Sturgeon Meat and Caviar Quality from Different Cultured Species

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Results
2.1. Proximate and Fatty Acid Composition of Eggs and Caviar
2.2. Fatty acid Composition of Lipid Fractions
2.3. Proximate Composition and Fatty acid Composition of Sturgeon Fillets
2.4. Color Parameters of Sturgeon Fillets
3. Materials and Methods
3.1. Samples Collection
3.2. Proximate Composition and Fatty Acid Analysis
3.3. Lipid Fractions Separation
3.4. Color Measurements in Fillets
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Ethical Approval
References
- Bronzi, P.; Chebanov, M.; Michaels, J.T.; Wei, Q.; Rosenthal, H.; Gessner, J. Sturgeon meat and caviar production: Global update 2017. J. Appl. Ichthyol. 2019, 35, 257–266. [Google Scholar] [CrossRef]
- Chebanov, M.; Billard, R. The culture of sturgeons in Russia: Production of juveniles for stocking and meat for human consumption. Aquat. Living Resour. 2001, 14, 375–381. [Google Scholar] [CrossRef]
- Bronzi, P.; Rosenthal, H.; Gessner, J. Global sturgeon aquaculture production: An overview. J. Appl. Ichthyol. 2011, 27, 169–175. [Google Scholar] [CrossRef]
- Bronzi, P.; Rosenthal, H. Present and future sturgeon and caviar production and marketing: A global market overview. J. Appl. Ichthyol. 2014, 30, 1536–1546. [Google Scholar] [CrossRef]
- CITES. CITES Trades Database for the Year 2018. Available online: https://trade.cites.org/ (accessed on 27 September 2019).
- Parisi, G.; Terova, G.; Gasco, L.; Piccolo, G.; Roncarati, A.; Moretti, V.M.; Centoducati, G.; Gatta, P.P.; Pais, A. Current status and future perspectives of Italian finfish aquaculture. Rev. Fish Biol. Fish. 2014, 24, 15–73. [Google Scholar] [CrossRef]
- Caprino, F.; Moretti, V.M.; Bellagamba, F.; Turchini, G.M.; Busetto, M.L.; Giani, I.; Paleari, M.A.; Pazzaglia, M. Fatty acid composition and volatile compounds of caviar from farmed white sturgeon (Acipenser transmontanus). Anal. Chim. Acta 2008, 617, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Bledsoe, G.E.; Bledsoe, C.D.; Rasco, B. Caviars and Fish Roe Products. Crit. Rev. Food Sci. Nutr. 2003, 43, 317–356. [Google Scholar] [CrossRef]
- Ovissipour, M.; Rasco, B. Fatty acid and amino acid profiles of domestic and wild beluga (Huso huso) roe and impact on fertilization ratio. J. Aquac. Res. Dev. 2011, 2. [Google Scholar] [CrossRef]
- DePeters, E.J.; Puschner, B.; Taylor, S.J.; Rodzen, J.A. Can fatty acid and mineral compositions of sturgeon eggs distinguish between farm-raised versus wild white (Acipenser transmontanus) sturgeon origins in California? Preliminary report. Forensic Sci. Int. 2013, 229, 128–132. [Google Scholar] [CrossRef]
- Gessner, J.; Würtz, S.; Kirschbaum, F.; Wirth, M. Biochemical composition of caviar as a tool to discriminate between aquaculture and wild origin. J. Appl. Ichthyol. 2008, 24, 52–56. [Google Scholar] [CrossRef]
- Czesny, S.; Dabrowski, K.; Christensen, J.E.; Van Eenennaam, J.; Doroshov, S. Discrimination of wild and domestic origin of sturgeon ova based on lipids and fatty acid analysis. Aquaculture 2000, 189, 145–153. [Google Scholar] [CrossRef]
- Mol, S.; Turan, S. Comparison of proximate, fatty acid and amino acid compositions of various types of fish roes. Int. J. Food Prop. 2008, 11, 669–677. [Google Scholar] [CrossRef]
- Hamzeh, A.; Moslemi, M.; Karaminasab, M.; Khanlar, M.A.; Faizbakhsh, R.; Batebi Navai, M.; Tahergorabi, R. Amino acid composition of roe from wild and farmed beluga sturgeon (Huso huso). J. Agric. Sci. Technol. 2015, 17, 357–364. [Google Scholar]
- Badiani, A.; Stipa, S.; Nanni, N.; Gatta, P.P.; Manfredini, M. Physical indices, processing yields, compositional parameters and fatty acid profile of three species of cultured sturgeon (genus Acipenser). J. Sci. Food Agric. 1997, 74, 257–264. [Google Scholar] [CrossRef]
- Vaccaro, A.M.; Buffa, G.; Messina, C.M.; Santulli, A.; Mazzola, A. Fatty acid composition of a cultured sturgeon hybrid (Acipenser naccarii x A. baerii). Food Chem. 2005, 93, 627–631. [Google Scholar] [CrossRef]
- Paleari, M.A.; Beretta, G.; Grimaldi, P.; Vaini, F. Composition of muscle tissue of farmed white sturgeon (Acipenser transmontanus) with particular reference to lipidic content. J. Appl. Ichthyol. 1997, 13, 63–66. [Google Scholar] [CrossRef]
- Wirth, M.; Kirschbaum, F.; Gessner, J.; Krüger, A.; Patriche, N.; Billard, R. Chemical and biochemical composition of caviar from different sturgeon species and origins. Nahr. Food 2000, 44, 233–237. [Google Scholar] [CrossRef]
- Wirth, M.; Kirschbaum, F.; Gessner, J.; Williot, P.; Patriche, N.; Billard, R. Fatty acid composition in sturgeon caviar from different species: Comparing wild and farmed origins. Int. Rev. Hydrobiol. 2002, 87, 629–636. [Google Scholar] [CrossRef]
- Ovissipour, M.; Al-Qadiri, H.M.; Lu, X.; Hu, Y.; Ross, C.F.; Van Eenennaam, J.P.; Doroshov, S.I.; Rasco, B. The effect of white sturgeon (Acipenser transmontanus) ovarian fat deposition on caviar yield and nutritional quality: Introducing image processing method for sturgeon ovary fat determination. Int. Aquat. Res. 2015, 7, 263–272. [Google Scholar] [CrossRef][Green Version]
- Gussoni, M.; Greco, F.; Vezzoli, A.; Paleari, M.A.; Moretti, V.M.; Beretta, G.; Caprino, F.; Lanza, B.; Zetta, L. Monitoring the effects of storage in caviar from farmed Acipenser transmontanus using chemical, SEM, and NMR methods. J. Agric. Food Chem. 2006, 54, 6725–6732. [Google Scholar] [CrossRef]
- Shin, J.H.; Oliveira, A.C.M.; Rasco, B.A. Quality attributes and microbial storage stability of caviar from cultivated white sturgeon (Acipenser transmontanus). J. Food Sci. 2010, 75, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Henderson, R.J. Fatty acid metabolism in freshwater fish with particular reference to polyunsaturated fatty acids. Arch. Für Tierernaehr. 1996, 49, 5–22. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.-C.; Chapman, F.A.; Wei, C.-I.; Portier, K.M.; O’Keefe, S. Differentiation of Cultured and Wild Sturgeon (Acipenser oxyrinchus desotoi) Based on Fatty Acid Composition. J. Food Sci. 1995, 60, 631–635. [Google Scholar] [CrossRef]
- Das, U.N. Essential Fatty acids—A Review. Curr. Pharm. Biotechnol. 2006, 7, 467–482. [Google Scholar] [CrossRef] [PubMed]
- Yanes-Roca, C.; Rhody, N.; Nystrom, M.; Main, K.L. Effects of fatty acid composition and spawning season patterns on egg quality and larval survival in common snook (Centropomus undecimalis). Aquaculture 2009, 287, 335–340. [Google Scholar] [CrossRef]
- Bell, J.G.; Castell, J.D.; Tocher, D.R.; MacDonald, F.M.; Sargent, J.R. Effects of different dietary arachidonic acid : Docosahexaenoic acid ratios on phospholipid fatty acid compositions and prostaglandin production in juvenile turbot (Scophthalmus maximus). Fish Physiol. Biochem. 1995, 14, 139–151. [Google Scholar] [CrossRef]
- Bell, J.G.; Ashton, I.; Secombes, C.J.; Weitzel, B.R.; Dick, J.R.; Sargent, J.R. Dietary lipid affects phospholipid fatty acid compositions, eicosanoid production and immune function in Atlantic salmon (Salmo salar). Prostaglandins Leukot. Essent. Fat. Acids 1996, 54, 173–182. [Google Scholar] [CrossRef]
- Lee, S.M. Review of the lipid and essential fatty acid requirements of rockfish (Sebastes schlegeli). Aquac. Res. 2001, 32, 8–17. [Google Scholar] [CrossRef]
- Mukhopadhyay, T.; Ghosh, S. Lipid profile and fatty acid composition of two silurid fish eggs. J. Oleo Sci. 2007, 56, 399–403. [Google Scholar] [CrossRef]
- Prabhakara Rao, P.G.; Balaswamy, K.; Narsing Rao, G.; Jyothirmayi, T.; Karuna, M.S.L.; Prasad, R.B.N. Lipid classes, fatty acid and phospholipid composition of roe lipids from Catla catla and Cirrhinus mrigala. Int. Food Res. J. 2013, 20, 275–279. [Google Scholar]
- Tocher, D.R.; Sargent, J.R. Analyses of lipids and fatty acids in ripe roes of some Northwest European marine fish. Lipids 1984, 19, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Bransden, M.P.; Battaglene, S.C.; Goldsmid, R.M.; Dunstan, G.A.; Nichols, P.D. Broodstock condition, egg morphology and lipid content and composition during the spawning season of captive striped trumpeter, Latris lineata. Aquaculture 2007, 268, 2–12. [Google Scholar] [CrossRef]
- Tocher, D.R. Fatty acid requirements in ontogeny of marine and freshwater fish. Aquac. Res. 2010, 41, 717–732. [Google Scholar] [CrossRef]
- Bruce, M.; Oyen, F.; Bell, G.; Asturiano, J.F.; Farndale, B.; Carrillo, M.; Zanuy, S.; Ramos, J.; Bromage, N. Development of broodstock diets for the European Sea Bass (Dicentrarchus labrax) with special emphasis on the importance of n - 3 and n - 6 highly unsaturated fatty acid to reproductive performance. Aquaculture 1999, 177, 85–97. [Google Scholar] [CrossRef]
- Mazorra, C.; Bruce, M.; Bell, J.G.; Davie, A.; Alorend, E.; Jordan, N.; Rees, J.; Papanikos, N.; Porter, M.; Bromage, N. Dietary lipid enhancement of broodstock reproductive performance and egg and larval quality in Atlantic halibut (Hippoglossus hippoglossus). Aquaculture 2003, 227, 21–33. [Google Scholar] [CrossRef]
- Badiani, A.; Anfossi, P.; Fiorentini, L.; Gatta, P.P.; Manfredini, M.; Nanni, N.; Stipa, S.; Tolomelli, B. Nutritional composition of cultured sturgeon (Acipenser spp.). J. Food Compos. Anal. 1996, 9, 171–190. [Google Scholar] [CrossRef]
- Stansby, M.E. Chemical Characteristics of Fish Caught in the Northeast Pacific Ocean. Mar. Fish. Rev. 1976, 38, 1–11. [Google Scholar]
- Gatlin, D.M.; Barrows, F.T.; Brown, P.; Dabrowski, K.; Gaylord, T.G.; Hardy, R.W.; Herman, E.; Hu, G.; Krogdahl, Å.; Nelson, R.; et al. Expanding the utilization of sustainable plant products in aquafeeds: A review. Aquac. Res. 2007, 38, 551–579. [Google Scholar] [CrossRef]
- Palmegiano, G.B.; Agradi, E.; Forneris, G.; Gai, F.; Gasco, L.; Rigamonti, E.; Sicuro, B.; Zoccarato, I. Spirulina as a nutrient source in diets for growing sturgeon (Acipenser baeri). Aquac. Res. 2005, 36, 188–195. [Google Scholar] [CrossRef]
- Ünal Şengör, G.F.; Alakavuk, D.Ü.; Tosun, Ş.Y.; Ulusoy, Ş. The chemical and sensory quality of smoked sturgeon (Huso huso): A case study. J. Aquat. Food Prod. Technol. 2010, 19, 310–317. [Google Scholar] [CrossRef]
- Wedekind, H. Chemical composition and processability of farmed sturgeon hybrids with special emphasis on Bester. Int. Rev. Hydrobiol. 2002, 87, 621–627. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists; Association of Official Analytica Chemists: Arlington, VA, USA, 1996. [Google Scholar]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Christie, W.W. Preparation of derivatives of fatty acids. In Lipid Analysis: Isolation, Separation and Structural Analysis of Lipids; The Oily Press: Bridgwater, UK, 2003; pp. 205–225. [Google Scholar]
- Kaluzny, M.A.; Duncan, L.A.; Merrit, M.V.; Epps, D.E. Rapid separation of lipid classes in high yield and purity using bonded phase columns. J. Lipid Res. 1985, 26, 135–140. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| Eggs | Caviar | |||||
|---|---|---|---|---|---|---|
| Sturgeon Species | Siberian (A. baerii) | Russian (A. gueldenstaedtii) | White (A. transmontanus) | Siberian (A. baerii) | Russian (A. gueldenstaedtii) | White (A. transmontanus) |
| n | 5 | 5 | 5 | 8 | 12 | 5 |
| Chemical composition | ||||||
| Moisture g/100 g | 59.5 ± 0.9 C | 53.9 ± 2.0 A | 57.1 ± 1.3 B | 57.3 ± 2.5 B | 52.7 ± 2.2 A | 54.6 ± 0.9 A,B |
| Protein g/100 g | 23.8 ± 0.8 | 24.1 ± 1.0 | 24.9 ± 0.6 | 23.9 ± 2.2 | 24.7 ± 1.2 | 24.0 ± 1.3 |
| Lipid g/100 g | 14.9 ± 0.9 A | 19.7 ± 1.8 B | 16.1 ± 1.2 A | 14.9 ± 0.7 A | 19.1 ± 3.2 B | 17.7 ± 1.2 A |
| Ash g/100 g | 1.9 ± 0.8 | 2.3 ± 0.6 | 1.9 ± 0.1 | 3.9 ± 0.3 | 3.5 ± 0.8 | 3.7 ± 0.3 |
| Eggs | Caviar | |||||||
|---|---|---|---|---|---|---|---|---|
| Sturgeon Species | Siberian A. baerii | Russian A. gueldenstaedtii | White A. transmontanus | Siberian A. baerii | Russian A. gueldenstaedtii | White A. transmontanus | ||
| n | 5 | 5 | 5 | 8 | 11 | 12 | ||
| Fatty acids (g/100 g) | ||||||||
| 14:0 | 0.9 ± 0.1 B | 1.3 ± 0.1 A | 1.4 ± 0.1 A | ** | 0.9 ± 0.1 C | 1.2 ± 0.1 B | 1.4 ± 0.2 A | ** |
| 16:0 | 18.7 ± 3.3 | 16.5 ± 0.6 | 17.4 ± 0.6 | 17.5 ± 0.7 | 17.0 ± 1.2 | 16.6 ± 0.8 | ||
| 18:0 | 3.3 ± 0.6 | 3.3 ± 0.3 | 3.1 ± 0.2 | 3.2 ± 0.1 A,B | 3.4 ± 0.4 B | 3.0 ± 0.3 A | * | |
| ΣSFA | 22.9 ± 4.0 | 21.0 ± 0.6 | 21.9 ± 0.7 | 21.5 ± 0.8 | 21.6 ± 1.0 | 21.1 ± 0.8 | ||
| 16:1n7 | 3.6 ± 0.8 | 3.5 ± 0.2 | 3.3 ± 0.3 | 3.5 ± 0.3 A,B | 3.6 ± 0.4 B | 3.3 ± 0.3 A | * | |
| 18:1n9 | 24.9 ± 11.6 | 24.6 ± 0.9 | 29.4 ± 1.4 | 28.4 ± 2.5 C | 25.4 ± 1.1 B | 30.6 ± 1.3 A | ** | |
| 18:1n7 | 2.3 ± 1.2 | 2.3 ± 1.3 | 3.0 ± 0.2 | 3.0 ± 0.2 B | 3.0 ± 0.1 B | 3.2 ± 0.2 A | ** | |
| 20:1n9 | 1.5 ± 0.4 A | 0.8 ± 0.1 B | 1.3 ± 0.1 A | ** | 1.3 ± 0.2 A | 1.0 ± 0.2 B | 1.4 ± 0.2 A | ** |
| ΣMUFA | 32.3 ± 11.7 | 31.3 ± 1.6 | 36.9 ± 1.1 | 36.4 ± 2.4 C | 33.2 ± 1.4 B | 38.7 ± 1.3 A | ** | |
| 18:2n6 | 13.4 ± 2.3 A | 20.6 ± 0.6 B | 14.2 ± 0.8 A | ** | 12.8 ± 1.0 A | 16.8 ± 3.9 B | 13.1 ± 1.4 A | ** |
| 18:3n6 | 1.5 ± 0.1 B | 0.9 ± 0.2 A | 0.8 ± 0.1 A | ** | 1.2 ± 0.2 B | 0.6 ± 0.5 A | 0.7 ± 0.6 A,B | * |
| 18:3n3 | 2.0 ± 0.3 | 2.1 ± 0.1 | 1.8 ± 0.1 | 1.8 ± 0.2 | 1.8 ± 0.3 | 1.8 ± 0.2 | ||
| 20:2n6 | 0.7 ± 0.1 | 0.7 ± 0.0 | 0.8 ± 0.1 | 0.7 ± 0.1 | 0.6 ± 0.1 | 0.7 ± 0.1 | ||
| 20:3n6 | 0.5 ± 0.1 | 0.4 ± 0.1 | 0.5 ± 0.1 | 0.4 ± 0.1 | 0.2 ± 0.2 | 0.3 ± 0.2 | ||
| 20:4n6 | 2.3 ± 0.3 A | 1.7 ± 0.2 B | 2.3 ± 0.2 A | ** | 2.1 ± 0.2 C | 1.8 ± 0.2 B | 2.4 ± 0.2 A | ** |
| 20:3n3 | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.2 ± 0.1 | 0.2 ± 0.0 | 0.1 ± 0.0 | 0.2 ± 0.0 | ||
| 20:5n3 | 5.6 ± 1.3 A B | 6.5 ± 0.6 B | 4.7 ± 0.7 A | * | 5.4 ± 1.2 A | 6.5 ± 0.6 B | 5.0 ± 0.6 A | ** |
| 22:5n6 | 0.2 ± 0.0 B | 0.2 ± 0.0 B | 0.3 ± 0.0 A | * | 0.2 ± 0.0 B | 0.1 ± 0.1 A | 0.1 ± 0.1 A,B | * |
| 22:5n3 | 1.4 ± 0.4 B | 1.9 ± 0.1 A | 1.6 ± 0.3 A B | * | 1.5 ± 0.4 A | 1.9 ± 0.2 B | 1.6 ± 0.2 A | ** |
| 22:6n3 | 15.7 ± 3.2 B | 11.2 ± 0.3 A | 12.9 ± 1.0 A B | * | 14.0 ± 0.7 | 13.0 ± 1.6 | 12.6 ± 1.0 | |
| ΣPUFA | 44.8 ± 7.8 | 47.7 ± 1.2 | 41.2 ± 1.6 | 41.6 ± 2.0 A | 44.9 ± 2.4 B | 39.9 ± 1.2 A | ** | |
| Σn3 | 26.1 ± 5.2 | 23.0 ± 0.9 | 22.1 ± 1.4 | 23.9 ± 1.9 B | 24.4 ± 1.4 B | 22.2 ± 1.0 A | ** | |
| Σn6 | 18.5 ± 2.8 A | 24.4 ± 0.7 B | 18.8 ± 0.4 A | 17.4 ± 0.8 A | 20.1 ± 3.2 B | 17.2 ± 0.7 A | ** | |
| n3/n6 | 1.4 ± 0.1 A | 0.9 ± 0.0 B | 1.2 ± 0.1 C | 1.4 ± 0.1 | 1.3 ± 0.3 | 1.3 ± 0.1 | ||
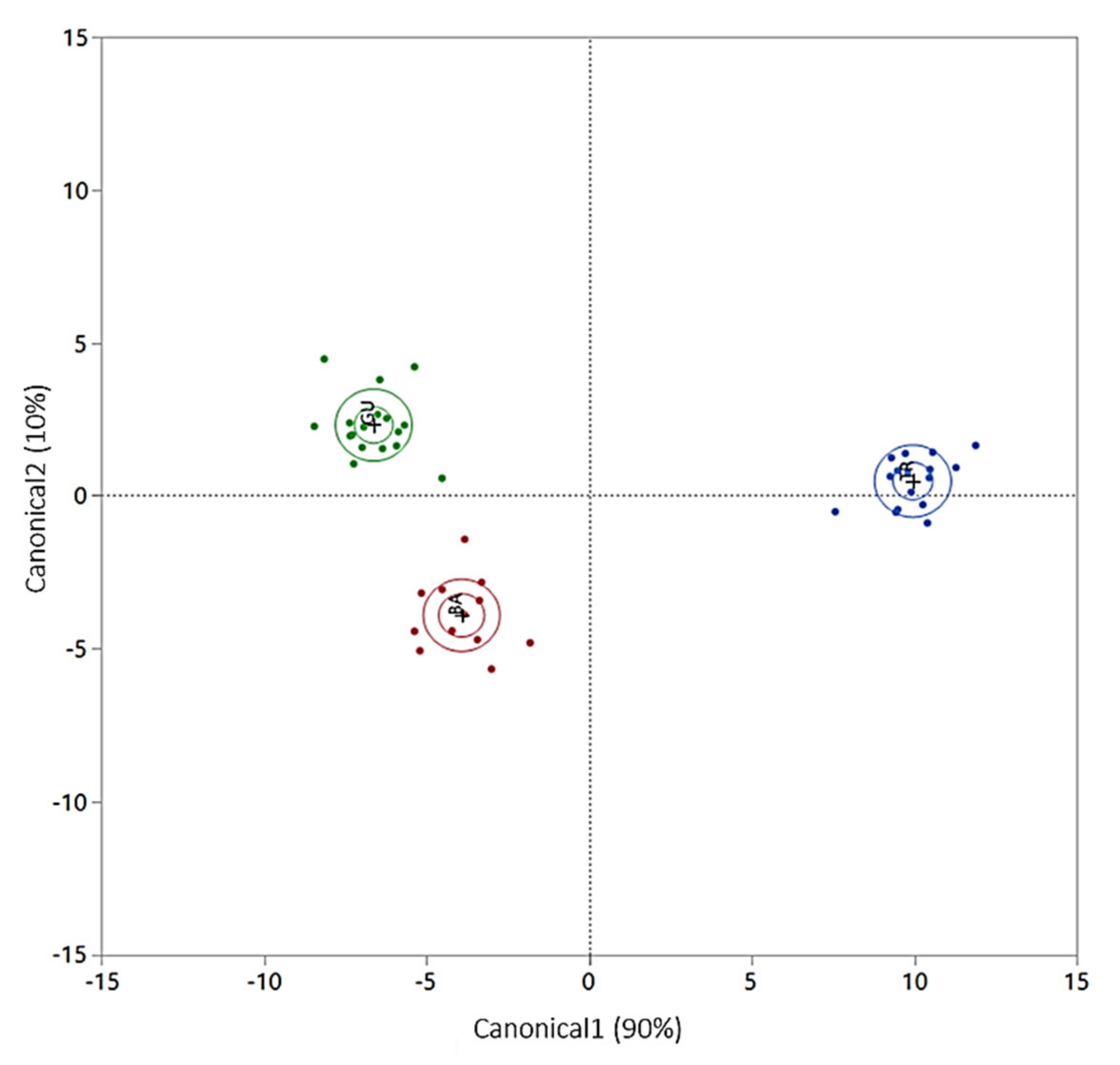
| Eggs and Caviar | ||||
|---|---|---|---|---|
| Species | Siberian A. baerii | Russian A. gueldenstaedtii | White A. transmontanus | Sign. |
| n | 13 | 16 | 17 | |
| C1 | −3.9 ± 1.0 A | −6.6 ± 0.1 B | 10.0 ± 1.0 C | ** |
| C2 | −3.9 ± 1.2 A | 2.3 ± 1.0 B | 0.48 ± 0.8 C | ** |
| Eggs | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Sturgeon Species | Siberian A. baerii | Russian A. gueldenstaedtii | White A. transmontanus | ||||||
| n | 5 | 5 | 5 | ||||||
| Fraction | PL | NL | PL | NL | PL | NL | |||
| Fatty acids (g/100 g) | Sign. | Sign. | Sign. | ||||||
| 14:0 | 2.1 ± 0.5 | 1.3 ± 0.1 | 2.3 ± 0.5 | 1.7 ± 0.2 | 2.1 ± 0.2 | 1.9 ± 0.3 | |||
| 16:0 | 22.2 ± 10.3 | 17.5 ± 1.5 | 21.4 ± 2.3 | 16.4 ± 3.0 | 22.3 ± 1.4 | 17.5 ± 1.5 | * | ||
| 18:0 | 8.6 ± 3.2 | 2.2 ± 0.3 | * | 9.3 ± 0.7 | 2.6 ± 0.7 | ** | 7.5 ± 0.4 | 2.5 ± 0.3 | ** |
| ΣSFA | 32.9 ± 15.1 | 21.1 ± 2.0 | 33.0 ± 5.3 | 20.7 ± 3.9 | * | 31.9 ± 3.1 | 21.8 ± 2.0 | ** | |
| 16:1n7 | 1.8 ± 0.8 | 4.1 ± 0.3 | ** | 1.9 ± 0.3 | 4.1 ± 0.8 | ** | 2.0 ± 0.2 | 3.8 ± 0.4 | ** |
| 18:1n9 | 15.8 ± 8.9 | 32.4 ± 3.5 | * | 14.8 ± 1.7 | 25.7 ± 4.8 | * | 18.4 ± 1.6 | 31.8 ± 2.6 | ** |
| 18:1n7 | 2.2 ± 1.1 | 2.9 ± 0.3 | 2.4 ± 0.2 | 2.9 ± 0.6 | 2.2 ± 0.3 | 2.9 ± 0.4 | |||
| 20:1n9 | 2.7 ± 1.4 | 1.2 ± 0.2 | 2.0 ± 0.5 | 0.8 ± 0.1 | * | 2.8 ± 0.6 | 1.2 ± 0.1 | ** | |
| ΣMUFA | 22.6 ± 12.7 | 40.5 ± 4.2 | 21.1 ± 3.0 | 33.4 ± 6.4 | * | 25.4 ± 2.8 | 39.7 ± 3.3 | ** | |
| 18:2n6 | 4.0 ± 1.1 | 13.9 ± 0.9 | ** | 5.8 ± 0.5 | 22.0 ± 4.1 | ** | 4.6 ± 0.6 | 15.4 ± 2.1 | ** |
| 18:3n6 | 1.2 ± 0.2 | 1.7 ± 0.2 | * | 1.2 ± 0.1 | 1.1 ± 0.3 | 1.1 ± 0.2 | 1.0 ± 0.1 | ||
| 18:3n3 | 1.3 ± 0.2 | 2.2 ± 0.2 | ** | 1.3 ± 0.1 | 2.4 ± 0.5 | * | 1.2 ± 0.2 | 2.1 ± 0.3 | * |
| 20:2n6 | 1.7 ± 0.4 | 0.6 ± 0.1 | ** | 1.9 ± 0.1 | 0.7 ± 0.1 | ** | 1.7 ± 0.2 | 0.7 ± 0.1 | ** |
| 20:3n6 | 1.6 ± 0.4 | 0.6 ± 0.1 | * | 1.5 ± 0.1 | 0.5 ± 0.1 | ** | 1.6 ± 0.4 | 0.6 ± 0.1 | ** |
| 20:4n6 | 3.5 ± 1.2 | 1.6 ± 0.3 | 3.0 ± 0.5 | 1.4 ± 0.5 | * | 3.9 ± 0.5 | 1.8 ± 0.2 | ** | |
| 20:3n3 | 1.0 ± 0.1 | 0.3 ± 0.0 | ** | 1.1 ± 0.1 | 0.3 ± 0.0 | ** | 1.0 ± 0.2 | 0.3 ± 0.0 | ** |
| 20:5n3 | 8.4 ± 3.0 | 4.4 ± 0.8 | 10.8 ± 1.0 | 5.9 ± 1.8 | * | 6.8 ± 0.8 | 4.0 ± 0.8 | * | |
| 22:5n6 | 0.7 ± 0.4 | 0.2 ± 0.0 | 0.8 ± 0.4 | 0.2 ± 0.1 | 0.8 ± 0.1 | 0.2 ± 0.1 | ** | ||
| 22:5n3 | 1.4 ± 0.8 | 1.4 ± 0.2 | 2.0 ± 0.4 | 2.2 ± 0.4 | 1.2 ± 0.3 | 1.7 ± 0.4 | |||
| 22:6n3 | 19.7 ± 7.3 | 11.5 ± 1.4 | 16.6 ± 0.9 | 9.3 ± 2.0 | ** | 18.7 ± 1.1 | 10.7 ± 1.9 | ** | |
| ΣPUFA | 44.6 ± 14.8 | 38.5 ± 3.5 | 45.8 ± 2.2 | 45.9 ± 10.1 | 42.6 ± 3.3 | 38.5 ± 5.9 | |||
| Σ n3 | 32.4 ± 11.3 | 20.6 ± 2.6 | 32.3 ± 1.2 | 21.0 ± 4.8 | * | 29.5 ± 2.3 | 19.7 ± 3.4 | * | |
| Σ n6 | 12.7 ± 3.5 | 18.9 ± 1.4 | * | 14.6 ± 1.1 | 26.1 ± 5.1 | * | 13.7 ± 1.5 | 19.9 ± 2.4 | * |
| n3/n6 | 2.5 ± 0.2 | 1.1 ± 0.1 | ** | 2.2 ± 0.1 | 0.8 ± 0.0 | ** | 1.8 ± 0.7 | 1.0 ± 0.1 | ** |
| Flesh | ||||
|---|---|---|---|---|
| Sturgeon Species | Siberian (A. baerii) | White (A. transmontanus) | White (A. transmontanus) | |
| Farmed for | Caviar production | Caviar production | Meat production | |
| Gender | Female | Female | Male | |
| Average fish weight | 5–8 kg | 30–50 kg | 6–10 kg | |
| n | 5 | 5 | 5 | |
| Proximate composition | Sign. | |||
| Moisture | 75.5 ± 1.6 | 75.2 ± 3.3 | 77.7 ± 1.1 | |
| Ash | 1.3 ± 0.2 | 1.2 ± 0.3 | 1.1 ± 0.0 | |
| Lipid | 5.6 ± 1.7 | 3.9 ± 2.5 | 2.6 ± 0.8 | |
| Protein | 17.6 ± 0.5 A | 19.6 ± 0.8 B | 18.6 ± 0.5 A,B | ** |
| Flesh | ||||
|---|---|---|---|---|
| Sturgeon Species | Siberian (A. baerii) | White (A. transmontanus) | White (A. transmontanus) | |
| Farmed for | Caviar production | Caviar production | Meat production | |
| Gender | Female | Female | Male | |
| Average fish weight | 5–8 kg | 30–50 kg | 6–10 kg | |
| n | 5 | 5 | 5 | |
| Fatty acid (g/100 g) | Sign | |||
| 14:0 | 1.3 ± 0.1 | 1.9 ± 0.6 | 1.7 ± 1.3 | |
| 16:0 | 15.5 ± 0.7 | 17.6 ± 0.8 | 16.9 ± 1.9 | |
| 18:0 | 2.4 ± 0.5 A | 3.9 ± 1.4 A | 5.9 ± 1.2 B | ** |
| ΣSFA | 19.2 ± 0.6 A | 23.4 ± 1.6 B | 24.5 ± 3.3 B | ** |
| 16:1n7 | 3.3 ± 0.6 | 3.4 ± 1.1 | 2.9 ± 2.0 | |
| 18:1n9 | 37.5 ± 2.6 A | 34.0 ± 2.3 A | 25.0 ± 6.3 B | ** |
| 18:1n7 | 2.8 ± 0.1 | 3.1 ± 0.1 | 2.9 ± 0.3 | |
| 20:1n9 | 1.8 ± 0.1 A | 2.2 ± 0.5 A | 0.5 ± 1.1 B | ** |
| ΣMUFA | 45.5 ± 2.8 A | 42.7 ± 3.5 A | 31.3 ± 5.5 B | ** |
| 18:2n6 | 16.1 ± 0.3 | 11.9 ± 0.7 | 14.1 ± 5.1 | |
| 18:3n6 | 1.3 ± 0.4 A | 0.4 ± 0.1 B | 0.4 ± 0.2 B | ** |
| 18:3n3 | 2.7 ± 0.4 A | 1.6 ± 0.4 B | 1.9 ± 0.8 A B | * |
| 20:2n6 | 0.9 ± 0.2 | 0.7 ± 0.2 | 0.8 ± 0.3 | |
| 20:3n6 | 0.5 ± 0.2 A,B | 0.3 ± 0.0 A | 0.6 ± 0.2 B | * |
| 20:4n6 | 1.6 ± 0.4 A | 2.0 ± 0.8 A | 3.6 ± 0.9 B | ** |
| 20:3n3 | 0.2 ± 0.1 | 0.2 ± 0.0 | 0.2 ± 0.2 | |
| 20:5n3 | 3.9 ± 0.8 A | 5.7 ± 0.8 A,B | 8.6 ± 4.5 B | * |
| 22:5n3 | 0.9 ± 0.6 | 1.4 ± 0.3 | 1.8 ± 0.7 | |
| 22:6n3 | 7.3 ± 1.5 A | 9.7 ± 1.9 A,B | 12.3 ± 3.2 B | * |
| ΣPUFA | 35.3 ± 2.7 A | 33.9 ± 2.1 A | 44.2 ± 2.9 B | ** |
| Σ n3 | 15.0 ± 2.2 A | 18.5 ± 1.5 A,B | 24.7 ± 7.4 B | * |
| Σ n6 | 20.4 ± 1.0 | 15.4 ± 1.2 | 19.5 ± 5.0 | |
| n3/n6 | 0.7 ± 0.1 | 1.2 ± 0.1 | 1.4 ± 0.9 | |
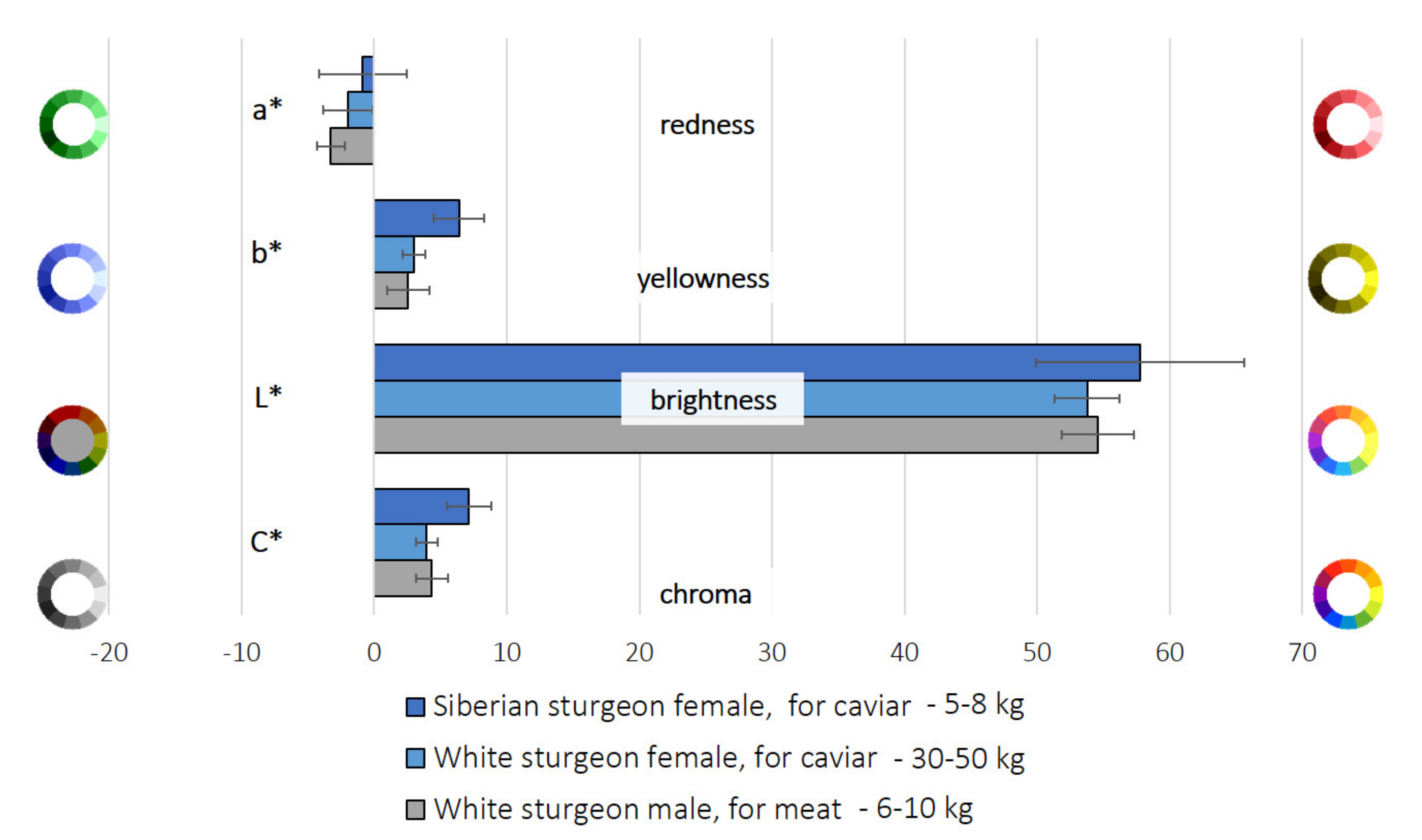
| Flesh | ||||
|---|---|---|---|---|
| Sturgeon Species | Siberian (A. baerii) | White (A. transmontanus) | White (A. transmontanus) | |
| Farmed for | Caviar production | Caviar production | Meat production | |
| Gender | Female | Female | Male | |
| Average fish weight | 5–8 kg | 30–50 kg | 6–10 kg | |
| Color parameters | Sign. | |||
| a* | −0.86 ± 3.29 | −1.98 ± 1.87 | −3.27 ± 1.04 | |
| b* | 6.41 ± 1.91 B | 2.99 ± 0.88 A | 2.56 ± 1.59 A | * |
| L* | 57.78 ± 7.85 | 53.76 ± 2.46 | 54.56 ± 2.70 | |
| C* | 7.15 ± 1.67 B | 3.97 ± 0.79 A | 4.36 ± 1.19 A | * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopez, A.; Vasconi, M.; Bellagamba, F.; Mentasti, T.; Moretti, V.M. Sturgeon Meat and Caviar Quality from Different Cultured Species. Fishes 2020, 5, 9. https://doi.org/10.3390/fishes5010009
Lopez A, Vasconi M, Bellagamba F, Mentasti T, Moretti VM. Sturgeon Meat and Caviar Quality from Different Cultured Species. Fishes. 2020; 5(1):9. https://doi.org/10.3390/fishes5010009
Chicago/Turabian StyleLopez, Annalaura, Mauro Vasconi, Federica Bellagamba, Tiziana Mentasti, and Vittorio Maria Moretti. 2020. "Sturgeon Meat and Caviar Quality from Different Cultured Species" Fishes 5, no. 1: 9. https://doi.org/10.3390/fishes5010009
APA StyleLopez, A., Vasconi, M., Bellagamba, F., Mentasti, T., & Moretti, V. M. (2020). Sturgeon Meat and Caviar Quality from Different Cultured Species. Fishes, 5(1), 9. https://doi.org/10.3390/fishes5010009