
Dietary Supplementation with Prebiotic Chitooligosaccharides Enhances the Growth Performance, Innate Immunity and Disease Resistance of Nile Tilapia (Oreochromis niloticus)





Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Husbandry and Bacterial Strain
2.2. Preparation of the Experimental Diet and Feeding Trial
2.3. Growth Parameters
2.4. Determination of Digestive Enzyme Activities in the Gut
2.5. Evaluation of Innate Immune Parameters
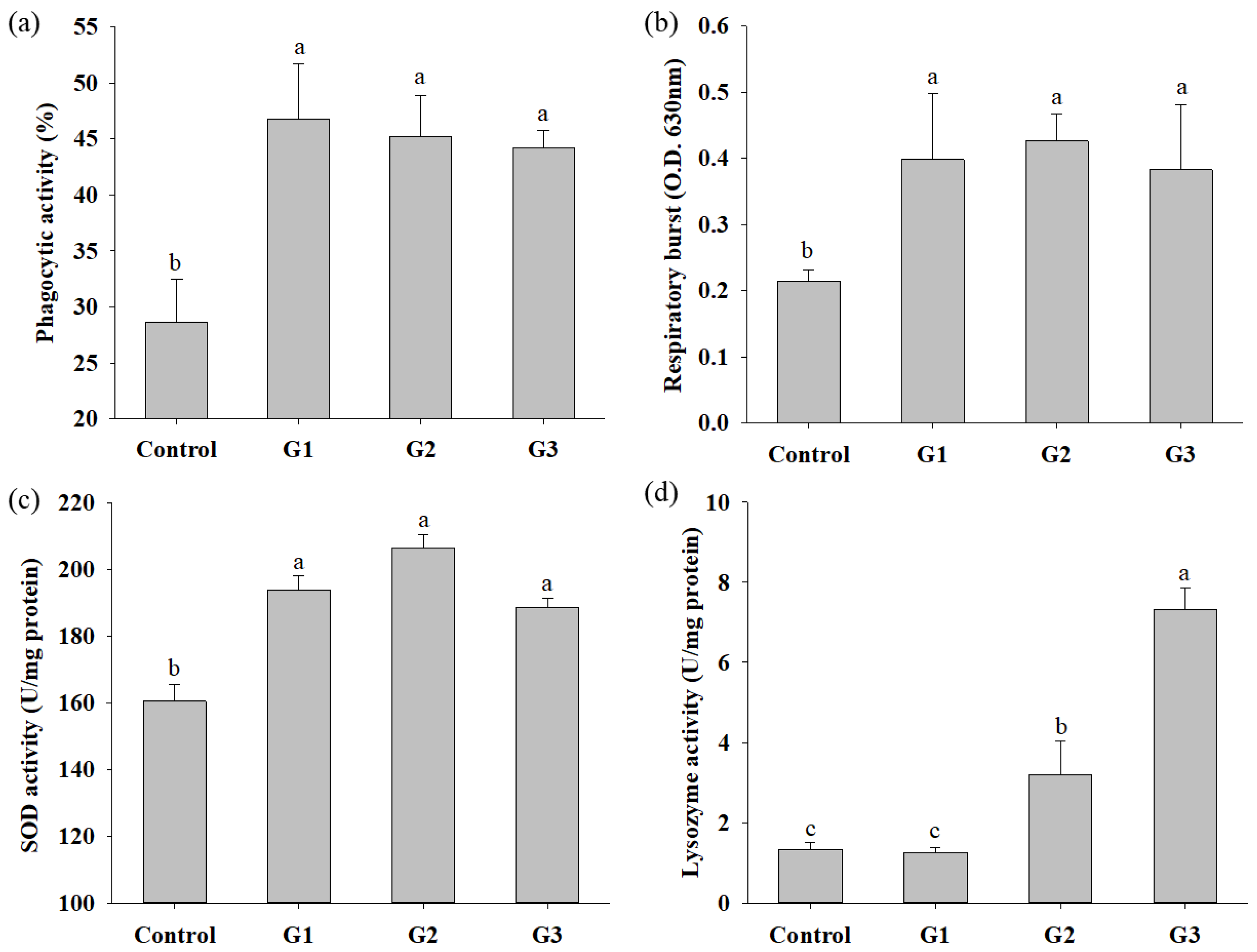
2.5.1. Phagocytic Activity Assay
2.5.2. Respiratory Burst Activity Assay
2.5.3. SOD and Lysozyme Activity Assay
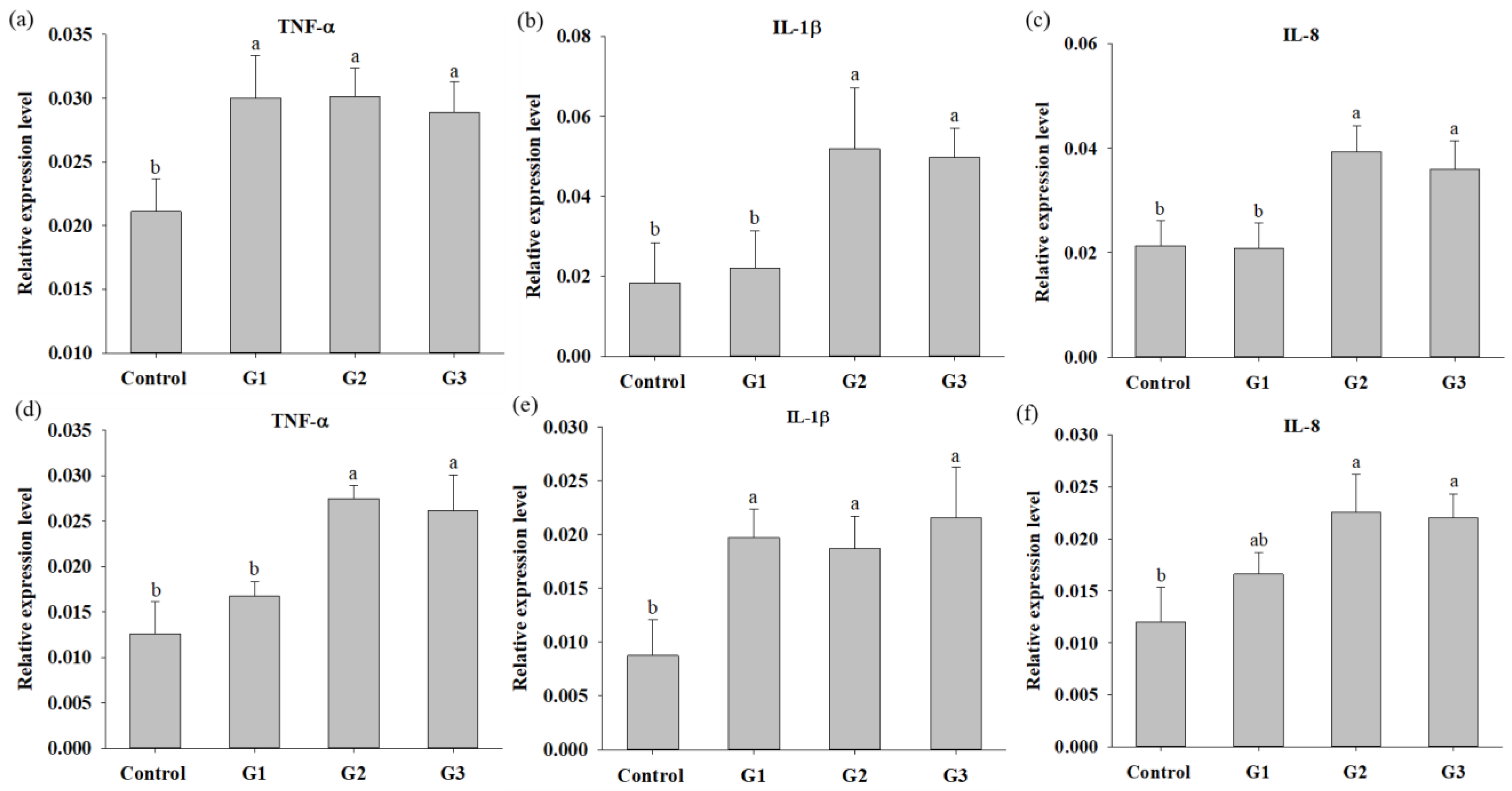
2.6. Determination of Indicator Gene Expression
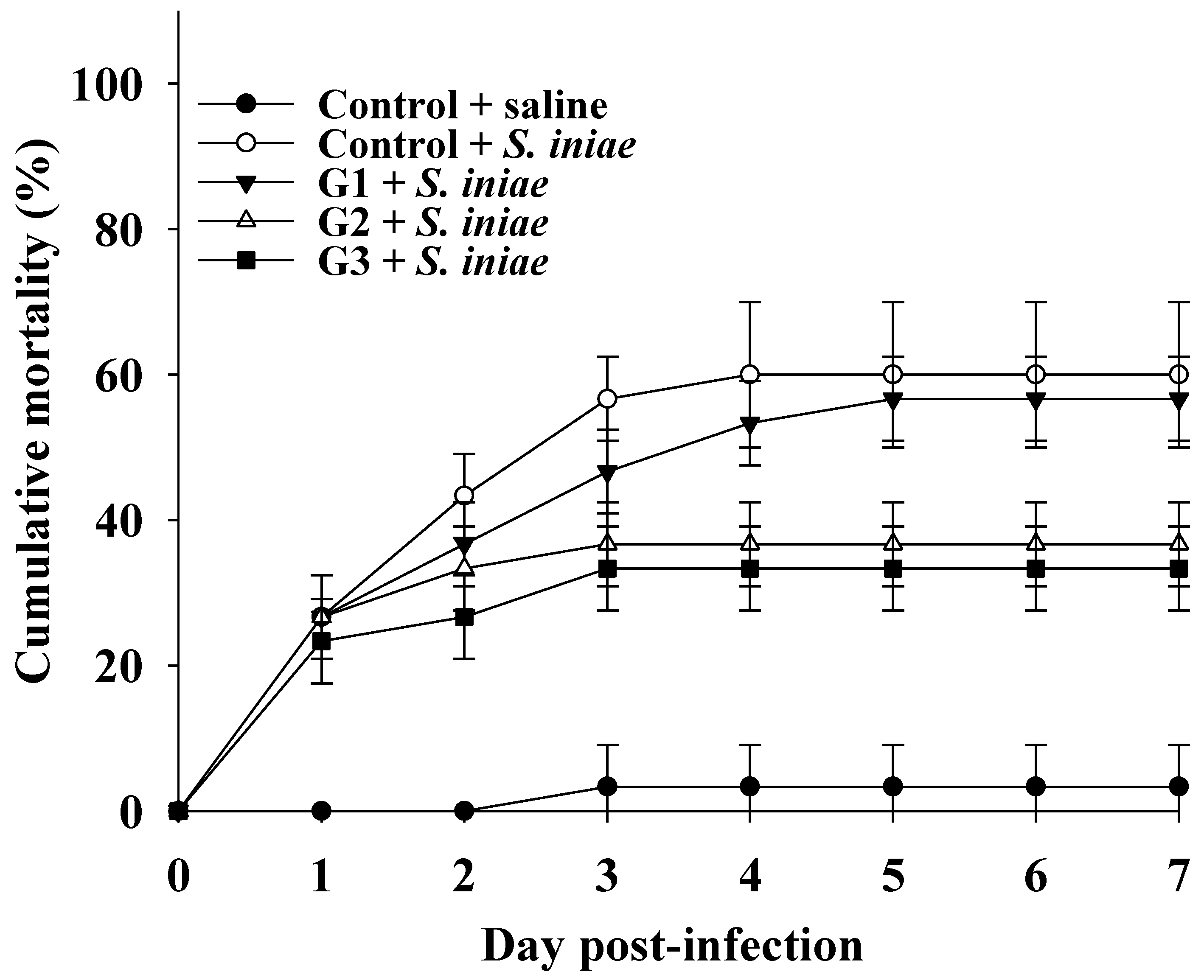
2.7. Challenge Test
2.8. Statistical Analysis
3. Results
3.1. COS Supplementation Enhances Growth Performance
3.2. COS Supplementation Enhances Digestive Enzyme Activity and Gene Expressions Involved in Nutrient Metabolism
3.3. COS Supplementation Enhances Innate Immunity
3.4. COS Supplementation Enhances Defense against S. Iniae Infection in Tilapia
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The state of world fisheries and aquaculture 2022. In Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar]
- Ortega, C.; Garcia, I.; Irgang, R.; Fajardo, R.; Tapia-Cammas, D.; Acosta, J.; Avendano-Herrera, R. First identification and characterization of Streptococcus iniae obtained from tilapia (Oreochromis aureus) farmed in Mexico. J. Fish Dis. 2018, 41, 773–782. [Google Scholar] [CrossRef] [PubMed]
- Rahmatullah, M.; Ariff, M.; Kahieshesfandiari, M.; Daud, H.M.; Zamri-Saad, M.; Sabri, M.Y.; Amal, M.N.A.; Ina-Salwany, M.Y. Isolation and pathogenicity of streptococcus iniae in cultured red hybrid tilapia in Malaysia. J. Aquat. Anim. Health 2017, 29, 208–213. [Google Scholar] [CrossRef]
- Bedu-Ferrari, C.; Biscarrat, P.; Langella, P.; Cherbuy, C. Prebiotics and the human gut microbiota: From breakdown mechanisms to the impact on metabolic health. Nutrients 2022, 14, 2096. [Google Scholar] [CrossRef]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion mechanisms mediated by probiotics and prebiotics and their potential impact on human health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef] [PubMed]
- Liaqat, F.; Eltem, R. Chitooligosaccharides and their biological activities: A comprehensive review. Carbohydr. Polym. 2018, 184, 243–259. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; Phil, L.; Sohail, M.; Hasnat, M.; Baig, M.; Ihsan, A.U.; Shumzaid, M.; Kakar, M.U.; Mehmood Khan, T.; Akabar, M.D.; et al. Chitosan oligosaccharide (COS): An overview. Int. J. Biol. Macromol. 2019, 129, 827–843. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.P. Research advances in chitooligosaccharides. Chin. J. Biochem. Pharm. 2003, 24, 210–212. [Google Scholar]
- Goncalves, C.; Ferreira, N.; Lourenco, L. Production of low molecular weight chitosan and chitooligosaccharides (cos): A review. Polymers 2021, 13, 2466. [Google Scholar] [CrossRef]
- de Oliveira, P.N.; Moussa, A.; Milhau, N.; Dosciatti Bini, R.; Prouillac, C.; Ferraz de Oliveira, B.; Dias, G.S.; Santos, I.A.; Morfin, I.; Sudre, G.; et al. In situ synthesis of Fe3O4 nanoparticles coated by chito-oligosaccharides: Physico-chemical characterizations and cytotoxicity evaluation for biomedical applications. Nanotechnology 2020, 31, 175602. [Google Scholar] [CrossRef]
- Yu, M.; Meng, T.; He, W.; Huang, H.; Liu, C.; Fu, X.; He, J.; Yin, Y.; Xiao, D. Dietary chito-oligosaccharides Improve intestinal immunity via regulating microbiota and Th17/Treg Balance-related immune signaling in piglets challenged by enterotoxigenic E. Coli. J. Agric. Food. Chem. 2021, 69, 15195–15207. [Google Scholar] [CrossRef]
- Zhang, J.; Cai, K.; Mishra, R.; Jha, R. In ovo supplementation of chitooligosaccharide and chlorella polysaccharide affects cecal microbial community, metabolic pathways, and fermentation metabolites in broiler chickens. Poult. Sci. 2020, 99, 4776–4785. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Yang, X.; Li, Q.; Ye, S.; Li, H. Enhanced immune response and resistance to edwardsiellosis following dietary chitooligosaccharide supplementation in the olive flounder (paralichthys olivaceus). Fish Shellfish Immunol. 2015, 47, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.D.; Van Dang, P.; Le, A.Q.; Nguyen, T.K.L.; Pham, D.H.; Van Nguyen, N.; Nguyen, Q.H. Effect of oligochitosan and oligo-beta-glucan supplementation on growth, innate immunity, and disease resistance of striped catfish (pangasianodon hypophthalmus). Biotechnol. Appl. Biochem. 2017, 64, 564–571. [Google Scholar] [CrossRef]
- Lin, S.; Mao, S.; Guan, Y.; Lin, X.; Luo, L. Dietary administration of chitooligosaccharides to enhance growth, innate immune response and disease resistance of trachinotus ovatus. Fish Shellfish Immunol. 2012, 32, 909–913. [Google Scholar] [CrossRef]
- Liu, L.; Zhou, Y.; Zhao, X.; Wang, H.; Wang, L.; Yuan, G.; Asim, M.; Wang, W.; Zeng, L.; Liu, X.; et al. Oligochitosan stimulated phagocytic activity of macrophages from blunt snout bream (megalobrama amblycephala) associated with respiratory burst coupled with nitric oxide production. Dev. Comp. Immunol. 2014, 47, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.W.; Liu, C.H.; Hu, S.Y. Dietary administration of probiotic paenibacillus ehimensis NPUST1 with bacteriocin-like activity improves growth performance and immunity against aeromonas hydrophila and streptococcus iniae in nile tilapia (oreochromis niloticus). Fish Shellfish Immunol. 2019, 84, 695–703. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analyses, 16th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1997. [Google Scholar]
- Tan, H.Y.; Chen, S.W.; Hu, S.Y. Improvements in the growth performance, immunity, disease resistance, and gut microbiota by the probiotic rummeliibacillus stabekisii in nile tilapia (oreochromis niloticus). Fish Shellfish Immunol. 2019, 92, 265–275. [Google Scholar] [CrossRef]
- Sankar, H.; Philip, B.; Philip, R.; Singh, I.S.B. Effect of probiotics on digestive enzyme activities and growth of cichlids, etroplus suratensis (pearl spot) and oreochromis mossambicus (tilapia). Aquac. Nutr. 2017, 23, 852–864. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Saputra, F.; Shiu, Y.L.; Chen, Y.C.; Puspitasari, A.W.; Danata, R.H.; Liu, C.H.; Hu, S.Y. Dietary supplementation with xylanase-expressing b.Amyloliquefaciens r8 improves growth performance and enhances immunity against aeromonas hydrophila in nile tilapia (oreochromis niloticus). Fish Shellfish Immunol. 2016, 58, 397–405. [Google Scholar] [CrossRef]
- Durak, I.; Yurtarslanl, Z.; Canbolat, O.; Akyol, O. A methodological approach to superoxide-dismutase (sod) activity assay based on inhibition of nitroblue tetrazolium (nbt) reduction. Clin. Chim. Acta 1993, 214, 103–104. [Google Scholar] [CrossRef]
- Ellis, A.E. Lysozyme assays. In Technigues in Fish Immunology; Stolen, J.S., Fletcher, T.C., Anderson, D.P., Roberson, B.S., Muiswinkel, W.B.v., Eds.; SOS Publications: Lagos, Nigeria, 1990. [Google Scholar]
- Shi, F.; Qiu, X.; Nie, L.; Hu, L.; Babu, V.S.; Lin, Q.; Zhang, Y.; Chen, L.; Li, J.; Lin, L.; et al. Effects of oligochitosan on the growth, immune responses and gut microbes of tilapia (oreochromis niloticus). Fish Shellfish Immunol. 2020, 106, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Su, P.; Han, Y.; Jiang, C.; Ma, Y.; Pan, J.; Liu, S.; Zhang, T. Effects of chitosan-oligosaccharides on growth performance, digestive enzyme and intestinal bacterial flora of tiger puffer (takifugu rubripes temminck et schlegel, 1850). J. Appl. Ichthyol. 2017, 33, 458–467. [Google Scholar] [CrossRef]
- Anguiano, M.; Pohlenz, C.; Buentello, A.; Gatlin, D.M., 3rd. The effects of prebiotics on the digestive enzymes and gut histomorphology of red drum (sciaenops ocellatus) and hybrid striped bass (morone chrysops x m. Saxatilis). Br. J. Nutr. 2013, 109, 623–629. [Google Scholar] [CrossRef]
- Magouz, F.I.; Bassuini, M.I.; Khalafalla, M.M.; Abbas, R.; Sewilam, H.; Aboelenin, S.M.; Soliman, M.M.; Amer, A.A.; Soliman, A.A.; Van Doan, H.; et al. Mannan oligosaccharide enhanced the growth rate, digestive enzyme activity, carcass composition, and blood chemistry of thinlip grey mullet (liza ramada). Animals 2021, 11, 3559. [Google Scholar] [CrossRef]
- Akter, M.N.; Sutriana, A.; Talpur, A.D.; Hashim, R. Dietary supplementation with mannan oligosaccharide influences growth, digestive enzymes, gut morphology, and microbiota in juvenile striped catfish, pangasianodon hypophthalmus. Aquac. Int. 2016, 24, 127–144. [Google Scholar] [CrossRef]
- Jeong, H.W.; Cho, S.Y.; Kim, S.; Shin, E.S.; Kim, J.M.; Song, M.J.; Park, P.J.; Sohn, J.H.; Park, H.; Seo, D.B.; et al. Chitooligosaccharide induces mitochondrial biogenesis and increases exercise endurance through the activation of sirt1 and ampk in rats. PLoS ONE 2012, 7, e40073. [Google Scholar] [CrossRef]
- Steele, N.C.; McMurtry, J.P.; Campbell, R.G.; Caperna, T.J.; Rosebrough, R.W. Effect of dietary energy intake and exogenous porcine growth hormone administration on circulating porcine growth hormone concentration and response to human growth hormone-releasing factor administration in growing swine. Domest. Anim. Endocrinol. 1995, 12, 293–298. [Google Scholar] [CrossRef]
- Tang, Z.R.; Yin, Y.L.; Nyachoti, C.M.; Huang, R.L.; Li, T.J.; Yang, C.; Yang, X.J.; Gong, J.; Peng, J.; Qi, D.S.; et al. Effect of dietary supplementation of chitosan and galacto-mannan-oligosaccharide on serum parameters and the insulin-like growth factor-I mRNA expression in early-weaned piglets. Domest. Anim. Endocrinol. 2005, 28, 430–441. [Google Scholar] [CrossRef]
- Ouyang, A.; Wang, H.; Su, J.; Liu, X. Mannose receptor mediates the activation of chitooligosaccharides on blunt snout bream (megalobrama amblycephala) macrophages. Front. Immunol. 2021, 12, 686846. [Google Scholar] [CrossRef]
- Ragland, S.A.; Criss, A.K. From bacterial killing to immune modulation: Recent insights into the functions of lysozyme. PLoS Pathog. 2017, 13, e1006512. [Google Scholar] [CrossRef] [PubMed]
- Biller, J.D.; Do Valle Polycarpo, G.; Moromizato, B.S.; Sidekerskis, A.P.D.; Da Silva, T.D.; Dos Reis, I.C.; Fierro-Castro, C. Lysozyme activity as an indicator of innate immunity of tilapia (oreochromis niloticus) when challenged with lps and streptococcus agalactiae. Rev. Bras. Zootec. 2021, 50, e20210053. [Google Scholar] [CrossRef]
- Wu, C.; Dai, Y.; Yuan, G.; Su, J.; Liu, X. Immunomodulatory effects and induction of apoptosis by different molecular weight chitosan oligosaccharides in head kidney macrophages from blunt snout bream (megalobrama amblycephala). Front. Immunol. 2019, 10, 869. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Administrations | |||
|---|---|---|---|---|
| Control | G1 | G2 | G3 | |
| COSs | 0 | 4 | 8 | 12 |
| Fish meal | 50 | 50 | 50 | 50 |
| Soybean meal | 300 | 300 | 300 | 300 |
| Wheat middling | 161 | 161 | 161 | 161 |
| Rice bran | 300 | 300 | 300 | 300 |
| α-starch | 60 | 60 | 60 | 60 |
| Cellulose | 70 | 66 | 62 | 58 |
| Skim milk | 10 | 10 | 10 | 10 |
| Soybean oil | 30 | 30 | 30 | 30 |
| Mineral mixture a | 16 | 16 | 16 | 16 |
| Vitamin mixture a | 3 | 3 | 3 | 3 |
| Proximate composition b | ||||
| Crude protein (%) | 26.8 | 27.3 | 27.4 | 27.5 |
| Crude lipid (%) | 12.1 | 12.2 | 12.6 | 12.7 |
| Moisture (%) | 10.4 | 10.4 | 10.6 | 10.5 |
| Ash (%) | 15.0 | 15.1 | 15.6 | 15.7 |
| Gene Name | Primer Sequences (5’→3’) | Amplicon Size | Accession No. |
|---|---|---|---|
| Glucose kinase (GK) | F: GCAGCGAGGAAGCCATGAAGA R: GAGGTCCCTGACGACTTTGTGG | 101 bp | XM_003451020 |
| Glucose-6-phosphatase (G6Pase) | F: AGCGCGAGCCTGAAGAAGTACT R: ATGGTCCACAGCAGGTCCACAT | 107 bp | XM_003448671 |
| Growth hormone receptor-1 (GHR-1) | F: GAATACAAGTCCTTCCGGGCTAA R: CTCATACTCCACACGCATCCA | 100 bp | AY973232 |
| Insulin-like growth factor-1 (IGF-1) | F: TGTCTGCCAGTAAGGATGTTCTTG R: GGCTTTCCACGCCACTTAAC | 100 bp | EU272149 |
| Tumor necrosis factor-α (TNF-α) | F:CCAGAAGCACTAAAGGCGAAGA R:CCTTGGCTTTGCTGCTGATC | 82 bp | AY428948 |
| Interleukin-1β (IL-1β) | F: TGTCGCTCTGGGCATCAA R: GGCTTGTCGTCATCCTTGTGA | 63 bp | KJ574402 |
| Interleukin-8 (IL-8) | F: CCTCGAGAAGGTGGATGTGAA R: CATGAGACCCAGGGCATCA | 100 bp | GQ355864 |
| β-actin | F: CCACACAGTGCCCATCTACGA R: CCACGCTCTGTCAGGATCTTCA | 111 bp | EU887951 |
| Parameters | Administrations (COS Supplementation Level g kg−1) | |||
|---|---|---|---|---|
| 0 (Control) | 4 (G1) | 8 (G2) | 12 (G3) | |
| Initial weight (g) | 1.04 ± 0.02 a | 1.04 ± 0.04 a | 1.04 ± 0.09 a | 1.04 ± 0.04 a |
| Finial weight (g) | 10.55 ± 0.9 d | 14.09 ± 0.97 c | 15.50 ± 0.51 b | 17.49 ± 0.37 a |
| Weight gain (WG) (g) | 9.50 ± 0.1 d | 13.86 ± 0.48 c | 15.23 ± 0.51 b | 16.45 ± 0.54 a |
| Feed efficiency (FE) | 0.66 ± 0.01 d | 0.72 ± 0.03 c | 0.76 ± 0.03 ab | 0.78 ± 0.04 a |
| Specific growth rate (SGR) | 1.51 ± 0.07 d | 2.09 ± 0.08 c | 2.31 ± 0.09 b | 2.51 ± 0.07 a |
| Survival rate (%) | 94.4 ± 1.9 a | 94.4 ± 1.9 a | 97.7 ± 3.8 a | 97.7 ± 3.8 a |
| Activity (U/mg) | Control | G1 | G2 | G3 |
|---|---|---|---|---|
| Protease | 0.23 ± 0.08 c | 0.36 ± 0.07 b | 0.39 ± 0.04 b | 0.54 ± 0.09 a |
| Amylase | 0.27 ± 0.06 c | 0.31 ± 0.04 bc | 0.35 ± 0.02 b | 0.49 ± 0.02 a |
| Cellulase | 0.23 ± 0.05 c | 0.35 ± 0.09 bc | 0.41 ± 0.07 ab | 0.58 ± 0.09 a |
| Xylanase | 0.17 ± 0.03 a | 0.16 ± 0.03 a | 0.21 ± 0.07 a | 0.18 ± 0.05 a |
| Lipase | 0.07 ± 0.008 b | 0.09 ± 0.01 b | 0.16 ± 0.04 a | 0.21 ± 0.07 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nurmalasari; Liu, C.-H.; Maftuch, I.M.; Hu, S.-Y. Dietary Supplementation with Prebiotic Chitooligosaccharides Enhances the Growth Performance, Innate Immunity and Disease Resistance of Nile Tilapia (Oreochromis niloticus). Fishes 2022, 7, 313. https://doi.org/10.3390/fishes7060313
Nurmalasari, Liu C-H, Maftuch IM, Hu S-Y. Dietary Supplementation with Prebiotic Chitooligosaccharides Enhances the Growth Performance, Innate Immunity and Disease Resistance of Nile Tilapia (Oreochromis niloticus). Fishes. 2022; 7(6):313. https://doi.org/10.3390/fishes7060313
Chicago/Turabian StyleNurmalasari, Chun-Hung Liu, Ir. M. Maftuch, and Shao-Yang Hu. 2022. "Dietary Supplementation with Prebiotic Chitooligosaccharides Enhances the Growth Performance, Innate Immunity and Disease Resistance of Nile Tilapia (Oreochromis niloticus)" Fishes 7, no. 6: 313. https://doi.org/10.3390/fishes7060313
APA StyleNurmalasari, Liu, C.-H., Maftuch, I. M., & Hu, S.-Y. (2022). Dietary Supplementation with Prebiotic Chitooligosaccharides Enhances the Growth Performance, Innate Immunity and Disease Resistance of Nile Tilapia (Oreochromis niloticus). Fishes, 7(6), 313. https://doi.org/10.3390/fishes7060313