
Cloning of Grass Carp Chemokine XC Receptor 1 (XCR1) Gene and Evaluation of Its Expression in Various Organs after GCRV Infection

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish and Virus Sources
2.2. Experimental Design and Sample Collection
2.3. RNA Extraction and cDNA Synthesis
2.4. Primer Design and Amplification and Sequencing of cDNA Fragment of Grass Carp XCR1 Gene
2.5. Sequence Analysis
2.6. qRT-PCR
2.7. Sequence Availability
2.8. Data Analysis
3. Results
3.1. cDNA Sequences of Grass Carp XCR1 Gene
3.2. Amino Acid Sequence of Grass Carp XCR1 Protein
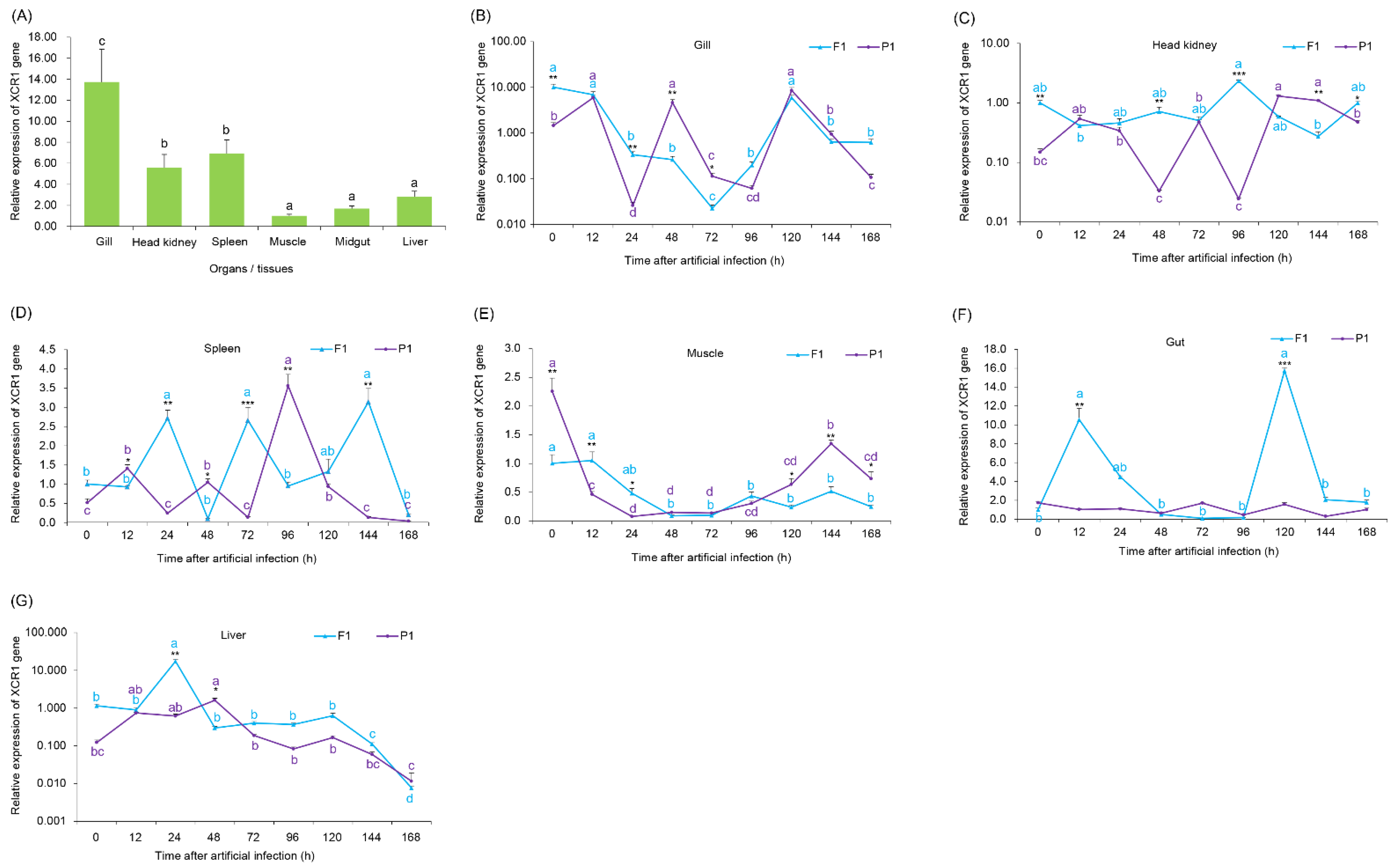
3.3. Expression of the XCR1 Gene in Various Grass Carp Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020. In Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Wang, H.; Ding, C.; Wang, J.; Zhao, X.; Jin, S.; Liang, J.; Luo, H.; Li, D.; Li, R.; Li, Y.; et al. Molecular cloning and expression analysis of coagulation factor VIII and plasminogen involved in immune response to GCRV, and immunity activity comparison of grass carp Ctenopharyngodon idella with different viral resistance. Fish Shellfish Immunol. 2019, 86, 794–804. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, H.; Li, R.; Li, Y.; Xiao, T. Different resistance potential to reovirus in grass carp (Ctenopharyngodon idella) populations and their immune characteristics. Aquac. Int. 2021, 29, 253–260. [Google Scholar] [CrossRef]
- Liang, H.-R.; Li, Y.-G.; Zeng, W.-W.; Wang, Y.-Y.; Wang, Q.; Wu, S.-Q. Pathogenicity and tissue distribution of grass carp reovirus after intraperitoneal administration. Virol. J. 2014, 11, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, W.; Wang, Y.; Liang, H.; Liu, C.; Song, X.; Shi, C.; Wu, S.; Wang, Q. A one-step duplex rRT-PCR assay for the simultaneous detection of grass carp reovirus genotypes I and II. J. Virol. Methods 2014, 210, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Zhong, L.; Liu, Q.; Xiao, T.; Su, J.; Chen, K.; Wang, H.; Dai, Y.; Chen, J. Characterization of grass carp spleen transcriptome during GCRV infection. Genet. Mol. Res. 2016, 15, gnr.15026650. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Lu, Y.; Xiao, X. Progress in the research on XCR I and its ligand. Int. J. Pathol. Clin. Med. 2013, 33, 57–61. [Google Scholar] [CrossRef]
- Nomiyama, H.; Hieshima, K.; Osada, N.; Kato-Unoki, Y.; Otsuka-Ono, K.; Takegawa, S.; Izawa, T.; Yoshizawa, A.; Kikuchi, Y.; Tanase, S.; et al. Extensive expansion and diversification of the chemokine gene family in zebrafish: Identification of a novel chemokine subfamily CX. BMC Genom. 2008, 9, 222. [Google Scholar] [CrossRef] [Green Version]
- Aghaallaei, N.; Bajoghli, B.; Schwarz, H.; Schorpp, M.; Boehm, T. Characterization of mononuclear phagocytic cells in medaka fish transgenic for a cxcr3a:gfp reporter. Proc. Natl. Acad. Sci. USA 2010, 107, 18079–18084. [Google Scholar] [CrossRef] [Green Version]
- Dorner, B.G.; Scheffold, A.; Rolph, M.S.; Hüser, M.B.; Kaufmann, S.H.E.; Radbruch, A.; Flesch, I.E.A.; Kroczek, R.A. MIP-1alpha, MIP-1beta, RANTES and ATAC/lymphotactin function together with IFN-gamma as Type 1 cytokines. Proc. Natl. Acad. Sci. USA 2002, 99, 6181–6186. [Google Scholar] [CrossRef] [Green Version]
- Dorner, B.G.; Dorner, M.B.; Zhou, X.; Opitz, C.; Mora, A.; Güttler, S.; Hutloff, A.; Mages, H.W.; Ranke, K.; Schaefer, M.; et al. Selective Expression of the Chemokine Receptor XCR1 on Cross-presenting Dendritic Cells Determines Cooperation with CD8+ T Cells. Immunity 2009, 31, 823–833. [Google Scholar] [CrossRef] [Green Version]
- Khurram, S.A.; Whawell, S.A.; Bingle, L.; Murdoch, C.; McCabe, B.M.; Farthing, P.M. Functional expression of the chemokine receptor XCR1 on oral epithelial cells. J. Pathol. 2010, 221, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Kroczek, R.A.; Henn, V. The Role of XCR1 and its Ligand XCL1 in Antigen Cross-Presentation by Murine and Human Dendritic Cells. Front. Immunol. 2012, 3, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, M.; Sun, B.; Nie, P. The first non-mammalian CXCR3 in a teleost fish: Gene and expression in blood cells and central nervous system in the grass carp (Ctenopharyngodon idella). Mol. Immunol. 2007, 44, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.Q.; Chang, M.X.; Sun, R.H.; Xiao, F.S.; Nie, P. The first non-mammalian CXCR5 in a teleost fish: Molecular cloning and expression analysis in grass carp (Ctenopharyngodon idella). BMC Immunol. 2010, 11, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Yang, M.; Li, C.; Wang, S.; Wang, Y.; Lin, F.; Zheng, L.; Yu, Y.; Qin, Q. Functional analysis of the CXCR1a gene response to SGIV viral infection in grouper. Fish Shellfish Immunol. 2019, 88, 217–224. [Google Scholar] [CrossRef] [PubMed]
- SWISS-MODEL Website. Available online: https://swissmodel.expasy.org/ (accessed on 1 January 2022).
- ProtParam Website. Available online: http://web.expasy.org/protparam/ (accessed on 1 January 2022).
- Conserved Domain Database. Available online: https://www.ncbi.nlm.nih.gov/cdd (accessed on 1 January 2022).
- Marchler-Bauer, A.; Lu, S.; Anderson, J.B.; Chitsaz, F.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H.; Geer, L.; Geer, R.C.; Gonzales, N.R.; et al. CDD: A Conserved Domain Database for the functional annotation of proteins. Nucleic Acids Res. 2011, 39, D225–D229. [Google Scholar] [CrossRef] [Green Version]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S.; Notes, A. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Weis, W.I.; Kobilka, B.K. Structural insights into G-protein-coupled receptor activation. Curr. Opin. Struct. Biol. 2008, 18, 734–740. [Google Scholar] [CrossRef] [Green Version]
- Kobilka, B.K. Structural insights into adrenergic receptor function and pharmacology. Trends Pharmacol. Sci. 2011, 32, 213–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, L.-Y.; Zhou, L.; Wang, H.-Q.; Luo, X.-C.; Dan, X.-M.; Li, Y.-W. Identification and expression analysis of three XCR1-like receptors from Epinephelus coioides after Cryptocaryon irritans infection. Fish Shellfish Immunol. 2017, 67, 95–102. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Jiang, W.-D.; Liu, X.-A.; Feng, L.; Wu, P.; Liu, Y.; Jiang, J.; Tan, B.-P.; Yang, Q.-H.; Kuang, S.-Y.; et al. Dietary biotin deficiency decreased growth performance and impaired the immune function of the head kidney, spleen and skin in on-growing grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2020, 97, 216–234. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, S.; Balasch, J.C.; Novoa, B.; Ribas, L.; Roher, N.; Krasnov, A.; Figueras, A. Comparative analysis of the acute response of the trout, O. mykiss, head kidney to in vivo challenge with virulent and attenuated infectious hematopoietic necrosis virus and LPS-induced inflammation. BMC Genom. 2008, 9, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, N.; Song, Y.-L.; Wang, B.; Zhang, X.-Y.; Zhang, X.-J.; Wang, Y.-L.; Cheng, Y.-Y.; Chen, D.-D.; Xia, X.-Q.; Lu, Y.-S.; et al. Fish gut-liver immunity during homeostasis or inflammation revealed by integrative transcriptome and proteome studies. Sci. Rep. 2016, 6, 36048. [Google Scholar] [CrossRef] [Green Version]
- Crozat, K.; Guiton, R.; Contreras, V.; Feuillet, V.; Dutertre, C.-A.; Ventre, E.; Manh, T.-P.V.; Baranek, T.; Storset, A.K.; Marvel, J.; et al. The XC chemokine receptor 1 is a conserved selective marker of mammalian cells homologous to mouse CD8α+ dendritic cells. J. Exp. Med. 2010, 207, 1283–1292. [Google Scholar] [CrossRef]
- Zhu, C.; Ou, Z. Progress in the research on lymphotactin (Ltn). Foreign Med. Sci. Sec. Pathophysiol. Clin. Med. 2002, 22, 320–322. [Google Scholar]
- Lei, Y.; Ripen, A.M.; Ishimaru, N.; Ohigashi, I.; Nagasawa, T.; Jeker, L.; Bösl, M.R.; Holländer, G.A.; Hayashi, Y.; Malefyt, R.D.W.; et al. Aire-dependent production of XCL1 mediates medullary accumulation of thymic dendritic cells and contributes to regulatory T cell development. J. Exp. Med. 2011, 208, 383–394. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | References |
|---|---|---|
| XCR1 3′-1 | 5′-TCAGTCTTTTCGCTGCCGTCCCACAA-3′ | This study |
| XCR1 3′-2 | 5′-CTTTTTGCTACACTGTGATCCTGGC-3′ | This study |
| XCR1 5′-1 | 5′-GCCCCCATCCCAAGAAAAAAAACACC-3′ | This study |
| XCR1 5′-2 | 5′-GCCGTGCCAGGATCACAGTGTAGCAA-3′ | This study |
| XCR1-PF | 5′-TGCTGCCTGAATCCTGTGT-3′ | This study |
| XCR1-PR | 5′-CGCCGTTGGAGTAAATAAGC-3′ | This study |
| β-actin Primer 1 | 5′-GGCTGTGCTGTCCCTGTATG-3 | [2] |
| β-actin Primer 2 | 5′-CTCTGGGCACCTGAACCTCT-3′ | [2] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, H.; Liu, Y.; Xiao, T.; Wang, H.; Xu, B. Cloning of Grass Carp Chemokine XC Receptor 1 (XCR1) Gene and Evaluation of Its Expression in Various Organs after GCRV Infection. Fishes 2022, 7, 130. https://doi.org/10.3390/fishes7030130
Yu H, Liu Y, Xiao T, Wang H, Xu B. Cloning of Grass Carp Chemokine XC Receptor 1 (XCR1) Gene and Evaluation of Its Expression in Various Organs after GCRV Infection. Fishes. 2022; 7(3):130. https://doi.org/10.3390/fishes7030130
Chicago/Turabian StyleYu, Hailuo, Yi Liu, Tiaoyi Xiao, Hongquan Wang, and Baohong Xu. 2022. "Cloning of Grass Carp Chemokine XC Receptor 1 (XCR1) Gene and Evaluation of Its Expression in Various Organs after GCRV Infection" Fishes 7, no. 3: 130. https://doi.org/10.3390/fishes7030130
APA StyleYu, H., Liu, Y., Xiao, T., Wang, H., & Xu, B. (2022). Cloning of Grass Carp Chemokine XC Receptor 1 (XCR1) Gene and Evaluation of Its Expression in Various Organs after GCRV Infection. Fishes, 7(3), 130. https://doi.org/10.3390/fishes7030130