
Hematogenesis Adaptation to Long-Term Hypoxia Acclimation in Zebrafish (Danio rerio)


Abstract
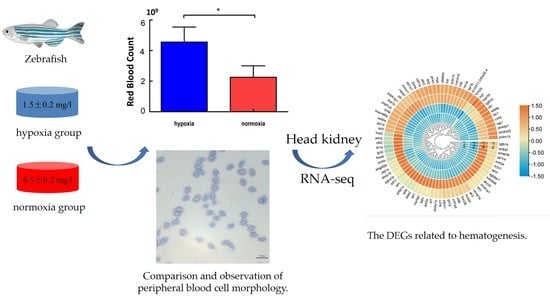
:
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Red Blood Cell Count
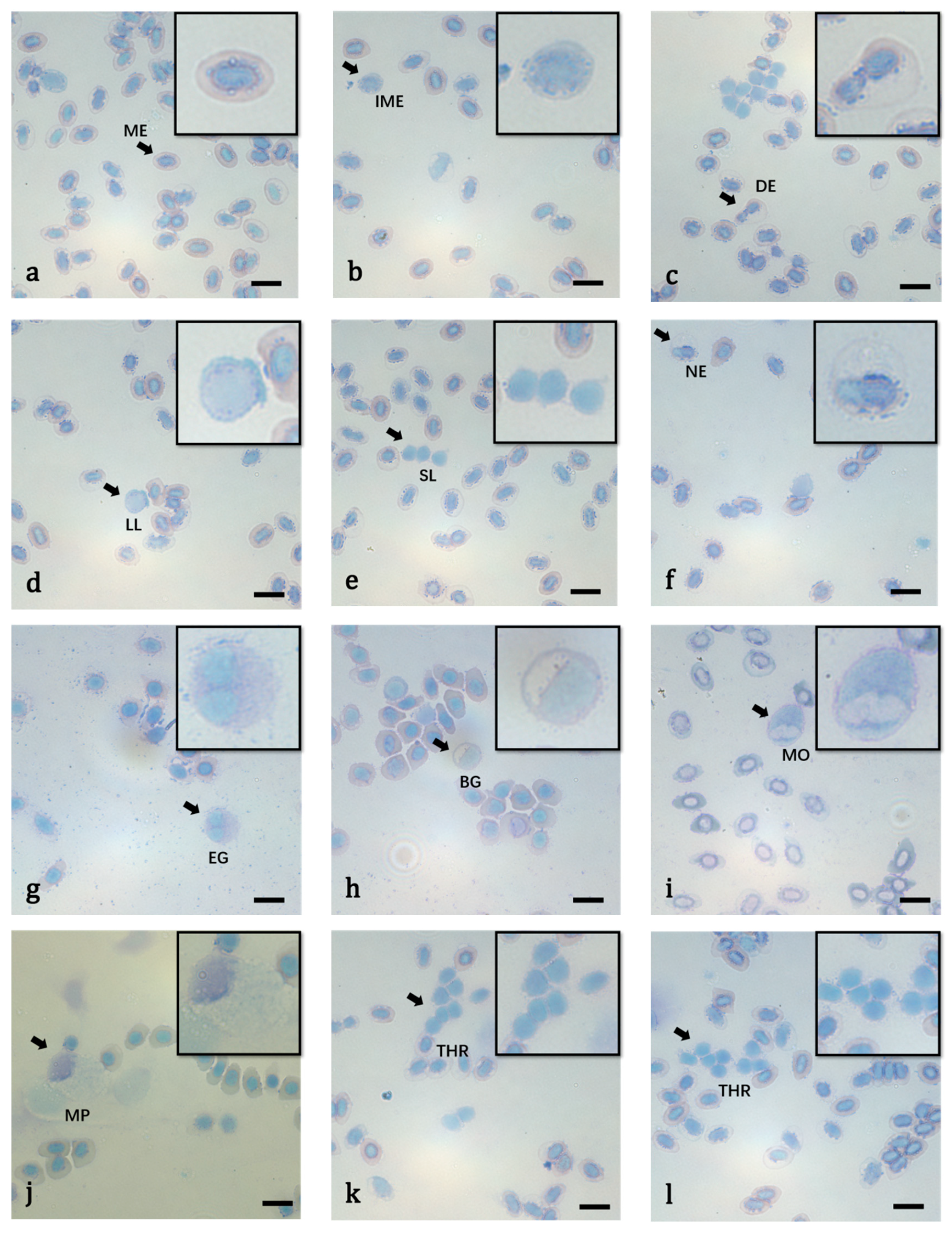
2.3. Light Microscopy Study
2.4. RNA Isolation, cDNA Library Construction and Illumina Sequencing
2.5. Differentially Expressed Gene Analysis and Functional Enrichment
2.6. Quantitative Real-Time PCR
2.7. Statistical Analysis
3. Results
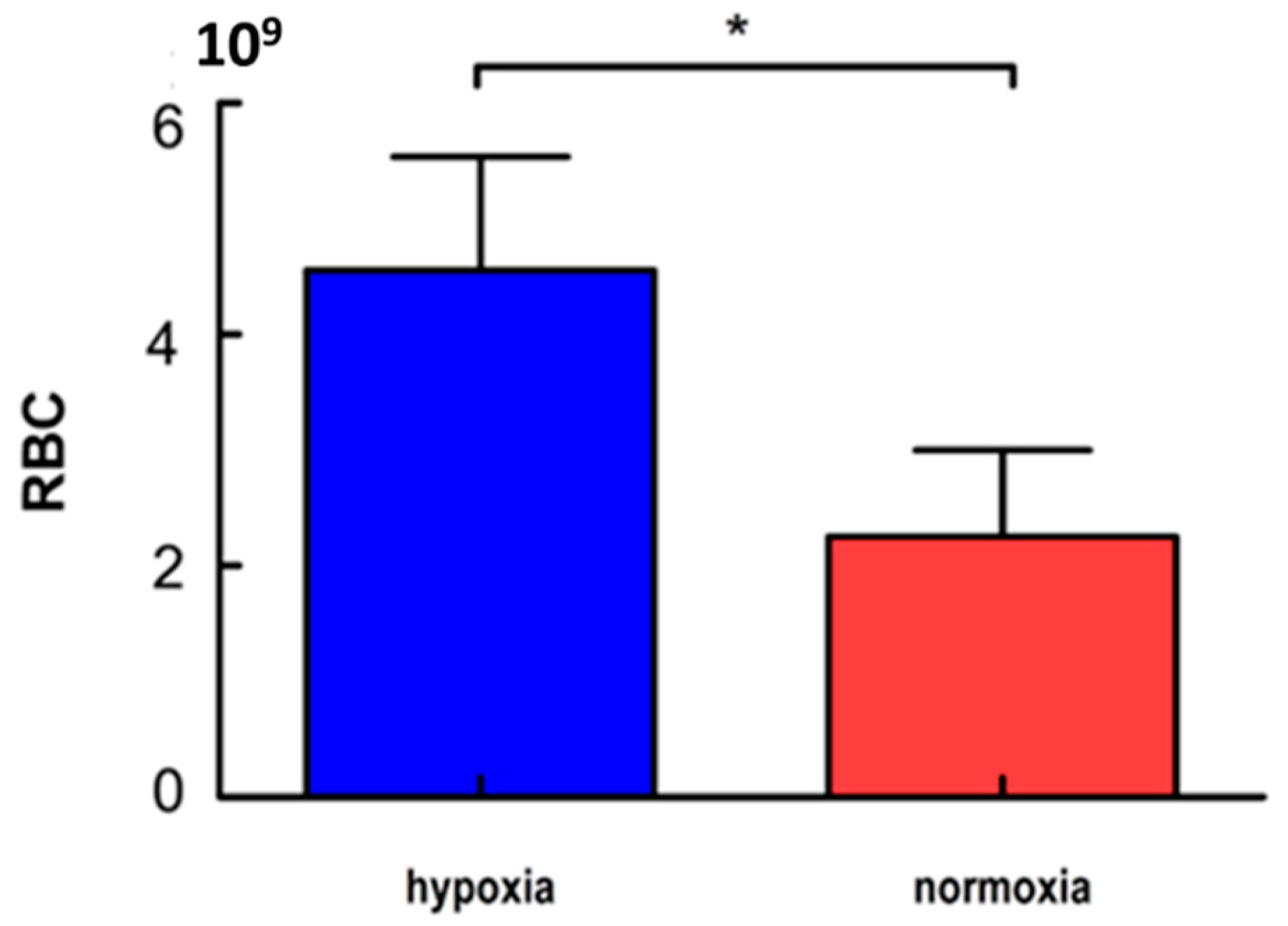
3.1. Erythrocyte Counts
3.2. Cell Morphology
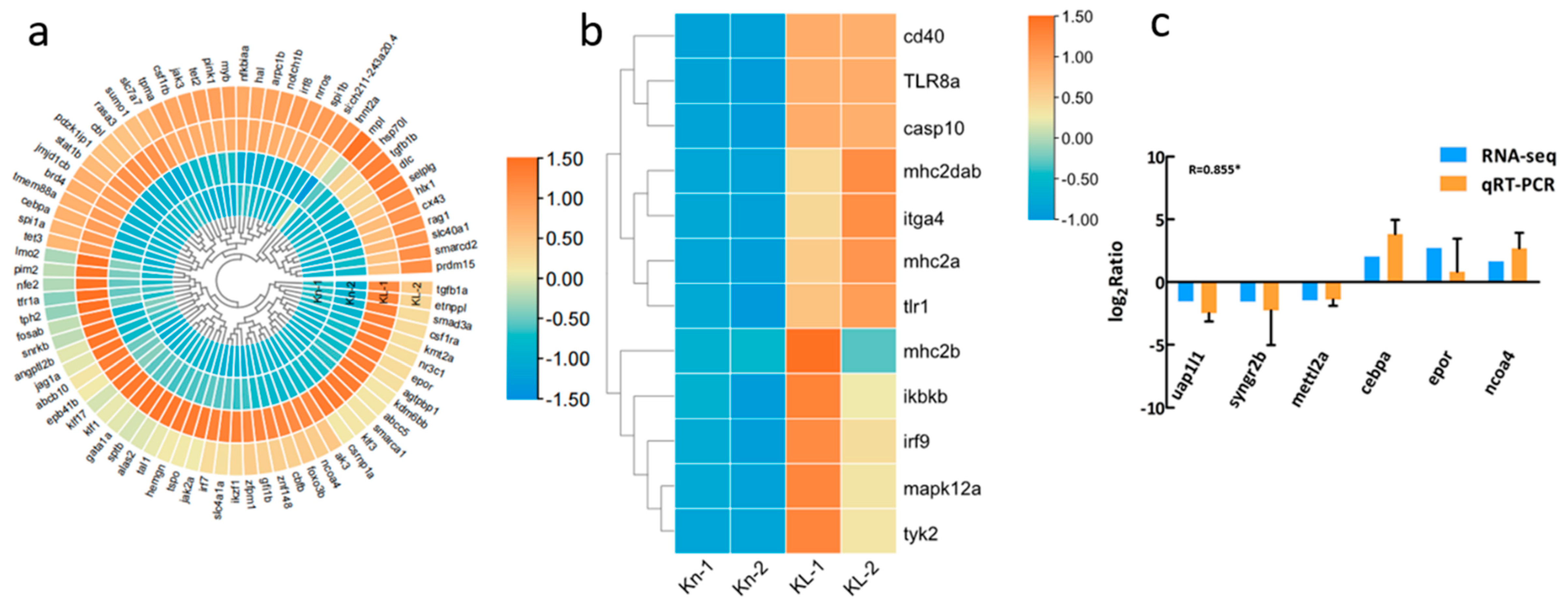
3.3. Analysis of the Differentially Expressed Genes
3.4. Transcriptome Data Validation by qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dabrowski, K.; Lee, K.J.; Guz, L.; Verlhac, V.; Gabaudan, J. Effects of dietary ascorbic acid on oxygen stress (hypoxia or hyperoxia), growth and tissue vitamin concentrations in juvenile rainbow trout (Oncorhynchus mykiss). Aquaculture 2003, 233, 383–392. [Google Scholar] [CrossRef]
- Pang, X.; Cao, Z.D.; Fu, S.J. The effects of temperature on metabolic interaction between digestion and locomotion in juveniles of three cyprinid fish (Carassius auratus, Cyprinus carpio and Spinibarbus sinensis). Comp. Biochem. Physiol. 2011, 159, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, G.E.; Renshaw, G.M.C. Hypoxic survival strategies in two fishes: Extreme anoxia tolerance in the North European crucian carp and natural hypoxic preconditioning in a coral-reef shark. J. Exp. Biol. 2004, 207, 3131–3139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sara, C.; Alex, D.; Joseph, M.F.; Daniel, S.D.; Gregory, A.L. A comparison of blood gases, biochemistry, and hematology to ecomorphology in a health assessment of pinfish (Lagodon rhomboides). PeerJ 2016, 4, e2262. [Google Scholar] [CrossRef] [Green Version]
- Ren, P.L.; Zhang, Y.M.; Geng, G.Q.; Qi, Y.M. Changes in Morphology and Quantity of Peripheral Blood Cells in Carassius auratus Collected from Polluted Water Area. Chin. J. Zool. 2008, 43, 37–42. [Google Scholar] [CrossRef]
- Haase, V.H. Hypoxic regulation of erythropoiesis and iron metabolism. American journal of physiology. Ren. Physiol. 2010, 299, F1–F13. [Google Scholar] [CrossRef] [Green Version]
- Soivio, A.; Nikinmaa, M.; Westman, K. The blood oxygen binding properties of hypoxic Salmo gairdneri. J. Comp. Physiol. 1980, 136, 83–87. [Google Scholar] [CrossRef]
- Suda, T.; Takubo, K.; Semenza, G.L. Metabolic Regulation of Hematopoietic Stem Cells in the Hypoxic Niche. Cell Stem Cell 2011, 9, 298–310. [Google Scholar] [CrossRef] [Green Version]
- Imanirad, P.; Dzierzak, E. Hypoxia and HIFs in regulating the development of the hematopoietic system. Blood Cells Mol. Dis. 2013, 51, 256–263. [Google Scholar] [CrossRef] [Green Version]
- Kettleborough, R.N.W.; Busch-Nentwich, E.M.; Harvey, S.A.; Dooley, C.M.; de Bruijn, E.; van Eeden, F.; Sealy, I.; White, R.J.; Herd, C.; Nijman, I.J.; et al. A systematic genome-wide analysis of zebrafish protein-coding gene function. Nature 2013, 496, 494–497. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.M.; Esain, V.; Frechette, G.M.; Harris, L.J.; Cox, A.G.; Cortes, M.; Garnaas, M.K.; Carroll, K.J.; Cutting, C.C.; Khan, T.; et al. Glucose metabolism impacts the spatiotemporal onset and magnitude of HSC induction in vivo. Blood 2013, 121, 2483–2493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paffett-Lugassy, N.; Hsia, N.; Fraenkel, P.G.; Paw, B.; Leshinsky, I.; Barut, B.; Bahary, N.; Caro, J.; Handin, R.; Zon, L.I. Functional conservation of erythropoietin signaling in zebrafish. Blood 2007, 110, 2718–2726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Z.H.; Wang, M.Q.; Wang, X.D.; Wang, L.L.; Qiu, L.M.; Song, L.S. Transcriptome sequencing reveals the involvement of reactive oxygen species in the hematopoiesis from Chinese mitten crab Eriocheir sinensis. Dev. Comp. Immunol. 2018, 82, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Klomp, J.; Hyun, J.; Klomp, J.E.; Pajcini, K.; Rehman, J.; Malik, A.B. Comprehensive transcriptomic profiling reveals SOX7 as an early regulator of angiogenesis in hypoxic human endothelial cells. J. Biol. Chem. 2020, 295, 4796–4808. [Google Scholar] [CrossRef] [Green Version]
- Léger, J.A.D.; Athanasio, C.G.; Zhera, A.; Chauhan, M.F.; Simmons, D.B.D. Hypoxic responses in Oncorhynchus mykiss involve angiogenesis, lipid, and lactate metabolism, which may be triggered by the cortisol stress response and epigenetic methylation. Comp. Biochem. Physiol.—Part D Genom. Proteom. 2021, 39, 100860. [Google Scholar] [CrossRef]
- Feng, J.F.; Guo, Y.; Gao, Y.F.; Zhu, L. Effects of Hypoxia on the Physiology of Zebrafish (Danio rerio): Initial Responses, Acclimation and Recovery. Bull. Environ. Contam. Toxicol. 2016, 96, 43–48. [Google Scholar] [CrossRef]
- Rees, B.B.; Sudradjat, F.A.; Love, J.W. Acclimation to hypoxia increases survival time of zebrafish, Danio rerio, during lethal hypoxia. J. Exp. Zool. 2001, 289, 266–272. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.J.; Wang, W.M. Haematological and biochemical characteristics of two aquacultured carnivorous cyprinids, topmouth culter Culter alburnus (Basilewsky) and yellowcheek carp Elopichthys bambusa (Richardson). Aquac. Res. 2010, 41, 1331–1338. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Daehwan, K.; Ben, L.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Cole, T.; Adam, R.; Loyal, G.; Geo, P.; Daehwan, K.; Kelley, D.R.; Harold, P.; Salzberg, S.L.; Rinn, J.L.; Lior, P. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.J.; Zhou, W.B.; Wang, B. Identification of crucial genes associated with lung adenocarcinoma by bioinformatic analysis. Medicine 2020, 99, e23052–e23062. [Google Scholar] [CrossRef]
- Cao, Y.L.; Tang, W.H.; Tang, W.X. Immune cell infiltration characteristics and related core genes in lupus nephritis: Results from bioinformatic analysis. BMC Immunol. 2019, 20, 37. [Google Scholar] [CrossRef] [Green Version]
- Bracamonte, S.E.; Johnston, P.R.; Knopf, K.; Monaghan, M.T. Experimental infection with Anguillicola crassus alters immune gene expression in both spleen and head kidney of the European eel (Anguilla anguilla). Mar. Genom. 2019, 45, 28–37. [Google Scholar] [CrossRef]
- Storey, J.D. A Direct Approach to False Discovery Rates. J. R. Stat. Soc. Ser. B (Stat. Methodol.) 2002, 64, 479–498. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.X.; Yi, S.K.; Wang, W.F.; He, Y.; Huang, Y.; Gao, Z.X.; Liu, H.; Wang, W.M.; Wang, H.L. Transcriptome comparison reveals insights into muscle response to hypoxia in blunt snout bream (Megalobrama amblycephala). Gene 2017, 624, 6–13. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Patterson, L.J.; Gering, M.; Eckfeldt, C.E.; Green, A.R.; Verfaillie, C.M.; Ekker, S.C.; Patient, R. The transcription factors Scl and Lmo2 act together during development of the hemangioblast in zebrafish. Blood 2007, 109, 2389–2398. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, J.; Hagen, A.; Hsu, K.; Deng, M.; Liu, T.X.; Look, A.T.; Kanki, J.P. Interplay of pu.1 and gata1 determines myelo-erythroid progenitor cell fate in zebrafish. Dev. Cell 2005, 8, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Zhu, P.P.; Tang, Y.; Fan, J.D.; Fang, J.; Peng, X.; Cui, H.M. Hematological Parameters and Blood Cell Morphology of Male and Female Schizothorax (Racoma) davidi (Sauvage). J. World Aquac. Soc. 2017, 48, 821–830. [Google Scholar] [CrossRef]
- Burrows, A.S.; Fletcher, T.C.; Manning, M.J. Haematology of the turbot, Psetta maxima (L.): Ultrastructural, cytochemical and morphological properties of peripheral blood leucocytes. J. Appl. Ichthyol. 2001, 17, 77–84. [Google Scholar] [CrossRef]
- Bert, P. La pression barométrique: Recherches de physiologie expérimentale; Masson: Paris, France, 1878. [Google Scholar]
- Viault, F. Sur l’augmentation considerable du nombre des globules rouges dans sang chez les habitants des haut plateauz de l’Amerique de Sud. Comp. Rend. Acad. Sci. 1890, 111, 917–918. [Google Scholar]
- Lin, Y.H. Factors Influencing the Hematology of Fishes. J. Anhui Agric. Sci. 2011, 39, 8657–8659. [Google Scholar] [CrossRef]
- Kupittayanant, P.; Kinchareon, W. Hematological and biochemical responses of the flowerhorn fish to hypoxia. J. Anim. Vet. Adv. 2011, 10, 2631–2638. [Google Scholar] [CrossRef]
- Chen, Y.X.; Xiao, L.Y.; Yan, T.M.; Zhao, H.T.; Shen, S.J.; Zhou, D.G. Hematological of wild and cultured schizothoracin fishes. Acta Hydrobiol. Sin. 2009, 33, 905–910. [Google Scholar] [CrossRef]
- Lin, G.H.; Zhang, F.W. On Hematological Studies of Singuo Red Carp--Differential Blood Count and Normal Size of Blood Cells. J. Nanchang Univ. (Sci. Ed.) 1987, 11, 41–48. [Google Scholar]
- Qu, Y.M.; Athey, B.D.; Arabnia, H.R.; Sung, A.H.; Liu, Q.Z.; Yang, J.Y.; Mao, J.H.; Deng, Y.P. High-throughput next-generation sequencing technologies foster new cutting-edge computing techniques in bioinformatics. BMC Genom. 2009, 10 (Suppl. 1), 1–3. [Google Scholar] [CrossRef] [Green Version]
- Oburoglu, L.; Romano, M.; Taylor, N.; Kinet, S. Metabolic regulation of hematopoietic stem cell commitment and erythroid differentiation. Curr. Opin. Hematol. 2016, 23, 198–205. [Google Scholar] [CrossRef]
- Wingert, R.A.; Brownlie, A.; Galloway, J.L.; Dooley, K.; Fraenkel, P.; Axe, J.L.; Davidson, A.J.; Barut, B.; Noriega, L.; Sheng, X.M.; et al. The chianti zebrafish mutant provides a model for erythroid-specific disruption of transferrin receptor 1. Development 2004, 131, 6225–6235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hattangadi, S.M.; Wong, P.; Zhang, L.; Flygare, J.; Lodish, H.F. From stem cell to red cell: Regulation of erythropoiesis at multiple levels by multiple proteins, RNAs, and chromatin modifications. Blood 2011, 118, 6258–6268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhrt, D.; Wojchowski, D.M. Emerging EPO and EPO receptor regulators and signal transducers. Blood 2015, 125, 3536–3541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gering, M.; Rodaway, A.R.; Göttgens, B.; Patient, R.K.; Green, A.R. The SCL gene specifies haemangioblast development from early mesoderm. EMBO J. 1998, 17, 4029–4045. [Google Scholar] [CrossRef] [Green Version]
- Lyons, S.E.; Lawson, N.D.; Lei, L.; Bennett, P.E.; Weinstein, B.M.; Liu, P.P. A nonsense mutation in zebrafish gata1 causes the bloodless phenotype in vlad tepes. Proc. Natl. Acad. Sci. USA 2002, 99, 5454–5459. [Google Scholar] [CrossRef] [Green Version]
- Tamassia, N.; Aguilera, B.F.; Gasperini, S.; Polletti, S.; Gardiman, E.; Ostuni, R.; Natoli, G.; Cassatella, M.A. Induction of OCT2 contributes to regulate the gene expression program in human neutrophils activated via TLR8. Cell Rep. 2021, 35, 109143–109165. [Google Scholar] [CrossRef]
- Morris, A.; Hewitt, C.; Young, S. The major histocompatibility complex: Its genes and their roles in antigen presentation. Mol. Asp. Med. 1994, 15, 377–503. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acc. Number | Gene | Primers Sequences (5′ to 3′) | Product Length |
|---|---|---|---|
| NM_200294.1 | uap1l1 | F-TGAGGGACGTGGCAGAAAAG | 80 |
| R-TTCTCCATCCACAAACGGCA | |||
| NM_001082858.1 | syngr2b | F-TTCCTCAGCTGGGTGTTTGC | 179 |
| R-TCAGCCAATAGGAAGCCCAC | |||
| NM_001017902.1 | mettl2a | F-AGCACAACGCGTGGGATAAT | 91 |
| R-GCTGGGAGAGGTTGACTGTT | |||
| NM_131885.2 | Cebpa | F-AGTCGGGACAAGGCGAAAAT | 176 |
| R-CAGTTGCCCATGGCTTTGAC | |||
| NM_001043334.1 | Epor | F-GTCACATGGGCAAGGTGGAG | 116 |
| R-ATAAGAAACGCCGTCCGGTT | |||
| NM_201129.1 | ncoa4 | F-GAGAGGACGCGGCAGTATTT | 102 |
| R-CCCCATAGGAGACATACTGGGT | |||
| NM_131031.2 | β-Actin | F-CACTGAGGCTCCCCTGAATC | 167 |
| R-GGGTCACACCATCACCAGAG |
| Normoxia | Hypoxia | |||||
|---|---|---|---|---|---|---|
| Parameters | Length (Mean ± SD) | Width (Mean ± SD) | n | Length (Mean ± SD) | Width (Mean ± SD) | n |
| Erythrocyte | 11.51 ± 0.88 | 8.23 ± 0.83 | 50 | 10.81 ± 0.72 ** | 7.19 ± 0.69 ** | 50 |
| Nucleus size of erythrocyte | 6.36 ± 0.55 | 4.06 ± 0.60 | 50 | 6.39 ± 0.54 | 3.45 ± 0.39 ** | 50 |
| Neutrophil | 11.52 ± 1.15 | 10.35 ± 1.19 | 40 | 11.23 ± 1.26 | 9.83 ± 1.40 | 30 |
| Nucleus size of neutrophil | 9.02 ± 1.42 | 6.47 ± 1.00 | 40 | 10.67 ± 11.19 | 6.29 ± 1.09 | 30 |
| Eosnophils | 12.13 ± 1.34 | 11.07 ± 1.25 | 30 | 10.25 ± 1.41 ** | 8.88 ± 1.14 ** | 20 |
| Nucleus size of eosnophils | 8.34 ± 1.71 | 5.94 ± 1.11 | 30 | 7.77 ± 1.37 | 5.97 ± 0.90 | 20 |
| Basophilic granulocyte | 10.99 ± 1.91 | 9.97 ± 1.46 | 20 | 9.72 ± 1.38 * | 9.03 ± 1.43 * | 20 |
| Nucleus size of basophilic granulocyte | 8.95 ± 1.71 | 4.98 ± 1.51 | 20 | 7.41 ± 1.23 ** | 4.58 ± 1.25 | 20 |
| Small lymphocyte | 6.33 ± 0.50 | 5.48 ± 0.76 | 50 | 6.32 ± 0.43 | 5.08 ± 0.86 * | 50 |
| Nucleus size of small lymphocyte | 5.54 ± 0.60 | 4.70 ± 0.59 | 50 | 6.03 ± 0.58 ** | 4.81 ± 0.76 | 50 |
| Large lymphocyte | 7.98 ± 0.77 | 6.31 ± 0.90 | 50 | 8.41 ± 1.21 * | 6.15 ± 1.03 | 50 |
| Nucleus size of large lymphocyte | 6.72 ± 1.01 | 5.49 ± 0.78 | 50 | 7.61 ± 0.91 ** | 5.66 ± 0.89 | 50 |
| Monocyte | 12.08 ± 1.09 | 10.96 ± 1.32 | 40 | 12.56 ± 1.41 | 11.21 ± 1.35 | 30 |
| Nucleus size of monocyte | 10.07 ± 1.24 | 6.66 ± 1.34 | 40 | 9.54 ± 1.26 | 6.11 ± 1.31 | 30 |
| Thrombocyte | 6.86 ± 1.10 | 5.08 ± 1.02 | 50 | 6.78 ± 1.40 | 4.12 ± 0.74 ** | 50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, L.; Chen, L.; Jiang, S.; Wu, Z.; Zhou, Y.; Xu, Q. Hematogenesis Adaptation to Long-Term Hypoxia Acclimation in Zebrafish (Danio rerio). Fishes 2022, 7, 98. https://doi.org/10.3390/fishes7030098
Shi L, Chen L, Jiang S, Wu Z, Zhou Y, Xu Q. Hematogenesis Adaptation to Long-Term Hypoxia Acclimation in Zebrafish (Danio rerio). Fishes. 2022; 7(3):98. https://doi.org/10.3390/fishes7030098
Chicago/Turabian StyleShi, Linghui, Lei Chen, Shouwen Jiang, Zhichao Wu, Yan Zhou, and Qianghua Xu. 2022. "Hematogenesis Adaptation to Long-Term Hypoxia Acclimation in Zebrafish (Danio rerio)" Fishes 7, no. 3: 98. https://doi.org/10.3390/fishes7030098
APA StyleShi, L., Chen, L., Jiang, S., Wu, Z., Zhou, Y., & Xu, Q. (2022). Hematogenesis Adaptation to Long-Term Hypoxia Acclimation in Zebrafish (Danio rerio). Fishes, 7(3), 98. https://doi.org/10.3390/fishes7030098