
Factors Influencing Early Growth of Juvenile Tiger Trout Stocked into Subalpine Lakes as Biocontrol and to Enhance Recreational Angling
 ,
,
Abstract
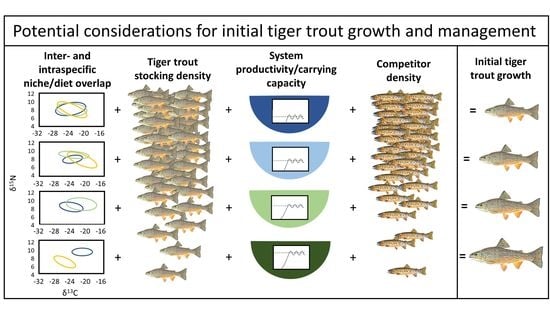
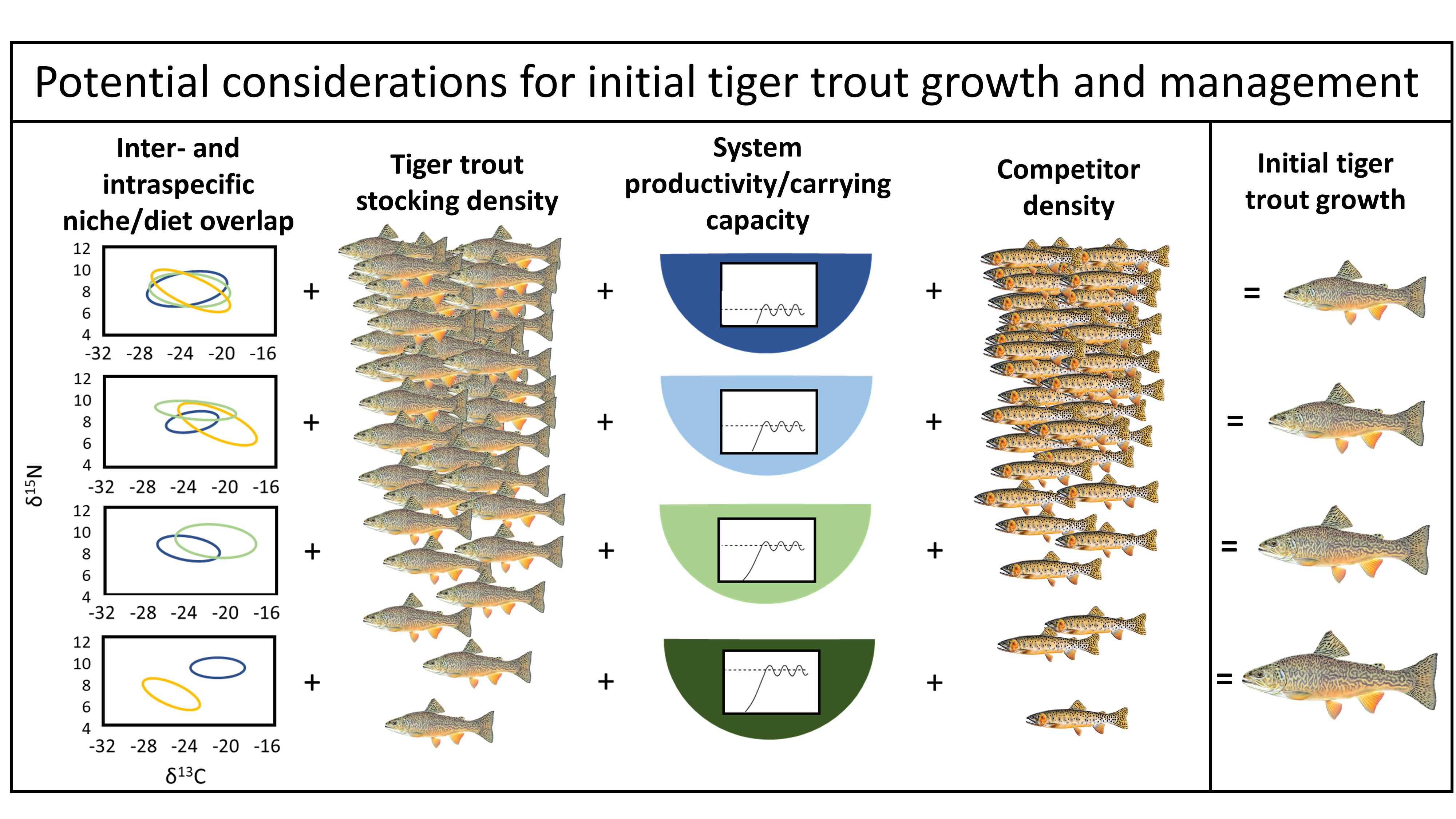
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Sampling
2.2.1. Physical Limnology, Zooplankton, and Invertebrate Collections
2.2.2. Fish Sampling and Collection of Biological Samples
2.3. Laboratory Analysis of Biological Samples
2.3.1. Stable Isotope Analysis
2.3.2. Age and Growth of Tiger Trout
2.4. Data Analysis
2.4.1. Stable Isotope Analysis of Trophic Niche Space and Overlap
2.4.2. Diet Characterization from Stable Isotopes of Carbon and Nitrogen
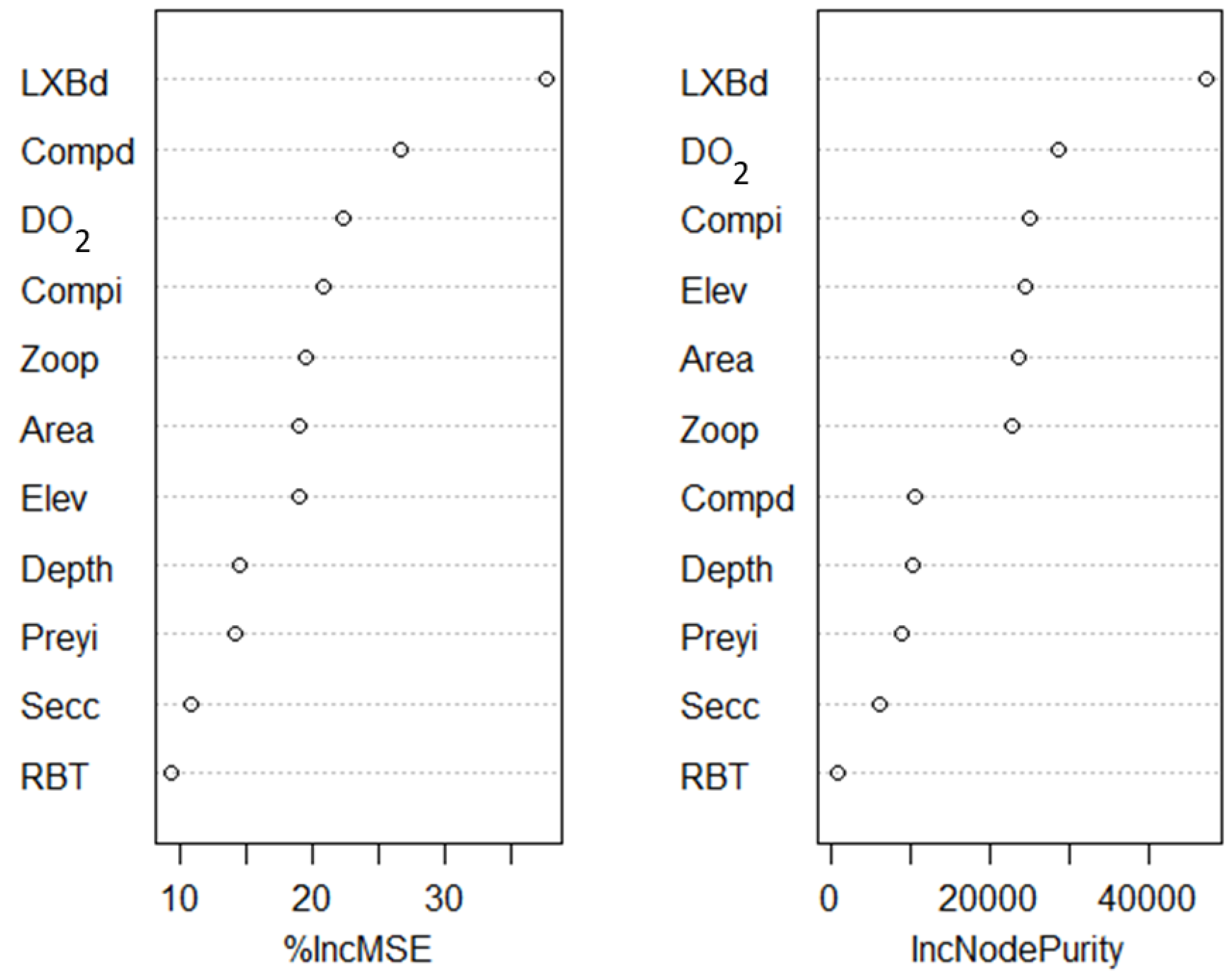
2.4.3. Predictors of Early Tiger Trout Growth and Random Forest Modeling
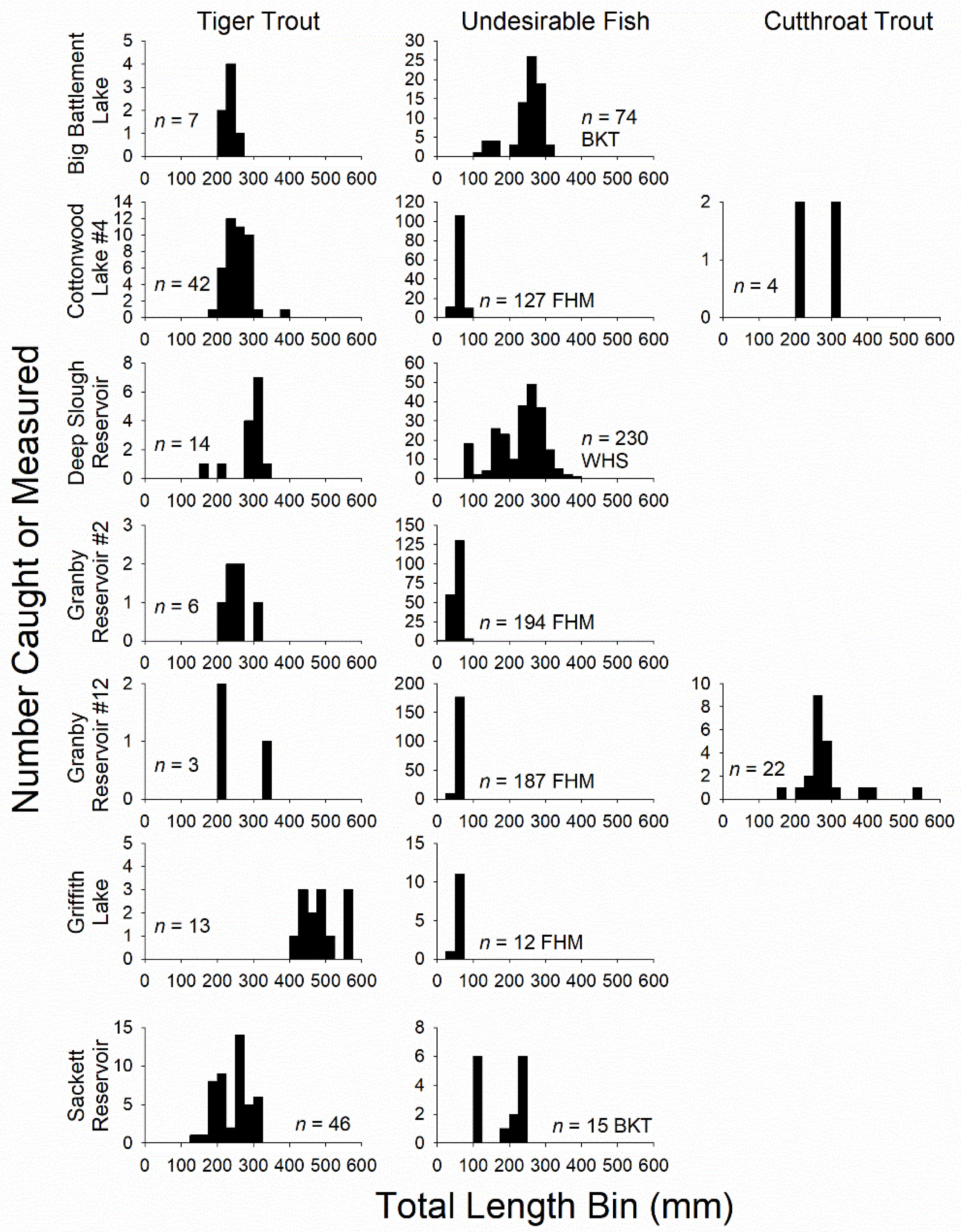
3. Results
3.1. Variation and Patterns in Tiger Trout Growth and System Attributes
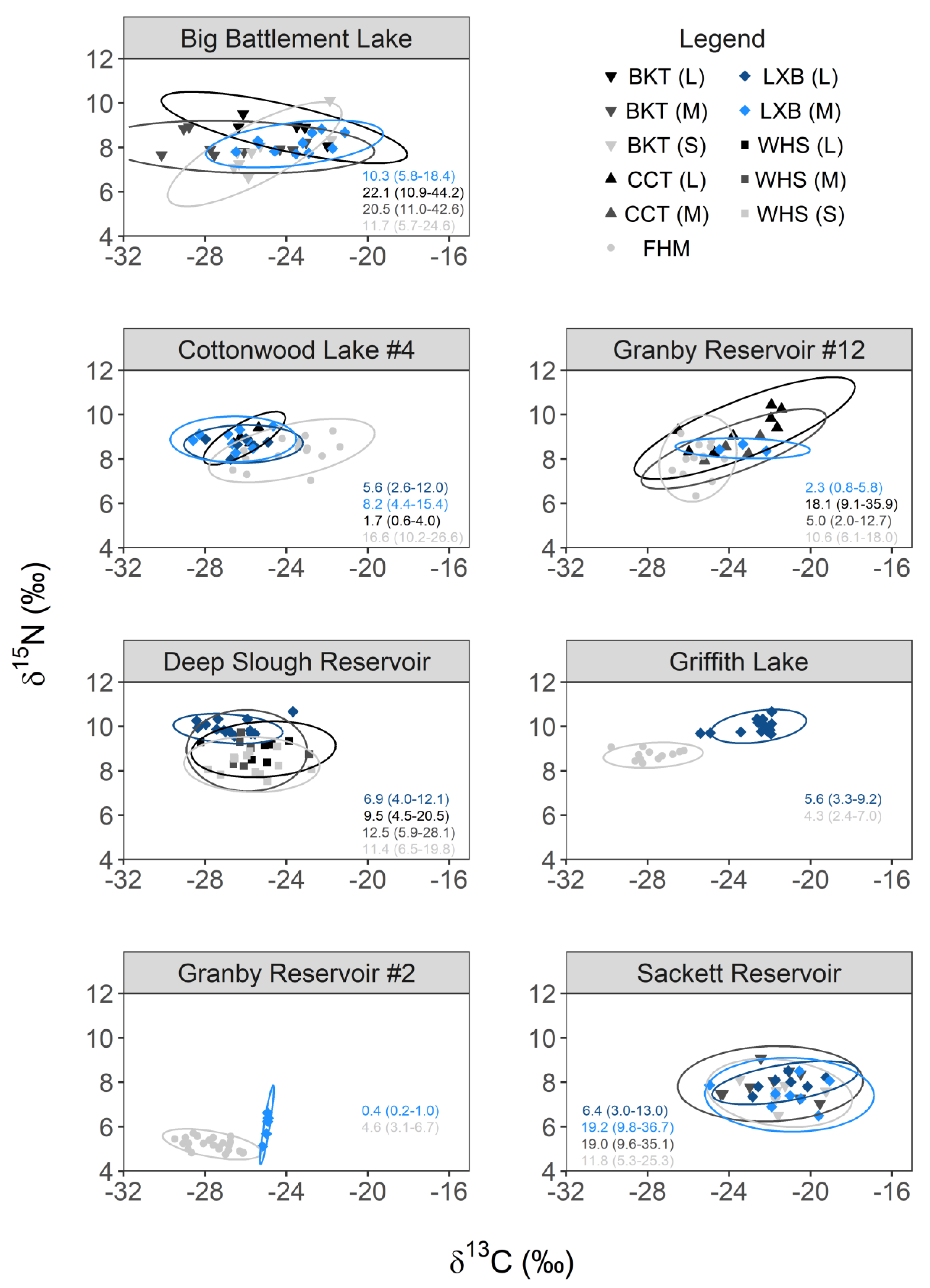
3.2. Trophic Niche Overlap and Potential for Competition
3.3. Rates of Piscivory among Size-Groups, Species and Study Systems
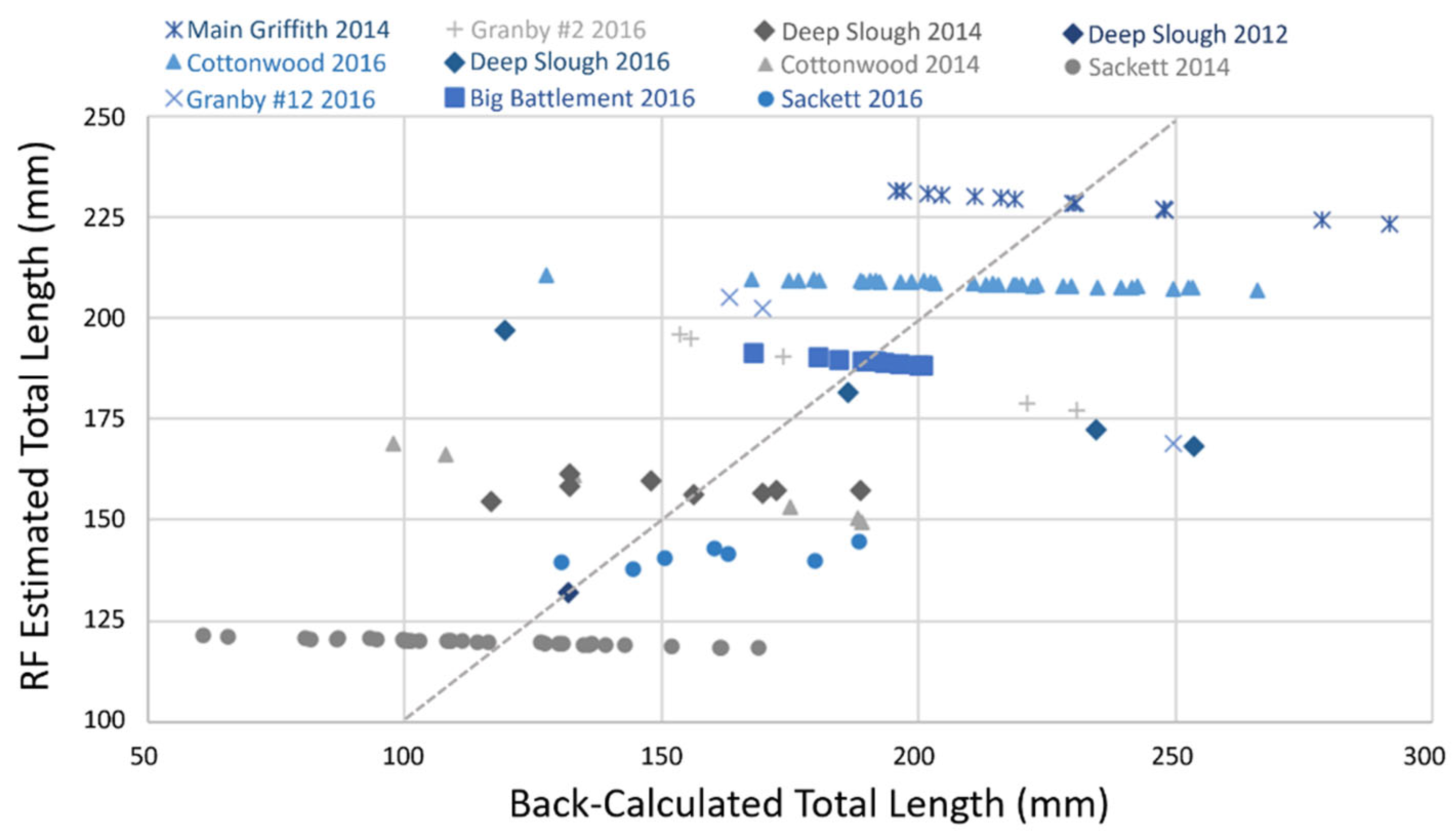
3.4. Predictors of Early Tiger Trout Growth and Random Forest Modeling
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pipalova, I.A. Review of grass carp use for aquatic weed control and its impact on water bodies. J. Aquat. Plant Manag. 2006, 44, 1–12. [Google Scholar]
- Bickerton, M.W.; Corleto, J.; Verna, T.N.; Williges, E.; Matadha, D. Comparative efficacy of Pimephales promelas, Fundulus diaphanous, and Gambusia affinis and influence of prey density for biological control of Culex pipiens molestus larvae. J. Am. Mosq. Control. Assoc. 2018, 34, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Koenig, M.K.; Meyer, K.A.; Kozfkay, J.R.; Dupont, J.M.; Schriever, E.B. Evaluating the ability of tiger muskellunge to eradicate brook trout in Idaho alpine lakes. N. Am. J. Fish. Manag. 2015, 35, 659–670. [Google Scholar] [CrossRef]
- Kerr, S.J.; Lasenby, T.A. Esocid Stocking: An Annotated Bibliography and Literature Review; Ontario Ministry of Natural Resources Fish and Wildlife Branch: Peterborough, ON, Canada, 2001.
- Irwin, B.J.; Devries, D.R.; Wright, R.A. Evaluating the potential for predatory control of gizzard shad by largemouth bass in small impoundments: A bioenergetics approach. Trans. Am. Fish. Soc. 2003, 132, 913–924. [Google Scholar] [CrossRef]
- Bunnell, D.B.; Madenjian, C.P.; Claramunt, R.M. Long-term changes of the Lake Michigan fish community following the reduction of exotic Alewife (Alosa pseudoharengus). Can. J. Fish. Aquat. Sci. 2006, 63, 2434–2446. [Google Scholar] [CrossRef]
- Lepak, J.M.; Cathcart, C.N.; Stacy, W.L. Tiger muskellunge predation on stocked salmonids intended for recreational fisheries. Lake Reserv. Manag. 2014, 30, 250–257. [Google Scholar] [CrossRef]
- Kozfkay, J.R.; Dillon, J.C.; Schill, D.J. Routine use of sterile fish in salmonid sport fisheries: Are we there yet? Fisheries 2006, 31, 392–401. [Google Scholar] [CrossRef]
- Farrell, C.J.; Johnson, B.M.; Hansen, A.G.; Myrick, C.A. Induced triploidy reduces mercury bioaccumulation in a piscivorous fish. Can. J. Fish. Aquat. Sci. 2022, 79, 200–212. [Google Scholar] [CrossRef]
- Martinez, P.J.; Bigelow, P.E.; Deleray, M.A.; Fredenberg, W.A.; Hansen, B.S.; Horner, N.J.; Lehr, S.K.; Schneidervin, R.W.; Tolentino, S.A.; Viola, A.E. Western lake trout woes. Fisheries 2009, 34, 424–442. [Google Scholar] [CrossRef]
- Winters, L.K.; Budy, P.; Thiede, G.P. Earning their stripes: The potential of tiger trout and other salmonids as biological controls of forage fishes in a western reservoir. N. Am. J. Fish. Manag. 2017, 37, 380–394. [Google Scholar] [CrossRef]
- Messner, J.; Schoby, G.; Belnap, M.; Amick, M.; Loffredo, J. Fisheries Management Annual Report, Salmon Region, IDFG 16–115; Idaho Department of Fish and Game: Salmon, ID, USA, 2017.
- Miller, A.L. Diet, Growth, and Age Analysis of Tiger Trout from Ten Lakes in Eastern Washington. Master’s Thesis, Eastern Washington University, Cheney, WA, USA, 2010. [Google Scholar]
- Winters, L.K.; Budy, P. Exploring crowded trophic niche space in a novel reservoir fish assemblage: How many is too many? Trans. Am. Fish. Soc. 2015, 144, 1117–1128. [Google Scholar] [CrossRef]
- Sogard, S.M. Size-selective mortality in the juvenile stage of teleost fishes: A review. Bull. Mar. Sci. 1997, 60, 1129–1157. [Google Scholar]
- Stige, L.C.; Rogers, L.A.; Neuheimer, A.B.; Hunsicker, M.E.; Yaragina, N.A.; Ottersen, G.; Ciannelli, L.; Langangen, Ø.; Durant, J.M. Density- and size-dependent mortality in fish early life stages. Fish Fish. 2019, 20, 962–976. [Google Scholar] [CrossRef]
- Olson, M.H. Ontogenetic niche shifts in largemouth bass: Variability and consequences for first-year growth. Ecology 1996, 77, 179–190. [Google Scholar] [CrossRef]
- Hanson, J.M.; Leggett, W.C. Experimental and field evidence for inter- and intraspecific competition in two freshwater fishes. Can. J. Fish. Aquat. Sci. 1985, 42, 280–286. [Google Scholar] [CrossRef]
- Nunn, A.D.; Vickers, L.H.; Mazik, K.; Bolland, J.D.; Peirson, G.; Axford, S.N.; Henshaw, A.; Cowx, I.G. Dynamic competition and resource partitioning during the early life of two widespread, abundant and ecologically similar fishes. Hydrobiologia 2020, 847, 2211–2224. [Google Scholar] [CrossRef]
- Kennedy, G.J.A.; Strange, C.D. The effects of intra- and inter-specific competition on the survival and growth of stocked juvenile Atlantic salmon, Salmo salar L., and resident trout, Salmo trutta L., in an upland stream. J. Fish. Biol. 1986, 28, 479–489. [Google Scholar] [CrossRef]
- Hansen, A.G.; Gardner, J.A.; Connelly, K.A.; Polacek, M.; Beauchamp, D.A. Resource use among top-level piscivores in a temperate reservoir: Implications for a threatened coldwater Specialist. Ecol. Freshw. Fish 2022, 31, 469–491. [Google Scholar] [CrossRef]
- Cutler, D.R.; Edwards, T.C., Jr.; Beard, K.J.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random forests for classification in ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef]
- California Department of Fish and Wildlife. High Mountain Lakes Project. Available online: https://www.wildlife.ca.gov/Regions/6/High-Mountain-Lakes-Project (accessed on 31 October 2022).
- Hansen, A.G.; Gardner, J.R.; Beauchamp, D.A.; Paradis, R.; Quinn, T.P. Recovery of sockeye salmon in the Elwha River, Washington, after dam removal: Dependence of smolt production on the resumption of anadromy by landlocked kokanee. Trans. Am. Fish. Soc. 2016, 145, 1303–1317. [Google Scholar] [CrossRef]
- Watkins, J.; Rudstam, L.; Holeck, K. Length-Weight Regressions for Zooplankton Biomass Calculations—A Review and a Suggestion for Standard Equations; Cornell Biological Field Station, Cornell University: Ithaca, NY, USA, 2011. [Google Scholar]
- Beauchamp, D.A.; Parrish, D.L.; Whaley, R.A. Coldwater fish in large standing waters. In Standard Methods for Sampling North American Freshwater Fishes; Bonar, S.A., Hubert, W.A., Willis, D.W., Eds.; American Fisheries Society: Bethesda, MD, USA, 2009; pp. 97–118. [Google Scholar]
- Sanderson, B.L.; Tran, C.D.; Coe, H.J.; Pelekis, V.; Steel, E.A.; Reichert, W.L. Nonlethal sampling of fish caudal fins yields valuable stable isotope data for threatened and endangered fishes. Trans. Am. Fish. Soc. 2009, 138, 1166–1177. [Google Scholar] [CrossRef]
- Post, D.M.; Layman, C.A.; Arrington, D.A.; Takimoto, G.; Quattrochi, J.; Montana, C.G. Getting to the fat of the matter: Models, methods, and assumptions for dealing with lipids in stable isotope analyses. Oecologia 2007, 152, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Quist, M.C.; Pegg, M.A.; DeVries, D.R. Age and growth. In Fisheries Techniques, 3rd ed.; Zale, A.V., Parrish, D.L., Sutton, T.M., Eds.; American Fisheries Society: Bethesda, MD, USA, 2013; pp. 677–721. [Google Scholar]
- Hansen, A.G.; Gardner, J.R.; Connelly, K.A.; Polacek, M.; Beauchamp, D.A. Trophic compression of lake food webs under hydrologic disturbance. Ecosphere 2018, 9, e02304. [Google Scholar] [CrossRef]
- Herwig, B.R.; Zimmer, K.D.; Staples, D.F. Using stable isotope data to quantify niche overlap and diets of muskellunge, northern pike and walleye in a deep Minnesota lake. Ecol. Freshw. Fish 2022, 31, 60–71. [Google Scholar] [CrossRef]
- Lysy, M.; Stasko, A.D.; Swanson, H.K. nicheROVER: Niche Region and Niche Overlap Metrics for Multidimensional Ecological Niches. Version 1.1.0. Available online: https://CRAN.R-project.org/package=nicheROVER (accessed on 24 June 2022).
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 24 June 2022).
- Swanson, H.K.; Lysy, M.; Power, M.; Stasko, A.D.; Johnson, J.D.; Reist, J.D. A new probabilistic method for quantifying n-dimensional ecological niches and niche overlap. Ecology 2015, 96, 318–324. [Google Scholar] [CrossRef]
- Parnell, A.C. simmr: A Stable Isotope Mixing Model. Version 0.4.5. Available online: https://CRAN.R-project.org/package=simmr (accessed on 24 June 2022).
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Hansen, A.G.; Beauchamp, D.A.; Baldwin, C.M. Environmental constraints on piscivory: Insights from linking ultrasonic telemetry to a visual foraging model for cutthroat trout. Trans. Am. Fish. Soc. 2013, 142, 300–316. [Google Scholar] [CrossRef]
- Murdoch, A.; Power, M. The effect of lake morphometry on thermal habitat use and growth in Arctic charr populations: Implications for understanding climate-change impacts. Ecol. Freshw. Fish 2013, 22, 453–466. [Google Scholar] [CrossRef]
- Estlander, S.; Nurminen, L.; Mrkvicka, T.; Olin, M.; Rask, M.; Lehtonen, H. Sex-dependent responses of perch to changes in water clarity and temperature. Ecol. Freshw. Fish 2015, 24, 544–552. [Google Scholar] [CrossRef]
- Church, K.D.W.; Grant, J.W.A. Does increasing habitat complexity favour particular personality types of juvenile Atlantic salmon, Salmo salar? Anim. Behav. 2018, 135, 165–176. [Google Scholar] [CrossRef]
- Andersen, M.R.; Kragh, T.; Sand-Jensen, K. Extreme diel dissolved oxygen and carbon cycles in shallow vegetated lakes. Proc. R. Soc. B. 2017, 284, 20171427. [Google Scholar] [CrossRef] [PubMed]
- Pennak, R.W. A Limnological Reconnaissance of Grand Mesa, Colorado; University of Colorado: Boulder, CO, USA, 1950. [Google Scholar]
- Grossman, G.D.; Simon, T.N. Density-dependent effects on salmonid populations: A review. Ecol. Freshw. Fish 2020, 29, 400–418. [Google Scholar] [CrossRef]
- Jeppesen, E.; Jensen, J.P.; Sondergaard, M.; Lauridsen, T.L.; Landkildehus, F. Trophic structure, species richness and biodiversity in Danish lakes: Changes along a phosphorus gradient. Freshw. Biol. 2000, 45, 201–218. [Google Scholar] [CrossRef]
- Jeppesen, E.; Noges, P.; Davidson, T.A.; Haberman, J.; Noges, T.; Blank, K.; Lauridsen, T.L.; Sondergaard, M.; Sayer, C.; Laugaste, R.; et al. Zooplankton as indicators in lakes: A scientific-based plea for including zooplankton in the ecological quality assessment of lakes according to the European Water Framework Directive (WFD). Hydrobiologia 2011, 676, 279. [Google Scholar] [CrossRef]
- Sorel, M.H.; Hansen, A.G.; Connelly, K.A.; Beauchamp, D.A. Trophic feasibility of reintroducing anadromous salmonids in three reservoirs on the North Fork Lewis River, Washington: Prey supply and consumption demand of resident fishes. Trans. Am. Fish. Soc. 2016, 145, 1331–1347. [Google Scholar] [CrossRef]
- Liaw, A.; Wiener, M. Classification and regression by random forest. R News 2002, 2, 18–22. [Google Scholar]
- Gini, C. Variabilità e Mutabilità; Tipografia di Paolo Cuppini: Bologna, Italy, 1912. [Google Scholar]
- Amundsen, P.; Knudsen, R.; Klemetsen, A. Intraspecific competition and density dependence of food consumption and growth in Arctic charr. J. Anim. Ecol. 2007, 767, 149–158. [Google Scholar] [CrossRef]
- Matte, J.-M.; Fraser, D.J.; Grant, J.W.A. Population variation in density-dependent growth, mortality and their trade-off in a stream fish. J. Anim. Ecol. 2020, 89, 541–552. [Google Scholar] [CrossRef]
- McIntyre, J.K.; Beauchamp, D.A.; Mazur, M.M.; Overman, N.C. Ontogenetic trophic interactions and benthopelagic coupling in Lake Washington: Evidence from stable isotopes and diet analysis. Trans. Am. Fish. Soc. 2006, 135, 1312–1328. [Google Scholar] [CrossRef]
- Ward, A.J.; Webster, M.M.; Hart, P.J.B. Intraspecific food competition in fishes. Fish Fish. 2006, 7, 231–261. [Google Scholar] [CrossRef]
- Prati, S.; Henriksen, E.H.; Smalås, A.; Knudsen, R.; Klemetsen, A.; Sánchez-Hernández, J.; Amundsen, P. The effect of inter- and intraspecific competition on individual and population niche widths: A four-decade study on two interacting salmonids. Oikos 2021, 130, 1679–1691. [Google Scholar] [CrossRef]
- Raborn, S.W.; Miranda, L.E.; Driscoll, M.T. Diet overlap and consumption patterns suggest seasonal flux in the likelihood for exploitative competition among piscivorous fishes. Ecol. Freshw. Fish 2004, 13, 276–284. [Google Scholar] [CrossRef]
- Sanchez-Hernandez, J.; Nunn, A.D.; Adams, C.E.; Amundsen, P.-A. Causes and consequences of ontogenetic dietary shifts: A global synthesis using fish models. Biol. Rev. 2019, 94, 539–554. [Google Scholar] [CrossRef] [PubMed]
- Marrin, D.L.; Erman, D.C. Evidence against competition between trout and nongame fishes in Stampede Reservoir, California. N. Am. J. Fish. Manag. 1982, 2, 262–269. [Google Scholar] [CrossRef]
- Tremblay, S.; Magnan, P. Interactions between two distantly related species, brook trout (Salvelinus fontinalis) and white sucker (Catostomus commersoni). Can. J. Fish. Aquat. Sci. 1991, 48, 857–867. [Google Scholar] [CrossRef]
- Teuscher, D.; Luecke, C. Competition between kokanees and Utah chub in Flaming Gorge Reservoir, Utah–Wyoming. Trans. Am. Fish. Soc. 1996, 125, 505–511. [Google Scholar] [CrossRef]
- Lachance, S.; Magnan, P. Performance of domestic, hybrid, and wild strains of brook trout, Salvelinus fontinalis, after stocking: The impact of intra- and interspecific competition. Can. J. Fish. Aquat. Sci. 1990, 47, 2278–2284. [Google Scholar] [CrossRef]
- Brodeur, P.; Magnan, P.; Legault, M. Response of fish communities to different levels of white sucker (Catostomus commersoni) biomanipulation in five temperate lakes. Can. J. Fish. Aquat. Sci. 2001, 58, 1998–2010. [Google Scholar] [CrossRef]
- Sanchez-Hernandez, J.; Eloranta, A.P.; Finstad, A.G.; Amundsen, P.-A. Community structure affects trophic ontogeny in a predatory fish. Ecol. Evol. 2017, 7, 358–367. [Google Scholar] [CrossRef]
- Rieman, B.E.; Myers, D.L. Influence of fish density and relative productivity on growth of kokanee in ten oligotrophic lakes and reservoirs in Idaho. Trans. Am. Fish. Soc. 1992, 121, 178–191. [Google Scholar] [CrossRef]
- Arranz, I.; Mehner, T.; Benejam, L.; Argillier, C.; Holmgren, K.; Jeppesen, E.; Lauridsen, T.L.; Volta, P.; Winfield, I.J.; Brucet, S. Density-dependent effects as key drivers of intraspecific size structure of six abundant fish species in lakes across Europe. Can. J. Fish. Aquat. Sci. 2015, 73, 519–534. [Google Scholar] [CrossRef]
- Persson, L.; Diehl, S.; Johansson, L.; Andersson, G.; Harmin, S.F. Shifts in fish communities along the productivity gradient of temperate lakes—Patterns and the importance of size-structured interactions. J. Fish. Biol. 1991, 38, 281–293. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Cole, J.J.; Hodgson, J.R.; Kitchell, J.F.; Pace, M.L.; Bade, D.; Cottingham, K.L.; Essington, T.E.; Houser, J.N.; Schindler, D.E. Trophic cascades, nutrients, and lake productivity: Whole-lake experiments. Ecol. Monogr. 2001, 71, 163–186. [Google Scholar] [CrossRef]
- Carlson, R.E. A trophic state index for lakes. Limnol. Oceanogr. 1977, 22, 361–369. [Google Scholar] [CrossRef]
- Richardson, W.B.; Zigler, S.J.; Dewey, M.R. Bioenergetic relations in submerged aquatic vegetation: An experimental test of prey use by juvenile bluegills. Ecol. Freshw. Fish 1998, 7, 1–12. [Google Scholar] [CrossRef]
- Savino, J.F.; Marschall, E.A.; Stein, R.A. Bluegill growth as modified by plant density: An exploration of underlying mechanisms. Oecologia 1992, 89, 153–160. [Google Scholar] [CrossRef]
- Waldrop, T.; Summerfelt, S.; Mazik, P.; Kenney, P.B.; Good, C. The effects of swimming exercise and dissolved oxygen on growth performance, fin condition and survival of rainbow trout Oncorhynchus mykiss. Aquac. Res. 2020, 51, 2582–2589. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System | Surface Area (ha) | Max. Depth (m) | Mean SD (m) | Elev. (m) | %DO ≤ 2 (mg/L) | Zooplankton Biomass Density (g Dry Weight/m2) |
|---|---|---|---|---|---|---|
| Big Battlement Lake | 16.6 | 13.7 | 4.88 | 3072 | 14% | 1.53 ± 2.12 (3) |
| Cottonwood Lake #4 | 15.3 | 10.7 | 3.46 | 3110 | 40% | 0.47 ± 0.10 (3) |
| Deep Slough Reservoir | 18.2 | 4.6 | 1.25 | 3053 | 20% | 1.47 ± 2.42 (3) |
| Granby Reservoir #2 | 5.4 | 6.7 | 2.88 | 3066 | 29% | 0.07 ± 0.09 (3) |
| Granby Reservoir #12 | 17.1 | 7.0 | 2.71 | 3048 | 5% | 0.50 ± 0.61 (3) |
| Griffith Lake | 25.3 | 9.4 | 3.63 | 3063 | 61% | 0.10 ± 0.02 (3) |
| Sackett Reservoir | 2.3 | 7.3 | 3.75 | 3188 | 0% | 0.03 ± 0.03 (3) |
| Year Stocked | Tiger Trout | Cutthroat Trout | Rainbow Trout |
|---|---|---|---|
| Big Battlement Lake | |||
| 2016 | 29 (476 @ 87.4) | ||
| Cottonwood Lake #4 | |||
| 2010 | 393 (6000 @ 245.6) | ||
| 2011 | 98(1500 @ 256.8) | ||
| 2012 | 158 (2416 @ 241.8) | ||
| 2013 | 721 (11000 @ 139.2) | 361 (5499 @ 249.2) | |
| 2014 | 138 (2100 @ 108.2) | 656 (10000 @ 127.5) | 197 (3000 @ 261.1) |
| 2015 | 196 (2985 @ 250.7) | ||
| 2016 | 207 (3153 @ 93.7) | 194 (2960 @ 257.3) | |
| Deep Slough Reservoir | |||
| 2011 | 55 (1000 @ 89.2) | ||
| 2012 | 132 (2400 @ 73.2) | ||
| 2014 | 164 (3000 @ 95.5) | ||
| 2016 | 83 (1505 @ 87.4) | ||
| Granby Reservoir #2 | |||
| 2011 | 65 (350 @ 23.6) | ||
| 2013 | 279 (1500 @ 34.5) | ||
| 2015 | 279 (1500 @ 34.5) | ||
| 2016 | 75 (404 @ 87.4) | ||
| Granby Reservoir #12 | |||
| 2011 | 292 (5000 @ 23.6) | ||
| 2013 | 292 (5000 @ 23.6) | ||
| 2015 | 292 (5000 @ 23.6) | ||
| 2016 | 14 (238 @ 87.4) | ||
| Griffith Lake | |||
| 2014 | 39 (1000 @ 95.5) | ||
| 2015 | 39 (1000 @ 95.5) | ||
| Sackett Reservoir | |||
| 2011 | 650 (1500 @ 109.0) | 633 (1455 @ 202.9) | |
| 2013 | 434 (1000 @ 255.5) | ||
| 2014 | 867 (2000 @ 95.5) | ||
| 2016 | 131 (303 @ 87.4) | ||
| Species | Experimental Gill Nets | Micro-Mesh Gill Nets | Miniature Trap Nets | |||
|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | |
| Big Battlement Lake 1 | ||||||
| Tiger trout | 0.36 (2) | 0.55 | 0.98 (5) | 1.18 | 0.00 | 0.00 |
| Brook trout | 9.29 (49) | 5.29 | 5.00 (25) | 3.47 | 0.00 | 0.00 |
| Cottonwood Lake #4 | ||||||
| Tiger trout | 6.03 (30) | 4.62 | 1.32 (12) | 1.16 | 0.00 | 0.00 |
| Cutthroat trout | 0.62 (3) | 0.42 | 0.11 (1) | 0.27 | 0.00 | 0.00 |
| Fathead minnow | 0.00 | 0.00 | 0.17 (1) | 0.41 | 3.68 (34) | 3.92 |
| Deep Slough Reservoir | ||||||
| Tiger trout | 3.42 (14) | 2.76 | Not fished 2 | 0.00 | 0.00 | |
| White sucker | 44.25 (183) | 22.50 | 2.75 (22) | 3.89 | ||
| Granby Reservoir #2 | ||||||
| Tiger trout | 0.45 (2) | 1.11 | 0.75 (4) | 1.85 | Not fished 2 | |
| Cutthroat trout | 0.00 | 0.00 | 0.19 (1) | 0.46 | ||
| Fathead minnow | 0.00 | 0.00 | 13.69 (78) | 19.92 | ||
| Granby Reservoir #12 | ||||||
| Tiger trout | 0.22 (3) | 0.47 | 0.00 | 0.00 | 0.00 | 0.00 |
| Cutthroat trout | 1.88 (19) | 2.50 | 1.00 (3) | 1.22 | 0.00 | 0.00 |
| Fathead minnow | 0.00 | 0.00 | 91.03 (435) | 101.97 | 7.92 (44) | 6.08 |
| Griffith Lake | ||||||
| Tiger trout | 2.21 (11) | 1.97 | 0.29 (2) | 0.72 | 0.00 | 0.00 |
| Fathead minnow | 0.00 | 0.00 | 1.98 (10) | 3.3 | 0.23 (2) | 0.00 |
| Sackett Reservoir | ||||||
| Tiger trout | 3.52 (20) | 2.80 | 4.14 (26) | 0.30 | 0.00 | 0.00 |
| Brook trout | 0.98 (6) | 1.13 | 1.47 (9) | 1.30 | 0.00 | 0.00 |
| Lake and Tiger Trout Size-Group | Species and Size-Group of Potential Competitor | |||
|---|---|---|---|---|
| Big Battlement Lake: | BKT (S) | BKT (M) | BKT (L) | |
| Medium + Large | 45 (21–78) | 88 (61–100) | 83 (42–100) | |
| Cottonwood Lake #4: | FHM (S) | CCT (M+L) | LXB (M) | LXB (L) |
| Medium | 34 (4–81) | 30 (11–58) | 66 (35–95) | |
| Large | 59 (13–98) | 33 (9–69) | 84 (46–100) | |
| Deep Slough Reservoir: | WHS (S) | WHS (M) | WHS (L) | |
| Medium + Large | 4 (0–29) | 43 (2–95) | 29 (1–89) | |
| Granby Reservoir #2: | FHM (S) | |||
| Medium + Large | 2 (0–16) | |||
| Granby Reservoir #12: | FHM (S) | CCT (M) | CCT (L) | |
| Medium | 31 (1–81) | 71 (27–100) | 68 (9–100) | |
| Griffith Lake: | FHM (S) | |||
| Large | 0.2 (0–2) | |||
| Sackett Reservoir: | BKT (S) | BKT (M) | LXB (M) | LXB (L) |
| Small + Medium | 69 (37–95) | 79 (46–99) | 41 (17–72) | |
| Large | 78 (40–100) | 94 (71–100) | 91 (61–100) | |
| Species and Size-Group | Sample Size | Fish | Aquatic Invertebrates | Terrestrial Invertebrates | Zooplankton | Tiger Salamander |
|---|---|---|---|---|---|---|
| Big Battlement Lake | ||||||
| Brook trout (S) | 7 | 38.0 (9.4–63.5) | 35.2 (5.5–77.9) | 26.8 (5.0–52.2) | ||
| Brook trout (M) | 8 | 11.0 (1.8–27.0) | 40.1 (16.6–62.5) | 22.7 (3.1–55.5) | 26.3 (4.9–51.5) | |
| Brook trout (L) | 7 | 17.1 (2.4–39.9) | 30.0 (7.2–51.4) | 33.6 (4.9–73.7) | 19.3 (3.2–42.2) | |
| Tiger trout (M) | 10 | 12.6 (2.0–30.7) | 45.3 (30.0–61.0) | 30.1 (5.2–56.6) | 11.9 (2.1–27.8) | |
| Tiger trout (L) | 1 | 19.5 (2.4–56.4) | 41.1 (11.0–74.9) | 24.1 (2.4–66.4) | 15.3 (2.1–42.7) | |
| Cottonwood Lake #4 | ||||||
| Cutthroat trout (M) | 2 | 23.3 (2.7–60.3) | 25.7 (2.7–68.5) | 17.2 (2.3–42.7) | 33.9 (3.4–75.2) | |
| Cutthroat trout (L) | 2 | 26.0 (3.0–63.5) | 25.7 (2.8–69.3) | 15.5 (2.2–40.0) | 32.8 (3.6–72.5) | |
| Tiger trout (M) | 7 | 16.3 (2.5–36.0) | 20.7 (2.8–51.6) | 8.9 (1.9–19.6) | 54.1 (21.3–78.1) | |
| Tiger trout (L) | 8 | 16.1 (3.4–31.9) | 20.2 (3.2–47.5) | 13.3 (4.6–23.2) | 50.5 (26.8–69.4) | |
| Rainbow trout (L) | 2 | 23.0 (2.3–61.4) | 25.9 (2.4–70.4) | 16.5 (2.1–44.6) | 34.6 (3.9–79.3) | |
| Deep Slough Reservoir | ||||||
| Tiger trout (M) | 2 | 24.6 (2.5–67.3) | 28.2 (2.9–69.8) | 16.7 (2.1–47.2) | 30.5 (3.1–73.9) | |
| Tiger trout (L) | 12 | 32.3 (2.3–61.4) | 35.4 (16.3–52.9) | 10.6 (1.9–23.9) | 21.7 (5.4–43.4) | |
| Granby Reservoir #2 | ||||||
| Tiger trout (M) | 3 | 17.5 (1.9–51.5) | 20.7 (2.1–64.7) | 25.2 (2.4–61.5) | 13.6 (1.8–37.8) | 23.0 (2.0–65.3) |
| Tiger trout (L) | 2 | 18.1 (1.6–5.4) | 22.4 (2.1–62.8) | 22.9 (2.2–63.9) | 15.1 (1.9–45.8) | 21.6 (2.1–63.5) |
| Granby Reservoir #12 | ||||||
| Cutthroat trout (M) | 2 | 21.9 (2.4–62.6) | 25.8 (2.9–68.9) | 30.1 (2.9–76.4) | 22.2 (2.6–60.9) | |
| Cutthroat trout (L) | 10 | 25.6 (6.9–44.5) | 25.8 (3.5–54.1) | 33.8 (4.3–69.7) | 14.8 (2.6–32.7) | |
| Tiger trout (M) | 2 | 22.0 (2.5–62.0) | 24.8 (3.0–65.0) | 31.8 (3.2–75.0) | 21.3 (2.6–55.5) | |
| Tiger trout (L) | 1 | 15.1 (1.9–41.0) | 36.9 (4.3–75.8) | 33.3 (3.2–79.4) | 14.7 (1.9–42.2) | |
| Griffith Lake | ||||||
| Tiger trout (L) | 13 | 5.9 (0.8–16.4) | 17.2 (1.5–43.9) | 62.9 (31.7–88.6) | 4.3 (0.7–12.0) | 9.8 (1.3–24.2) |
| Sackett Reservoir | ||||||
| Brook trout (S) | 6 | 66.3 (50.4–80.7) | 20.7 (5.0–40.0) | 13.0 (2.6–26.8) | ||
| Brook trout (M) | 9 | 14.6 (2.3–32.3) | 58.3 (47.1–69.6) | 14.9 (2.3–32.8) | 12.2 (2.7–23.9) | |
| Tiger trout (S) | 1 | 59.6 (16.4–92.0) | 31.8 (2.9–74.6) | 8.6 (1.5–20.6) | ||
| Tiger trout (M) | 6 | 10.6 (1.6–26.4) | 68.4 (50.6–82.6) | 10.5 (1.7–27.1) | 10.5 (1.8–23.7) | |
| Tiger trout (L) | 8 | 18.0 (2.9–37.1) | 55.8 (45.0–66.5) | 17.5 (2.9–37.6) | 8.7 (1.7–18.9) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hansen, A.G.; Cristan, E.T.; Moll, M.M.; Miller, M.W.; Gardunio, E.I.; Lepak, J.M. Factors Influencing Early Growth of Juvenile Tiger Trout Stocked into Subalpine Lakes as Biocontrol and to Enhance Recreational Angling. Fishes 2022, 7, 342. https://doi.org/10.3390/fishes7060342
Hansen AG, Cristan ET, Moll MM, Miller MW, Gardunio EI, Lepak JM. Factors Influencing Early Growth of Juvenile Tiger Trout Stocked into Subalpine Lakes as Biocontrol and to Enhance Recreational Angling. Fishes. 2022; 7(6):342. https://doi.org/10.3390/fishes7060342
Chicago/Turabian StyleHansen, Adam G., Erik T. Cristan, Miranda M. Moll, Michael W. Miller, Eric I. Gardunio, and Jesse M. Lepak. 2022. "Factors Influencing Early Growth of Juvenile Tiger Trout Stocked into Subalpine Lakes as Biocontrol and to Enhance Recreational Angling" Fishes 7, no. 6: 342. https://doi.org/10.3390/fishes7060342
APA StyleHansen, A. G., Cristan, E. T., Moll, M. M., Miller, M. W., Gardunio, E. I., & Lepak, J. M. (2022). Factors Influencing Early Growth of Juvenile Tiger Trout Stocked into Subalpine Lakes as Biocontrol and to Enhance Recreational Angling. Fishes, 7(6), 342. https://doi.org/10.3390/fishes7060342