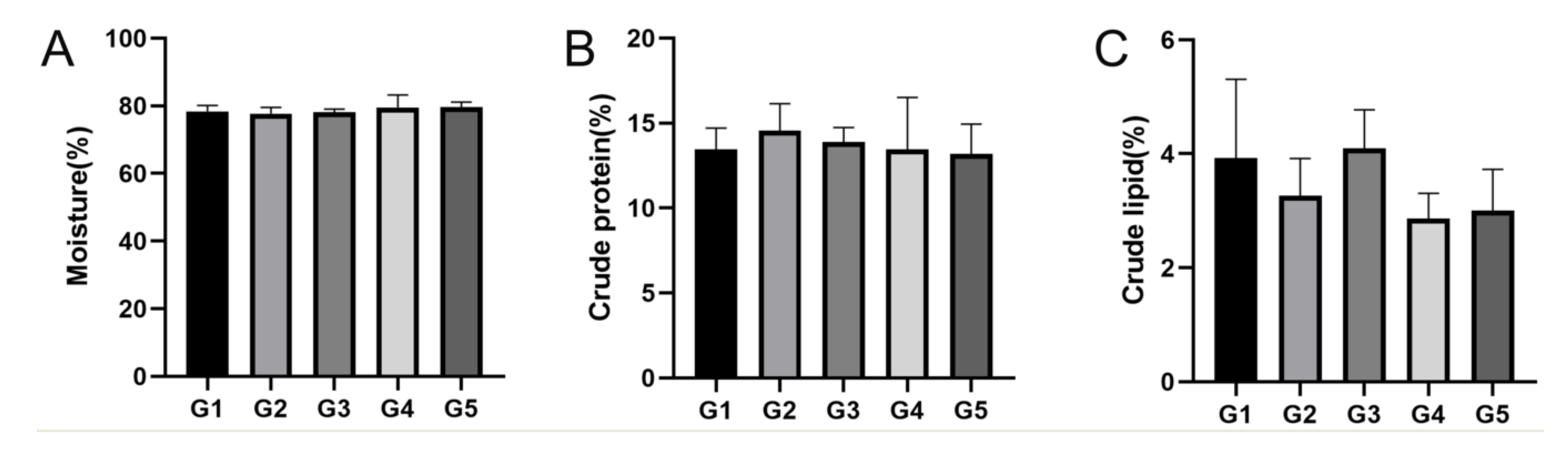
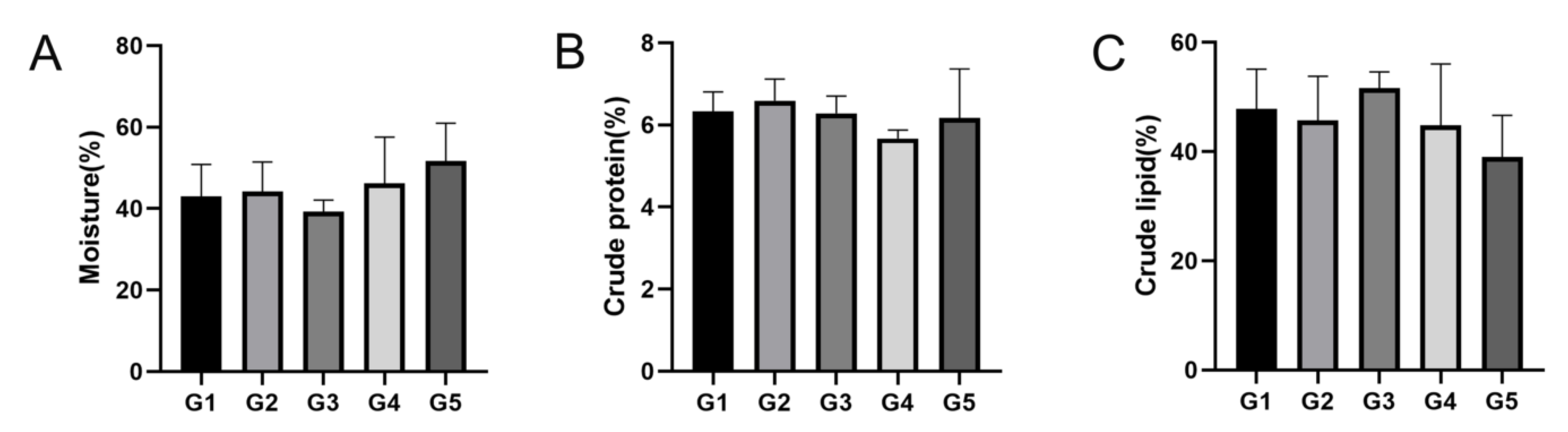
At the conclusion of the breeding experiment, 10 E. sinensis’ muscle and hepatopancreatic tissue were randomly collected from each pond to analyze the nutritional composition. Moisture was detected using the 105 °C constant temperature drying weight loss method (GB5 009.3-2016), using an electronic balance with an accuracy of 0.001 g. Crude fat content was evaluated using the Soxhlet extraction method (GB5 009.6-2016), the equipment uses a set of cord extractors. Crude protein content was determined using the Kjeldahl method (GB5 009.5-2016), using a Haineng K1160 fully automated total nitrogen analyzer. The amino acid compositions were analyzed and determined using a Hitachi L8900 automatic amino acid analyzer.
2.5. Measurements of Immune-Related Gene Expression Levels
At the conclusion of the breeding experiment, 3
E. sinensis’ hemolymphs were randomly collected from each pond to analyze the immune-related gene expression. Total RNA was extracted from the hemolymph using TRIzol. cDNA was prepared using a PrimeScript RT reagent Kit with a gDNA Eraser kit (TaKaRa), and cDNA samples were used as a qRT-PCR template. The Bio-Rad CFX Manager 3.1 real-time PCR instrument was used for relative quantification under the following reaction conditions: 95 °C for 10 min, followed by 39 cycles, 95 °C for 30 s, 58–60 °C for 30 s, and 72 °C for 30 s. Melting reaction conditions were 65–95 °C, with plate reading for 30 s to record the amount of fluorescence. The PCR primer sequences of
LBGP,
Tube,
Dorsal,
PPAE,
SOD,
GST, and
GPx genes are listed in
Table 3. β-actin was used as the internal reference gene. The relative changes in gene expression levels were detected using the 2
−ΔΔ-CT method. The experiment performs a triplicate assay on each sample.
2.6. Statistical Analysis
Statistical analyses were carried out using SPSS 26.0 software (IBM Corporation, New Orchard Road, Armonk, NY 10504, USA, 2019). Data are presented as mean ± standard deviation (SD). Percentage data were square-root arcsine transformed to achieve homogeneity of variance and then statistically analyzed. One-way ANOVA and Duncan’s multiple comparisons were used to compare significant differences between groups. The significance level was set at p < 0.05.
The R package “HH” of ANCOVA was utilized to detect if water quality (pH, and NO2-N) is a cofactor for growth and immunity, in which growth index, amino acid composition, and expression of immune related genes are dependent variables, density is an independent variable, and environmental factors are covariates. The significance level was set at p < 0.05, and the extremely significant level was set at p < 0.01.
4. Discussion
Stocking density is a crucial factor that affects aquatic animals, and numerous studies have revealed that a high stocking density can negatively impact the growth, nutritional composition, and immune responses of aquatic animals [
12,
18,
19,
20]. In the white shrimp
Litopenaeus vannamei (Boone), the lower stocking densities had higher final body weights than the higher density groups, while the highest density stocking can cause serious damage to the ability of their immune systems [
21]. In
Palaemonetes sinensis, the final body weight and weight gain rate showed a tendency to increase and then decrease with increasing stocking density, and the activity of digestion-related enzymes and some non-specific immune indicators decreased significantly with increasing stocking density [
22]. In rice and crab coculture systems with different stocking densities, both growth performance and nutrient fractions tend to increase and then decrease with increasing stocking density [
23]. In addition, a suitable breeding density is crucial for controlling the quality of aquaculture water [
24]. Aquatic organisms need adequate levels of dissolved oxygen to facilitate growth and prevent disease [
25]. Studies have reported that intensive crab farming at high density may cause the water quality of lakes to deteriorate [
26]. At the same time, farming shrimp and crab at an appropriate density level is not only conducive to improving water quality, but also helps to improve the growth, nutrition, and immune response of shrimps and crabs [
27]. To date, few studies have analyzed the growth performance, nutritional quality, and immune-related gene expression of
E. sinensis in pond ecosystems under the influence of stocking density. Therefore, it is of urgency to explore the appropriate stocking density of aquatic animals. In this study, the water quality of all experimental ponds conformed to the Class III fishery and surface water quality standards, and the results illustrate that stocking density had a certain impact on the growth, nutrient composition, and expression of immune-related genes in
E. sinensis. Crustaceans grow by molting, and
E. sinensis go through 18–21 molts over their lifespan [
28]. In this study, four molting processes were observed. Research on the growth performance of
E. sinensis is mainly reflected in aspects of BW, SGR, CL, CW, and CF [
29]. In this study, final BW, WGR, SGR, GR
CL, and GR
CW are higher in the lower density group than in the higher density group, indicating that controlling the stocking density within a certain range can effectively enhance the growth performance of Chinese mitten crabs. Liu et al. found that the final BW, WGR, and SGR of rice crabs decreased with increasing stocking density [
30], which is in accordance with the results of this study. Peters et al. reported that the CW/CL of
E. sinensis is a constant value of approximately 1.13, which can be used as an evaluation indicator for germplasm identification [
31]. However, the mean value of CW/CL of
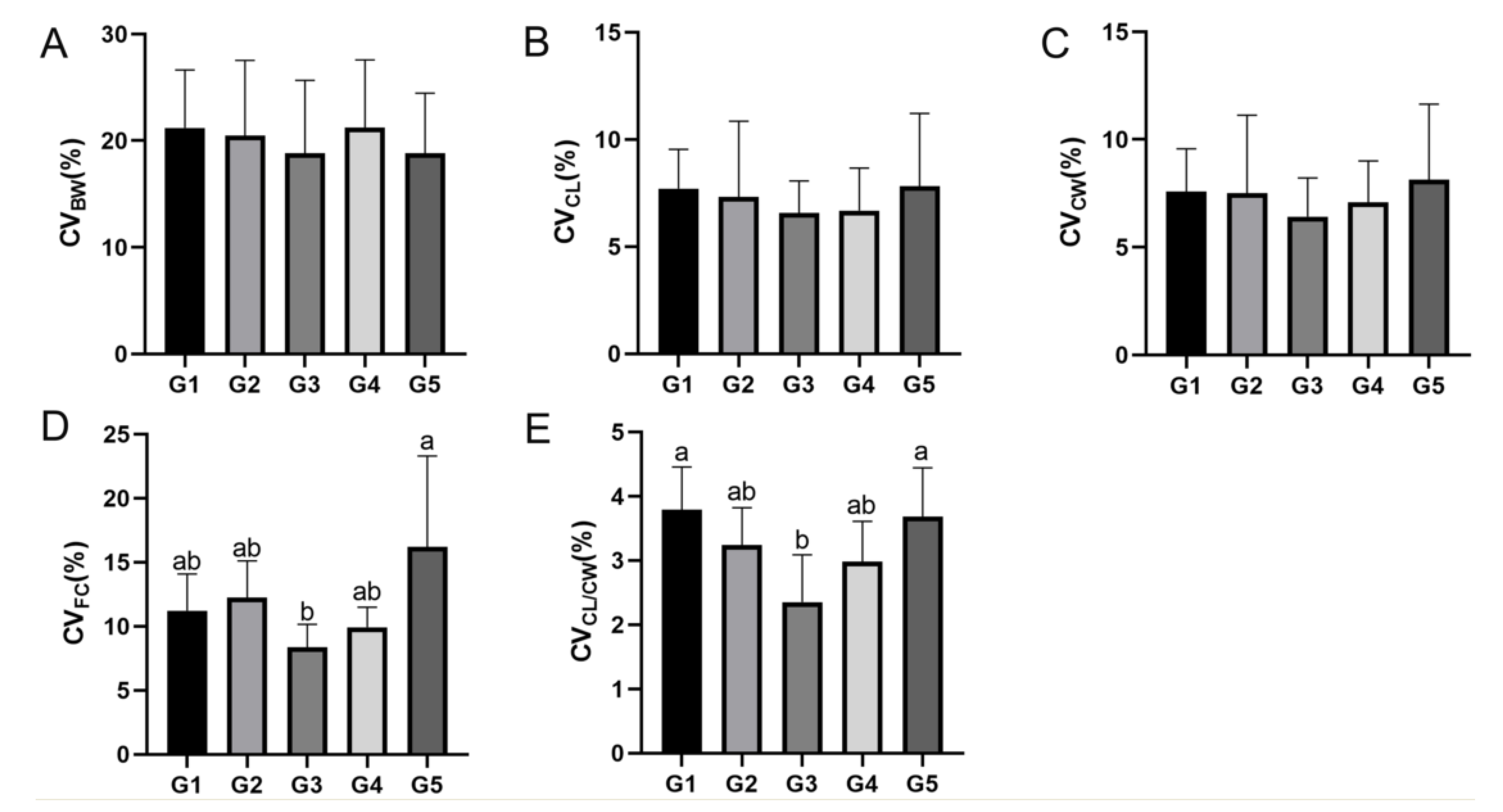
E. sinensis measured in this study was 1.07–1.08, less than 1.13, and the mean value of CW/CL increased with an increase in stocking density, but there was no significant difference. The reasons for this result may be related to the novel variety “Changjiang No. 2” used in this study. Compared with the wild variety, “Changjiang No. 2” was obtained through five generations of group breeding, and has the characteristics of fast growth, large individuals, and excellent quality. In addition, the values of CV
BW, CV
CL, CV
CW, CV
CF, and CV
CW/CL in the G3 group were lower than those in the other groups, suggesting that the size and nutrient accumulation of
E. sinensis in G3 were largely uniform during the breeding process, and the stocking density had a certain influence on the growth and nutrient accumulation of
E. sinensis. In fish, size differences are eventually magnified as the fish grow [
32], while no significant effect of initial size on the growth of juvenile
Pacific abalone was observed [
33]. In this study, the G3 group had a higher starting body weight than the other groups, but no significant difference in final body weight. These results indicate that initial size and weight may have no significant effect on the growth in invertebrates.
Salas-Leiton et al. have shown that high stocking density may also lead to intensified intraspecific struggles of Senegalese sole (
Solea senegalensis), such as intensifying competition for space and food, which may further affect feeding growth, behavior, energy metabolism, and final nutrient accumulation in the body [
34]. In the current study, the nutritional composition of
E. sinensis, including proximate and amino acid composition, influenced by stocking density in the pond ecosystem was also analyzed. The crude protein and lipid contents decreased as stocking density increased, and, in the muscle and hepatopancreas tissues, the crude lipid content in G3 was higher than that in the G1, G2, G4, and G5 groups (
p > 0.05). In the muscle tissue, the content of numerous amino acids, including leucine, lysine, EAA, TAA, and DAA in G3 were higher than those in the other groups, which is in line with the results of Wang et al. who studied the nutritional components of rice crab culture at different stocking densities, and showed that the crude protein content and EAA levels of
E. sinensis cultured at a density of 0.4 inds/m
2 were higher than those in other stocking densities [
35]. These results suggest that high stocking density negatively affects the nutritional composition of
E. sinensis.
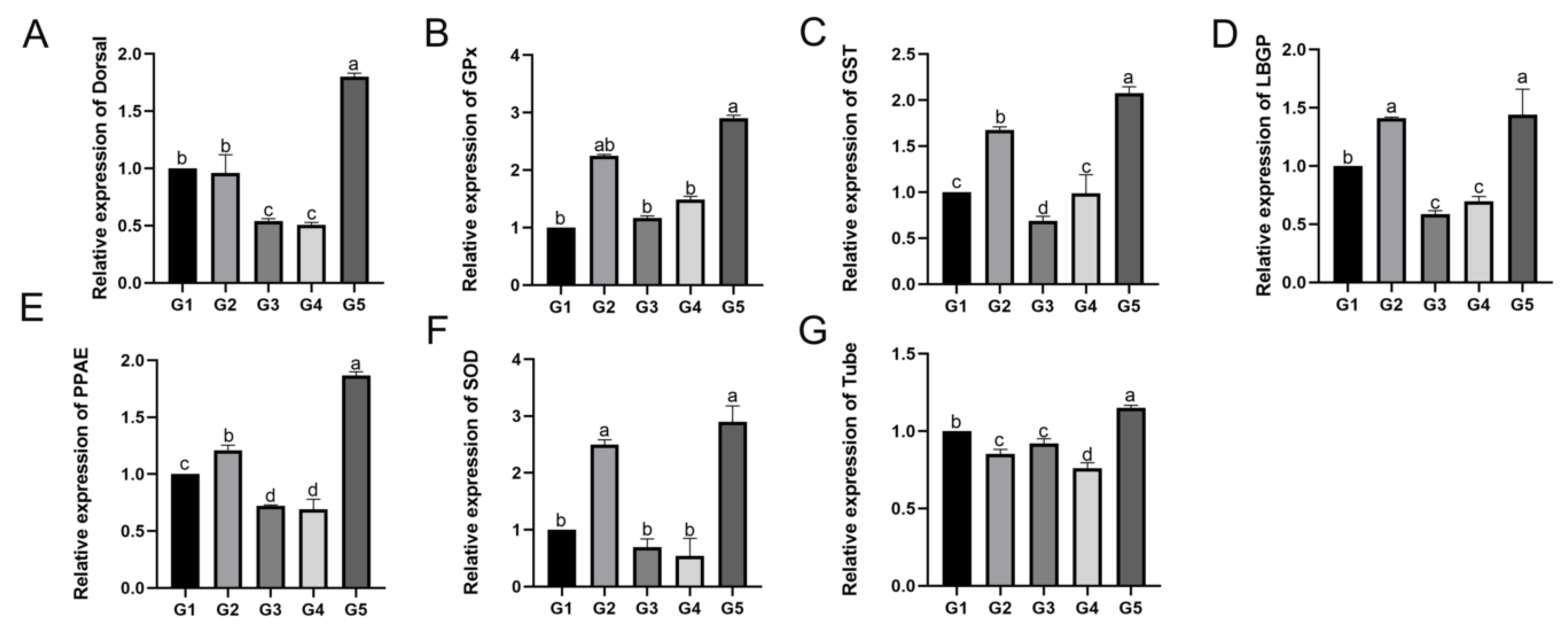
The hemolymph is an important immune organ of shrimps and crabs; thus, immune-related gene expression was evaluated in the hemolymph, including
LBGP,
Tube,
Dorsal,
PPAE,
SOD,
GST, and
GPx. LGBP is a classic pattern recognition protein in crustaceans that can activate the prophenoloxidase system and is immune-specific to Gram-negative bacteria and fungi [
36].
Dorsal and
Tube are important mediator genes and core components in the Toll signaling pathway and play significant roles in immune signaling [
37,
38]. PPAE is a key component of the prophenoloxidase activation system; serine protease homologous (SPH) is a vital cofactor of PPAE, and a previous study has revealed that SPH plays a significant role in the innate immune response [
39]. SOD, GPx, and GST are important antioxidant enzymes in crustaceans, among which, GPx is an important reactive oxygen radical scavenger in the body [
40], and forms an antioxidant defense system together with SOD [
41] and catalase. GST is widely present in various organisms, and is a multifunctional antioxidant enzyme that participates in the metabolism of a variety of endogenous or exogenous toxic substances [
42]. In this study, from the point of view of molecular level, it is shown that the culture density has some effect on the immune system of
E. sinensis. Both too low or too high density farming had some stressful effects on
E. sinensis, while the expression of immune genes had lower levels in the G3 and G4 density groups, which showed that the stress of these two culture density groups was less than other groups.
The results of this study show that the stocking density not only affected the growth performance of
E. sinensis, but also affected the water quality of the aquaculture pond. With the increase in density, pH decreased and NO
2-N increased significantly in the G5 group. Some studies reported the effects of pH and NO
2-N on the growth, nutrition, and immunity of aquatic animals. Long WC et al. found that too low a pH negatively affected the growth of
Lithodes aequispinus [
43]; pH (pH 6 and 8.5) stress significantly altered the expression of EsCaM mRNA in
E. sinensis gills, hepatopancreas, blood cells, intestine, and muscle [
44]; excessive levels of NO
2-N can negatively affect the growth of aquatic animals, for example,
Macrobrachium rosenbergii [
45],
Litopenaeus schmitti [
46] and
Lateolabrax Japonicus [
47]. In order to better clarify whether the changes in the growth indicators of
E. sinensis were only affected by the stocking density factor or were affected by the combination of the cultivation density and water environment factors in this study, ANCOVA was performed with pH and NO
2-N as covariates, density as the independent variable, and growth performance-related indexes as dependent variables. The results showed that, as a synergetic factor of culture density, the pH value of the culture pond affected the growth performance of Chinese mitten-handed crab, while NO
2-N had no significant effect on the growth performance-related index of
E. sinensis.
This study has some limitations. First, we did not conduct a comprehensive analysis and evaluation of the health and growth benefits of E. sinensis under different stocking densities in this region. Thus, other key influencing factors in the pond culture of E. sinensis, such as research on crab specifications, water quality, and bait, should be further supplemented to improve relevant data. In addition, in-depth research into the molecular mechanisms that influence stocking density of the E. sinensis is required.



 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}