
Blood Transcriptome Analysis Reveals Gene Expression Differences between Yangtze Finless Porpoises from Two Habitats: Natural and Ex Situ Protected Waters


Abstract
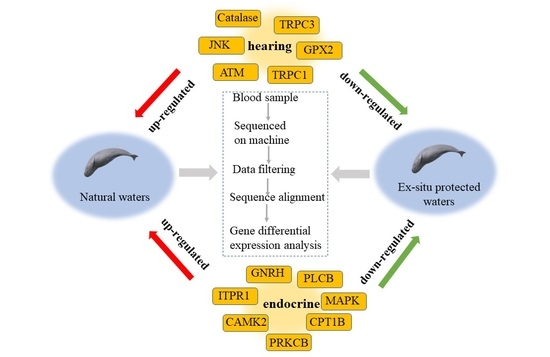
:
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Samples Collection
2.3. Total RNA Extraction, mRNA Sequencing and Analysis
2.4. Differentially Expressed Gene Screening and Enrichment Analysis
2.5. Real-Time Quantitative PCR Validation
2.6. Supporting Information
3. Results
3.1. Sequencing Data and Alignment Analysis
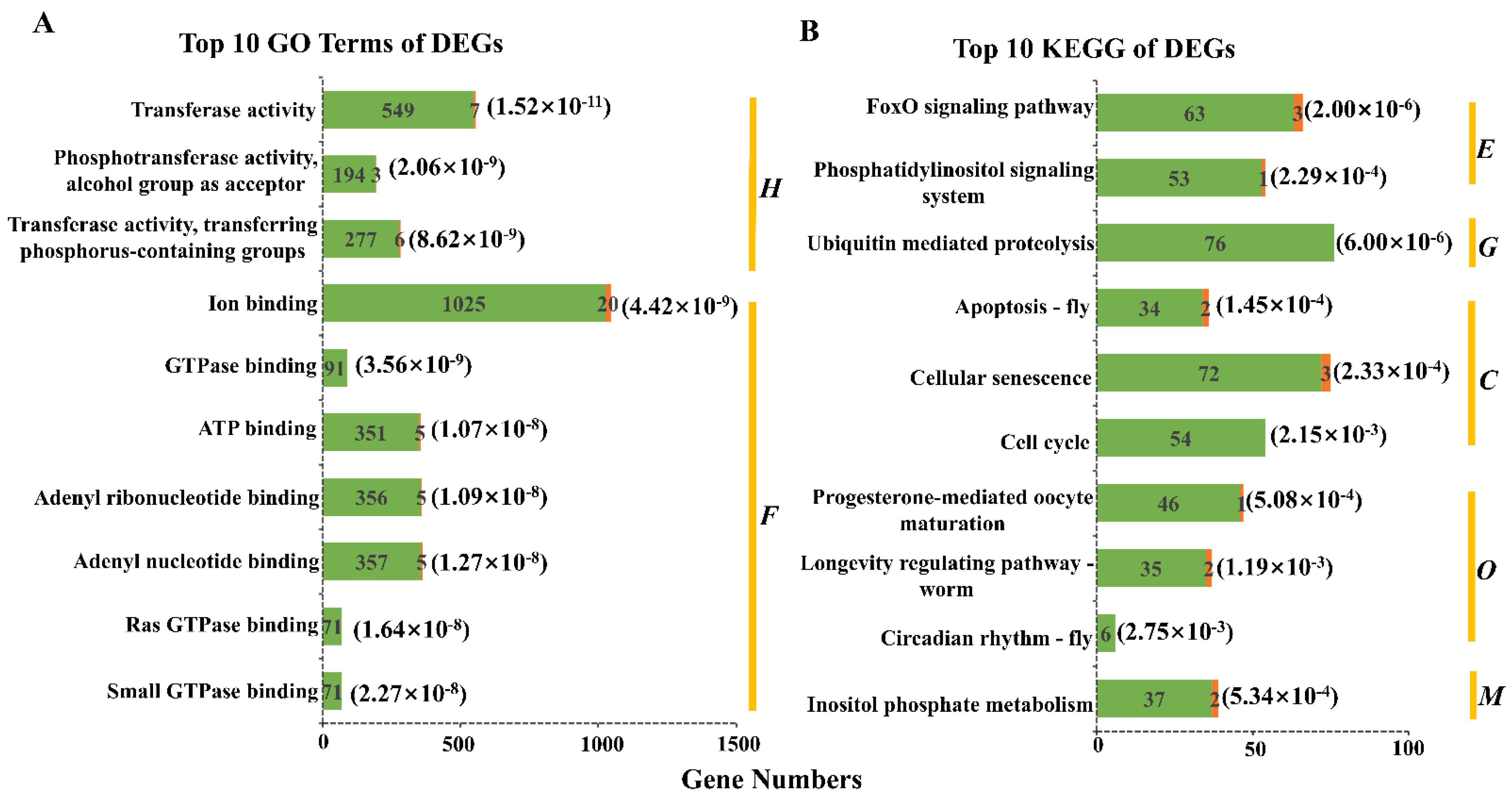
3.2. Analysis and Functional Annotation of Differentially Expressed Genes
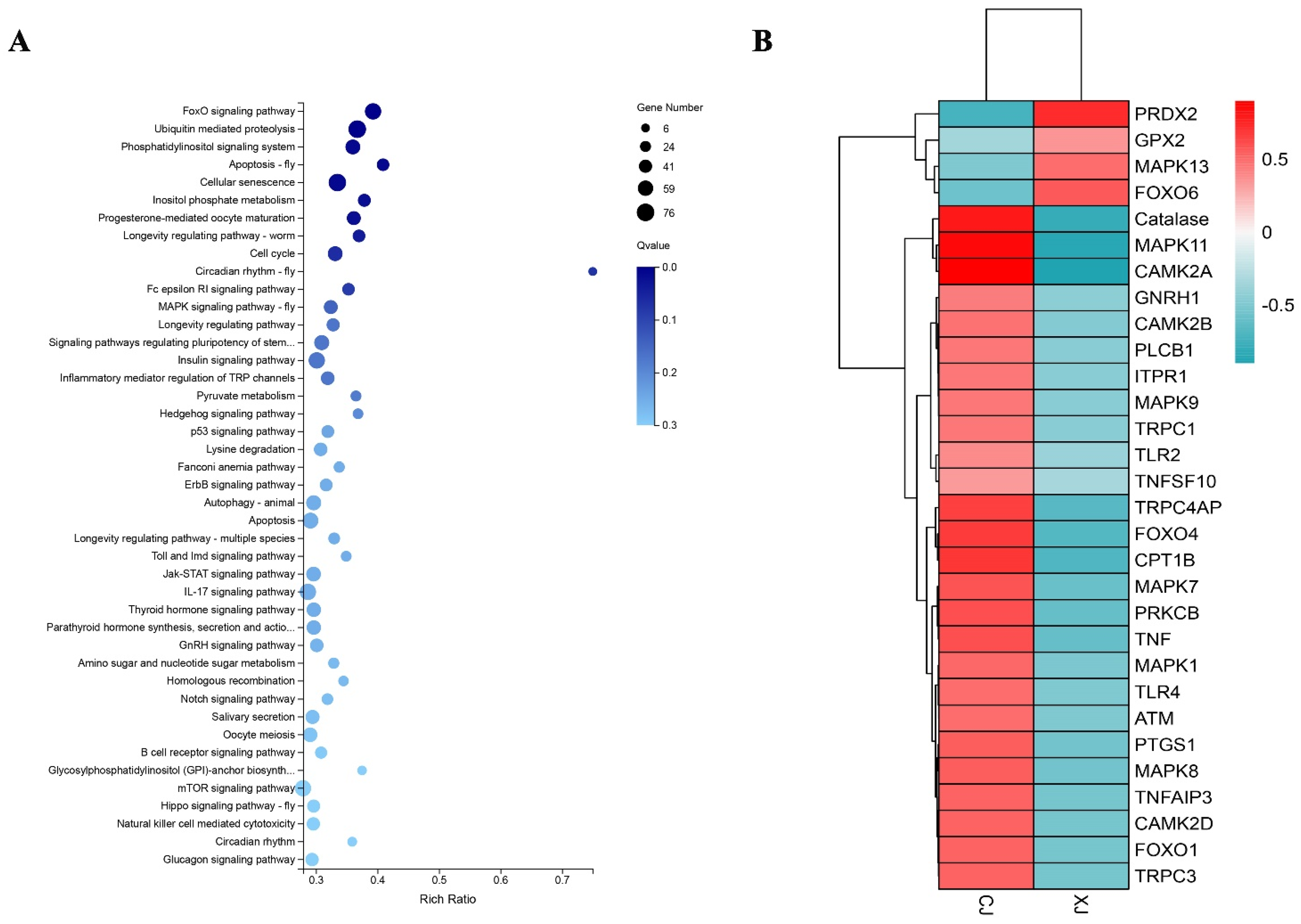
3.3. Enrichment Analysis of Differentially Expressed Genes
3.4. Analysis of Key Genes
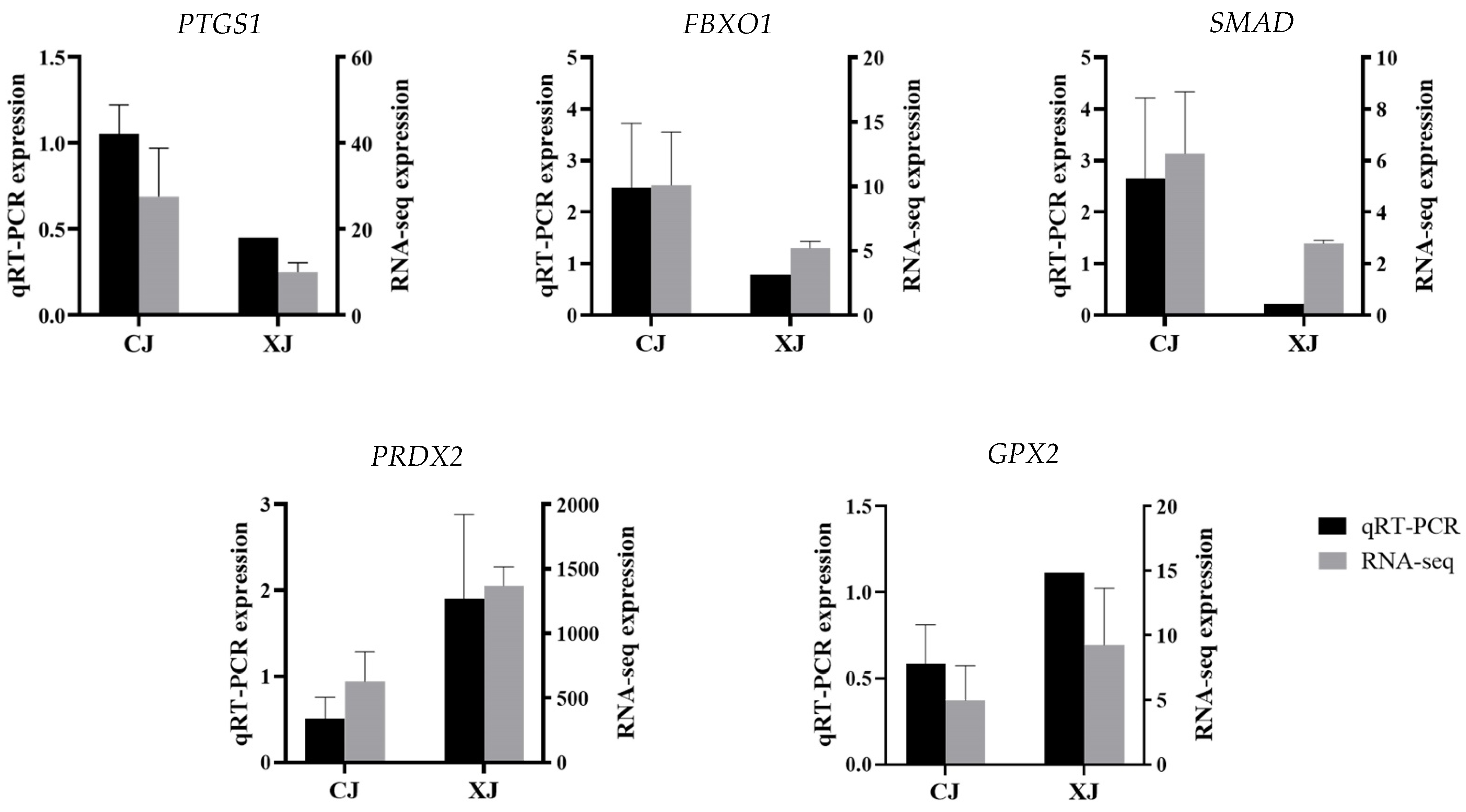
3.5. Validation of RNA-Seq Results by qRT-PCR
4. Discussion
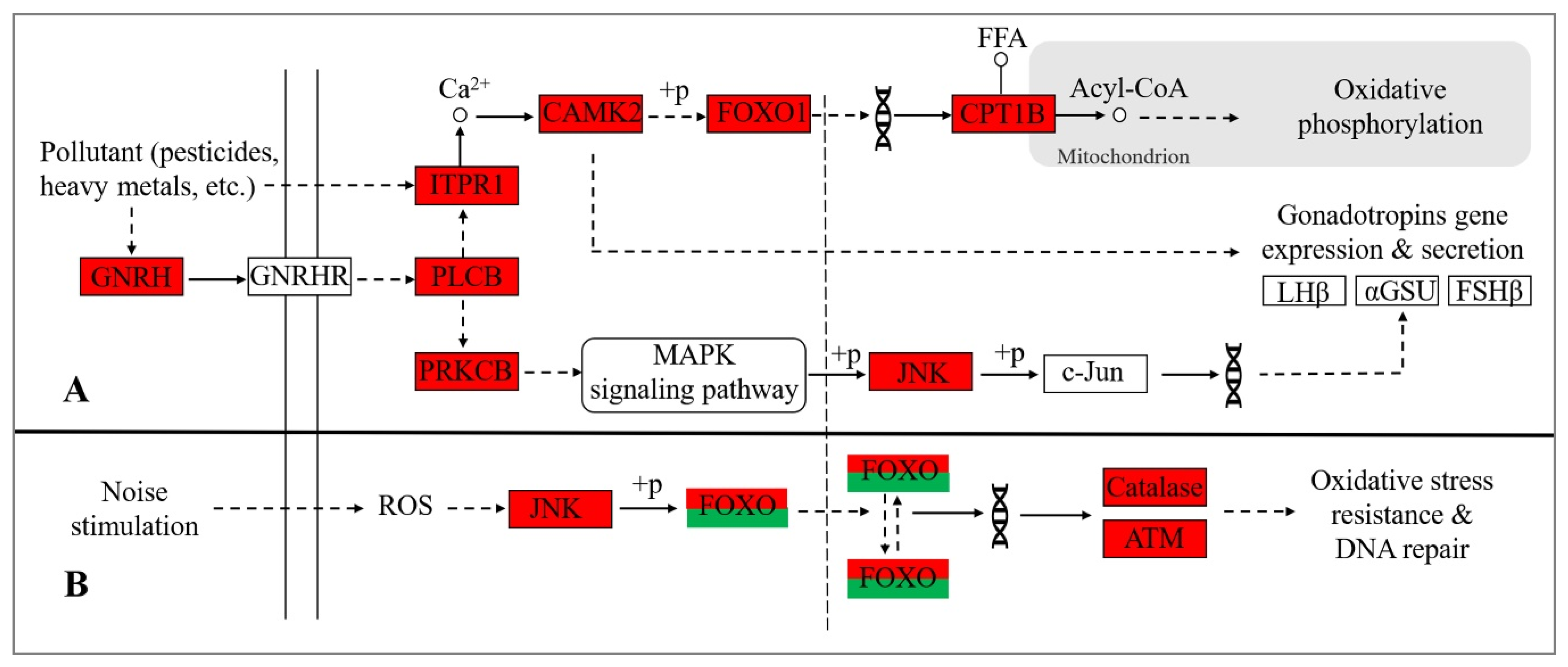
4.1. Water Pollution May Interfere with the Endocrine Activity of Yangtze Finless Porpoise
4.2. Noise Pollution May Interfere with the Hearing Function of Yangtze Finless Porpoise
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344, 1246752. [Google Scholar] [CrossRef] [PubMed]
- McGowan, P.J.; Traylor-Holzer, K.; Leus, K. IUCN Guidelines for Determining When and How Ex Situ Management Should Be Used in Species Conservation. Conserv. Lett. 2016, 10, 361–366. [Google Scholar] [CrossRef]
- Kleinman-Ruiz, D.; Soriano, L.; Casas-Marce, M.; Szychta, C.; Sánchez, I.; Fernández, J.; Godoy, J.A. Genetic evaluation of the Iberian lynx ex situ conservation programme. Heredity 2019, 123, 647–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Oliveira, M.J.; Aguiar-Silva, F.H.; de Moraes, W.; Sanaiotti, T.M.; Banhos, A.; Moreira, N. Ex situ population of the Harpy Eagle and its potential for integrated conservation. ZooKeys 2022, 1083, 109–128. [Google Scholar] [CrossRef] [PubMed]
- Turvey, S.T.; Pitman, R.L.; Taylor, B.L.; Barlow, J.; Akamatsu, T.; Barrett, L.A.; Zhao, X.; Reeves, R.R.; Stewart, B.S.; Wang, K.; et al. First human-caused extinction of a cetacean species? Biol. Lett. 2007, 3, 537–540. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Turvey, S.; Zhao, X.; Mei, Z. Neophocaena asiaeorientalis ssp. asiaeorientalis. In The IUCN Red List of Threatened Species; Version 3.1. 2013. Available online: http://www.iucnredlist.org (accessed on 20 February 2021).
- Mei, Z.; Chen, M.; Han, Y.; Hao, Y.; Zheng, J.; Wang, K.; Wang, D. Thresholds of population persistence for the Yangtze finless porpoise: Implications for conservation managements. Integr. Zool. 2021, 16, 538–547. [Google Scholar] [CrossRef]
- Zhang, X.; Yu, D.; Wang, H.; Wan, A.; Chen, M.; Tao, F.; Song, Z. Effects of fish community on occurrences of Yangtze finless porpoise in confluence of the Yangtze and Wanhe Rivers. Environ. Sci. Pollut. Res. 2015, 22, 9524–9533. [Google Scholar] [CrossRef]
- Han, H.; Li, H.; Zhang, K. Spatial-Temporal Coupling Analysis of the Coordination between Urbanization and Water Ecosystem in the Yangtze River Economic Belt. Int. J. Environ. Res. Public Health 2019, 16, 3757. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.-T.; Akamatsu, T.; Duan, P.-X.; Zhou, L.; Yuan, J.; Li, J.; Lei, P.-Y.; Chen, Y.-W.; Yang, Y.-N.; Wang, K.-X.; et al. Underwater noise pollution in China’s Yangtze River critically endangers Yangtze finless porpoises (Neophocaena asiaeorientalis asiaeorientalis). Environ. Pollut. 2020, 262, 114310. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Qian, Z.; Ruan, Y.; Hao, Y.; Dong, W.; Li, K.; Mei, Z.; Wang, K.; Wu, C.; Wu, J.; et al. First evaluation of legacy persistent organic pollutant contamination status of stranded Yangtze finless porpoises along the Yangtze River Basin, China. Sci. Total Environ. 2020, 710, 136446. [Google Scholar] [CrossRef]
- Whitehead, A.; Triant, D.A.; Champlin, D.; Nacci, D. Comparative transcriptomics implicates mechanisms of evolved pollution tolerance in a killifish population. Mol. Ecol. 2010, 19, 5186–5203. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, E.L.; Jaszczyszyn, Y.; Naquin, D.; Thermes, C. The Third Revolution in Sequencing Technology. Trends Genet. 2018, 34, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Huang, Y.; Wu, H.; Li, C.; Ling, S.; Sun, J.; Shen, H.; Yue, B.; Zhang, X. Blood transcriptome analysis revealed the immune changes and immunological adaptation of wildness training giant pandas. Mol. Genet. Genom. 2022, 297, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Remy, S.; Govarts, E.; Wens, B.; De Boever, P.; Hond, E.D.; Croes, K.; Sioen, I.; Baeyens, W.; Van Larebeke, N.; Koppe, J.; et al. Metabolic targets of endocrine disrupting chemicals assessed by cord blood transcriptome profiling. Reprod. Toxicol. 2016, 65, 307–320. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, J.; Wang, L.; Tian, H.; Sui, J. Transcriptome analysis revealed potential mechanisms of differences in physiological stress responses between caged male and female magpies. BMC Genom. 2019, 20, 447. [Google Scholar] [CrossRef]
- Nabi, G.; Hao, Y.; Robeck, T.R.; Jinsong, Z.; Wang, D. Physiological consequences of biologic state and habitat dynamics on the critically endangered Yangtze finless porpoises (Neophocaena asiaeorientalis ssp. asiaeorientalis) dwelling in the wild and semi-natural environment. Conserv. Physiol. 2018, 6, coy072. [Google Scholar]
- Nabi, G.; Li, Y.; McLaughlin, R.W.; Mei, Z.; Wang, K.; Hao, Y.; Zheng, J.; Wang, D. Immune Responses of the Critically Endangered Yangtze Finless Porpoises (Neophocaena asiaeorientalis ssp. asiaeorientalis) to Escalating Anthropogenic Stressors in the Wild and Seminatural Environments. Front. Physiol. 2020, 10, 1594. [Google Scholar] [CrossRef]
- Wang, Z.-T.; Duan, P.-X.; Akamatsu, T.; Chen, Y.-W.; An, X.; Yuan, J.; Lei, P.-Y.; Li, J.; Zhou, L.; Liu, M.-C.; et al. Riverside underwater noise pollution threaten porpoises and fish along the middle and lower reaches of the Yangtze River, China. Ecotoxicol. Environ. Saf. 2021, 226, 112860. [Google Scholar] [CrossRef]
- Hao, Y.-J.; Zhao, Q.-Z.; Wu, H.-P.; Chen, D.-Q.; Gong, C.; Li, L.; Wang, D. Physiological responses to capture and handling of free-ranging male Yangtze finless porpoises (Neophocaena phocaenoides asiaeorientalis). Mar. Freshw. Behav. Physiol. 2009, 42, 315–327. [Google Scholar] [CrossRef]
- Li, R.; Li, Y.; Kristiansen, K.; Wang, J. SOAP: Short oligonucleotide alignment program. Bioinformatis 2008, 24, 713–714. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, D.; Lin, D.; Guo, H.; Gu, H.; Ying, C.; Zhang, Y.; Zhang, J.; Liu, K.; Tang, W. Integrated analysis of blood mRNAs and microRNAs reveals immune changes with age in the Yangtze finless porpoise (Neophocaena asiaeorientalis). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2021, 256, 110635. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, H.A. On a general theoretical foundation for endocrinology. Sci. Prog. 2019, 102, 43–60. [Google Scholar] [CrossRef] [Green Version]
- Dai, C.; Tang, B.; Hao, Y.; Wang, K.; Gong, C.; Yuan, J. Study on the health evaluation system of Yangtze finless porpoise (Neophocaena asiaeorientalis asiaeorientalis). J. Anhui Agric. Univ. 2021, 48, 403. [Google Scholar]
- Sokal, A.; Jarmakiewicz-Czaja, S.; Tabarkiewicz, J.; Filip, R. Dietary Intake of Endocrine Disrupting Substances Presents in Environment and Their Impact on Thyroid Function. Nutrients 2021, 13, 867. [Google Scholar] [CrossRef]
- Graceli, J.B.; Dettogni, R.S.; Merlo, E.; Niño, O.; da Costa, C.S.; Zanol, J.F.; Rios-Morris, E.A.; Miranda-Alves, L.; Denicol, A.C. The impact of endocrine-disrupting chemical exposure in the mammalian hypothalamic-pituitary axis. Mol. Cell Endocrinol. 2020, 518, 110997. [Google Scholar] [CrossRef]
- Huang, T.; Zhao, Y.; He, J.; Cheng, H.; Martyniuk, C.J. Endocrine disruption by azole fungicides in fish: A review of the evidence. Sci. Total Environ. 2022, 822, 153412. [Google Scholar] [CrossRef]
- Yamindago, A.; Lee, N.; Lee, N.; Jo, Y.; Woo, S.; Yum, S. Fluoxetine in the environment may interfere with the neurotransmission or endocrine systems of aquatic animals. Ecotoxicol. Environ. Saf. 2021, 227, 112931. [Google Scholar] [CrossRef]
- Barra, R.O.; Chiang, G.; Saavedra, M.F.; Orrego, R.; Servos, M.R.; Hewitt, L.M.; McMaster, M.E.; Bahamonde, P.; Tucca, F.; Munkittrick, K.R. Endocrine Disruptor Impacts on Fish from Chile: The Influence of Wastewaters. Front. Endocrinol. 2021, 12, 208. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chen, Z.; Zhang, J.; Wu, S.; Yang, L.; Chen, L.; Shao, Y. The challenge of micropollutants in surface water of the Yangtze River. Sci. Total Environ. 2021, 780, 146537. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Tang, X.; Guo, W.; Lin, L.; Zhao, L.; Hu, Y.; Liu, M. Spatiotemporal distribution dynamics of heavy metals in water, sediment, and zoobenthos in mainstream sections of the middle and lower Changjiang River. Sci. Total Environ. 2020, 714, 136779. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.P.; Wang, J.C.; Schipma, M.J.; Zhang, X.; Edwards, J.R.; El Muayed, M. Cadmium-mediated pancreatic islet transcriptome changes in mice and cultured mouse islets. Toxicol. Appl. Pharmacol. 2021, 433, 115756. [Google Scholar] [CrossRef]
- Xi, W.; Lee, C.K.F.; Yeung, W.S.B.; Giesy, J.P.; Wong, M.H.; Zhang, X.; Hecker, M.; Wong, C.K.C. Effect of perinatal and postnatal bisphenol A exposure to the regulatory circuits at the hypothalamus–pituitary–gonadal axis of CD-1 mice. Reprod. Toxicol. 2011, 31, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Ramkumar, V.; Sheth, S.; Dhukhwa, A.; Al Aameri, R.; Rybak, L.P.; Mukherjea, D. Transient Receptor Potential (TRP) Channels and Auditory Functions. Antioxid. Redox Signal. 2021. [Google Scholar] [CrossRef] [PubMed]
- Tadros, S.F.; Kim, Y.; Phan, P.A.B.; Birnbaumer, L.; Housley, G.D. TRPC3 ion channel subunit immunolocalization in the cochlea. Histochem. Cell Biol. 2010, 133, 137–147. [Google Scholar] [CrossRef]
- Popov, V.V.; Supin, A.Y.; Wang, D.; Wang, K.; Dong, L.; Wang, S. Noise-induced temporary threshold shift and recovery in Yangtze finless porpoises Neophocaena phocaenoides asiaeorientalis. J. Acoust. Soc. Am. 2011, 130, 574–584. [Google Scholar] [CrossRef]
- Ju, T.; Zhang, T.C.; Wang, Z.T.; Xie, Y.; Zheng, C.H.; Wang, K.X.; Wang, D. Characteristics of riprapping underwater noise and its possible impacts on the Yangtze finless porpoise. Tech. Acoust. 2017, 36, 580–588. [Google Scholar]
- Frye, M.D.; Ryan, A.F.; Kurabi, A. Inflammation associated with noise-induced hearing loss. J. Acoust. Soc. Am. 2019, 146, 4020–4032. [Google Scholar] [CrossRef] [Green Version]
- Fuentes-Santamaría, V.; Alvarado, J.C.; Melgar-Rojas, P.; Gabaldón-Ull, M.C.; Miller, J.M.; Juiz, J.M. The Role of Glia in the Peripheral and Central Auditory System Following Noise Overexposure: Contribution of TNF-α and IL-1β to the Pathogenesis of Hearing Loss. Front. Neuroanat. 2017, 11, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abi-Hachem, R.N.; Zine, A.; Van De Water, T.R. The injured cochlea as a target for inflammatory processes, initiation of cell death pathways and application of related otoprotectives strategies. Recent Pat. CNS Drug Discov. 2010, 5, 147–163. [Google Scholar] [CrossRef] [PubMed]
- Arpornchayanon, W.; Canis, M.; Ihler, F.; Settevendemie, C.; Strieth, S. TNF-α inhibition using etanercept prevents noise-induced hearing loss by improvement of cochlear blood flow in vivo. Int. J. Audiol. 2013, 52, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Pyykkö, I.; Zou, J. Involvement of Ubiquitin-Editing Protein A20 in Modulating Inflammation in Rat Cochlea Associated with Silver Nanoparticle-Induced CD68 Upregulation and TLR4 Activation. Nanoscale Res. Lett. 2016, 11, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, S.K.; Woo, J.I.; Lee, H.Y.; Park, R.; Shimada, J.; Pan, H.; Gellibolian, R.; Lim, D.J. Toll-like receptor 2-dependent NF-κB activation is involved in nontypeable Haemophilus influenzae-induced monocyte chemotactic protein 1 up-regulation in the spiral ligament fibrocytes of the inner ear. Infect. Immun. 2007, 75, 3361–3372. [Google Scholar] [CrossRef] [Green Version]
- Waqas, M.; Gao, S.; Salam, I.U.; Ali, M.K.; Ma, Y.; Li, W. Inner Ear Hair Cell Protection in Mammals against the Noise-Induced Cochlear Damage. Neural Plast. 2018, 2018, 3170801. [Google Scholar] [CrossRef] [Green Version]
- Slepecky, N. Overview of mechanical damage to the inner ear: Noise as a tool to probe cochlear function. Hear. Res. 1986, 22, 307–321. [Google Scholar] [CrossRef]
- Fetoni, A.R.; De Bartolo, P.; Eramo, S.L.M.; Rolesi, R.; Paciello, F.; Bergamini, C.; Fato, R.; Paludetti, G.; Petrosini, L.; Troiani, D. Noise-Induced Hearing Loss (NIHL) as a Target of Oxidative Stress-Mediated Damage: Cochlear and Cortical Responses after an Increase in Antioxidant Defense. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 4011–4023. [Google Scholar] [CrossRef] [Green Version]
- Vurusaner, B.; Poli, G.; Basaga, H. Tumor suppressor genes and ROS: Complex networks of interactions. Free Radic. Biol. Med. 2012, 52, 7–18. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Raw Reads (M) | Total Clean Reads (M) | Total Clean Bases (Gb) | Clean Reads Q20 (%) | Clean Reads Q30 (%) | Total Mapping (%) | Uniquely Mapping (%) |
|---|---|---|---|---|---|---|---|
| CJ_1 | 59.93 | 58.21 | 5.82 | 98.02 | 94.36 | 78.56 | 41.76 |
| CJ_2 | 72.27 | 70.90 | 7.09 | 97.98 | 94.28 | 83.50 | 53.97 |
| CJ_3 | 101.56 | 100.27 | 10.03 | 98.03 | 94.43 | 87.75 | 63.36 |
| XJ_1 | 66.32 | 64.62 | 6.46 | 98.02 | 94.09 | 77.62 | 41.93 |
| XJ_2 | 69.75 | 67.55 | 6.76 | 98.09 | 94.28 | 73.90 | 34.36 |
| XJ_3 | 85.73 | 83.27 | 8.33 | 98.09 | 94.28 | 77.75 | 43.38 |
| Category | Pathway ID | Pathway Name | Genes Included (Up/Down) | p Value |
|---|---|---|---|---|
| Endocrine system | ko04914 | Progesterone-mediated oocyte maturation | 46/1 | 0.0005081 |
| ko04910 | Insulin signaling pathway | 66/0 | 0.0094483 | |
| ko04919 | Thyroid hormone signaling pathway | 49/2 | 0.0277375 | |
| ko04928 | Parathyroid hormone synthesis, secretion and action | 51/0 | 0.0277375 | |
| ko04912 | GnRH signaling pathway | 43/1 | 0.0298513 | |
| ko04922 | Glucagon signaling pathway | 42/0 | 0.0490603 | |
| Immune system | ko04664 | Fc epsilon RI signaling pathway | 34/2 | 0.0034951 |
| ko04624 | Toll and Imd signaling pathway | 21/1 | 0.0223963 | |
| ko04657 | IL-17 signaling pathway | 63/3 | 0.0279501 | |
| ko04662 | B cell receptor signaling pathway | 32/1 | 0.0402123 | |
| ko04650 | Natural killer cell mediated cytotoxicity | 41/1 | 0.0442341 | |
| Sensory system | ko04750 | Inflammatory mediator regulation of TRP channels | 42/3 | 0.0101365 |
| Category | Gene Name | Gene Description | log2 (XJ/CJ) | p Value |
|---|---|---|---|---|
| Genes related to endocrine system | GNRH1 | gonadotropin releasing hormone 1 | −2.1928 | 7.83 × 10−12 |
| ITPR1 | inositol 1,4,5-trisphosphate receptor type 1 | −1.1483 | 2.95 × 10−132 | |
| PLCB1 | phospholipase C beta 1 | −1.1312 | 5.16 × 10−11 | |
| PRKCB | protein kinase C beta | −1.2728 | 0 | |
| CAMK2A | calcium/calmodulin dependent protein kinase II alpha | −1.3253 | 2.46 × 10−49 | |
| CAMK2B | calcium/calmodulin dependent protein kinase II beta | −1.0355 | 6.43 × 10−4 | |
| CAMK2D | calcium/calmodulin dependent protein kinase II delta | −1.2762 | 5.04 × 10−106 | |
| FOXO1 | forkhead box O1 | −1.0255 | 4.15 × 10−114 | |
| CPT1B | carnitine palmitoyltransferase 1B | −1.0995 | 2.12 × 10−186 | |
| MAPK1 | mitogen-activated protein kinase 1 | −1.2105 | 0 | |
| MAPK7 | mitogen-activated protein kinase 7 | −1.1761 | 2.09 × 10−46 | |
| MAPK8 | mitogen-activated protein kinase 8 | −1.3110 | 1.74 × 10−31 | |
| MAPK9 | mitogen-activated protein kinase 9 | −1.1895 | 9.52 × 10−42 | |
| MAPK11 | mitogen-activated protein kinase 11 | −1.1115 | 1.46 × 10−149 | |
| MAPK13 | mitogen-activated protein kinase 13 | 1.2805 | 1.24 × 10−64 | |
| Genes associated with noise-induced hearing loss | FOXO4 | forkhead box O4 | −1.0029 | 1.75 × 10−148 |
| FOXO6 | forkhead box O6 | 2.1451 | 2.92 × 10−3 | |
| Catalase | RasGEF domain family member 1A | −1.2289 | 8.25 × 10−187 | |
| ATM | ATM serine/threonine kinase | −1.2877 | 3.19 × 10−81 | |
| GPX2 | glutathione peroxidase 2 | 1.2800 | 1.60 × 10−149 | |
| PTGS1 | prostaglandin-endoperoxide synthase 1 | −1.7015 | 0 | |
| PRDX2 | peroxiredoxin 2 | 1.2595 | 0 | |
| TLR2 | toll like receptor 2 | −1.0665 | 1.46 × 10−182 | |
| TLR4 | toll like receptor 4 | −1.3451 | 1.02 × 10−204 | |
| TNF | tumor necrosis factor | −1.4458 | 4.19 × 10−17 | |
| TNFAIP3 | TNF alpha induced protein 3 | −1.0094 | 5.16 × 10−186 | |
| TNFSF10 | TNF superfamily member 10 | −1.0813 | 9.92 × 10−127 | |
| TRPC1 | transient receptor potential cation channel subfamily C member 1 | −1.4755 | 1.36 × 10−9 | |
| TRPC3 | transient receptor potential cation channel subfamily C member 3 | −1.4399 | 1.28 × 10−3 | |
| TRPC4AP | transient receptor potential cation channel subfamily C member 4 associated protein | −1.0109 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Yin, D.; Lin, D.; Yan, Y.; Zhu, X.; Ying, C.; Zhang, J.; Xu, P.; Liu, K. Blood Transcriptome Analysis Reveals Gene Expression Differences between Yangtze Finless Porpoises from Two Habitats: Natural and Ex Situ Protected Waters. Fishes 2022, 7, 96. https://doi.org/10.3390/fishes7030096
Liu W, Yin D, Lin D, Yan Y, Zhu X, Ying C, Zhang J, Xu P, Liu K. Blood Transcriptome Analysis Reveals Gene Expression Differences between Yangtze Finless Porpoises from Two Habitats: Natural and Ex Situ Protected Waters. Fishes. 2022; 7(3):96. https://doi.org/10.3390/fishes7030096
Chicago/Turabian StyleLiu, Wang, Denghua Yin, Danqing Lin, Yan Yan, Xiaoyan Zhu, Congping Ying, Jialu Zhang, Pao Xu, and Kai Liu. 2022. "Blood Transcriptome Analysis Reveals Gene Expression Differences between Yangtze Finless Porpoises from Two Habitats: Natural and Ex Situ Protected Waters" Fishes 7, no. 3: 96. https://doi.org/10.3390/fishes7030096
APA StyleLiu, W., Yin, D., Lin, D., Yan, Y., Zhu, X., Ying, C., Zhang, J., Xu, P., & Liu, K. (2022). Blood Transcriptome Analysis Reveals Gene Expression Differences between Yangtze Finless Porpoises from Two Habitats: Natural and Ex Situ Protected Waters. Fishes, 7(3), 96. https://doi.org/10.3390/fishes7030096