
Antibacterial Activity, Antioxidant Capacity and Immune Gene Modulation of Six Medicinal Plants in CHSE-214 Cells


Abstract

1. Introduction
2. Materials and Methods
2.1. Preparation of Plant Extracts
2.2. Antibacterial Activity Assay
2.3. Antioxidant Activity Assays
2.3.1. DPPH Radical Scavenging Assay
2.3.2. ABTS Radical Scavenging Assay
2.3.3. Total Polyphenol Content
2.4. Cell Culture
2.5. Cell Viability Assay and IC50 Determination
2.6. Immune Gene Expression Analysis
2.7. Statistical Analysis
3. Results
3.1. Antibacterial Activity
3.2. DPPH Radical Scavenging Activity
3.3. ABTS Radical Scavenging Activity
3.4. Total Polyphenol Content
3.5. Cell Viability on CHSE-214 Cells
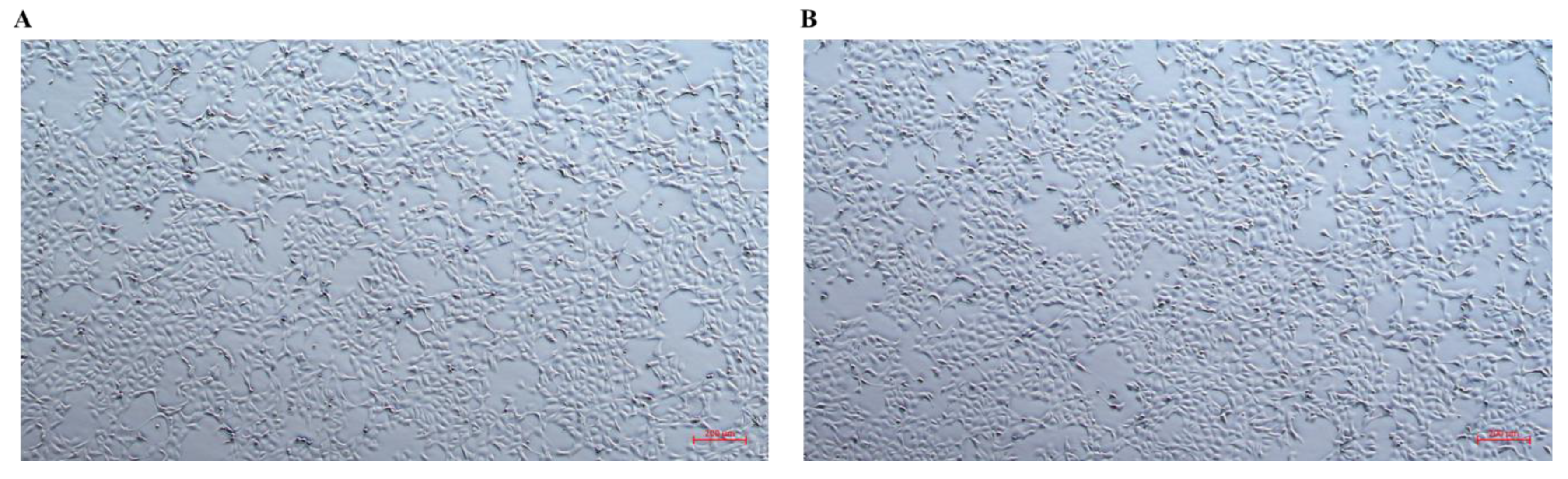
3.6. Morphological Changes in CHSE-214 Cells
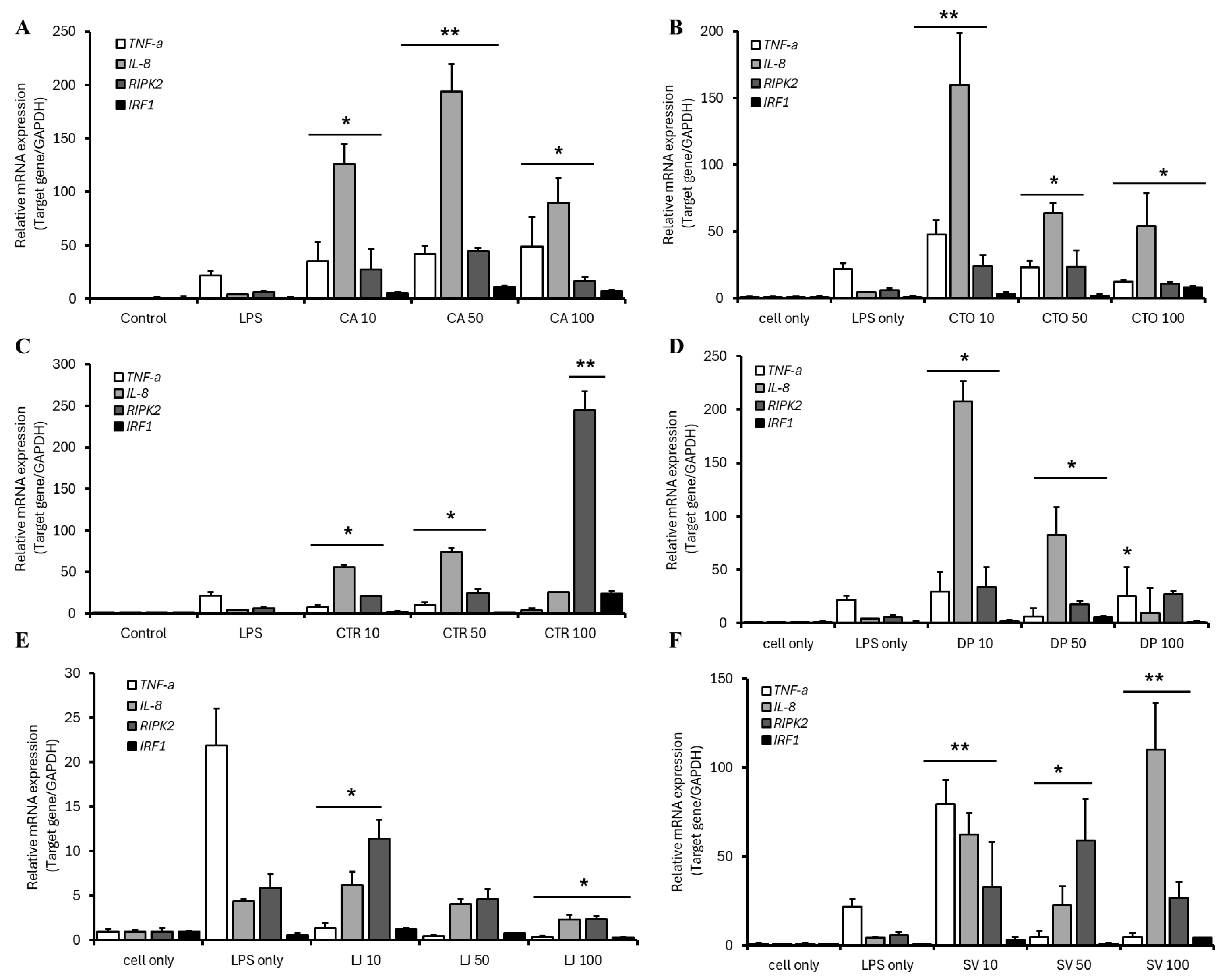
3.7. Relative mRNA Gene Expression in CHSE-214 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABTS | 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) |
| CA | Chenopodium album |
| CTO | Cassia tora |
| CTR | Cudrania tricuspidata |
| DP | Di-oscorea polystachya |
| DPPH | 2,2-Diphenyl-1-picrylhydrazyl |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| IC50 | Half-maximal inhibitory concentration |
| IL-8 | Interleukin-8 |
| IRF1 | Interferon regulatory factor 1 |
| LJ | Lonicera japonica |
| RIPK2 | Receptor-interacting serine/threonine-protein kinase 2 |
| MIC | Minimum inhibitory concentration |
| SV | Solidago virgaurea subsp. gigantea |
| TNF-α | Tumor necrosis factor-alpha |
| TPC | Total polyphenol content |
References
- Menanteau-Ledouble, S.; Kumar, G.; Saleh, M.; El-Matbouli, M. Aeromonas salmonicida: Updates on an old acquaintance. Dis. Aquat. Org. 2016, 120, 49–68. [Google Scholar] [CrossRef]
- Gouife, M.; Chen, S.; Huang, K.; Nawaz, M.; Jin, S.; Ma, R.; Xie, J. Photobacterium damselae subsp. damselae in mariculture. Aquac. Int. 2022, 30, 1453–1480. [Google Scholar] [CrossRef]
- Haines, A.N.; Gauthier, D.T.; Nebergall, E.E.; Cole, S.D.; Nguyen, K.M.; Rhodes, M.W.; Vogelbein, W.K. First report of Streptococcus parauberis in wild finfish from North America. Vet. Microbiol. 2013, 166, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Miranda, J.J.; Castillo-Pérez, L.J.; Ponce-Hernández, A.; Carranza-Álvarez, C. Summary of economic losses due to bacterial pathogens in aquaculture industry. In Bacterial Fish Diseases; Academic Press: Cambridge, MA, USA, 2022; pp. 399–417. [Google Scholar] [CrossRef]
- Ma, J.; Bruce, T.J.; Jones, E.M.; Cain, K.D. A review of fish vaccine development strategies: Conventional methods and modern biotechnological approaches. Microorganisms 2019, 7, 569. [Google Scholar] [CrossRef] [PubMed]
- Reverter, M.; Bontemps, N.; Lecchini, D.; Banaigs, B.; Sasal, P. Use of plant extracts in fish aquaculture as an alternative to chemotherapy: Current status and future perspectives. Aquaculture 2014, 433, 50–61. [Google Scholar] [CrossRef]
- Wang, W.; Sun, J.; Liu, C.; Xue, Z. Application of immunostimulants in aquaculture: Current knowledge and future perspectives. Aquac. Res. 2017, 48, 1–23. [Google Scholar] [CrossRef]
- Vijayaram, S.; Sun, Y.Z.; Zuorro, A.; Ghafarifarsani, H.; Van Doan, H.; Hoseinifar, S.H. Bioactive immunostimulants as health-promoting feed additives in aquaculture: A review. Fish Shellfish Immunol. 2022, 130, 294–308. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.; Soliman, A.A.; Gewaily, M.S.; Amer, A.A.; Shukry, M.; Khalil, R.H.; Shehata, A.I. Dietary effects of Saccharomyces cerevisiae and Allium sativum on growth, antioxidant status, hepatic and intestinal histoarchitecture, expression of growth- and immune-related genes, and resistance of Oreochromis niloticus to Aeromonas sobria. Fish Shellfish Immunol. 2024, 148, 109493. [Google Scholar] [CrossRef]
- Le Xuan, C.; Linh, N.V.; Wannavijit, S.; Outama, P.; Fontana, C.M.; Meepowpan, P.; Van Doan, H. Influences of makiang (Syzygium nervosum) seed powder on growth performance, immunological response, antioxidant and immune related gene expression in juvenile Nile tilapia (Oreochromis niloticus). Aquaculture 2024, 588, 740943. [Google Scholar] [CrossRef]
- Noorbakhsh, M.F.; Ghaemi, M.; Gholamhosseini, A.; Heidari, A.A. Effects of dietary supplement of basil extract on biochemical and immunological parameters and growth performance in Oncorhynchus mykiss. Aquac. Nutr. 2024, 2024, 5388049. [Google Scholar] [CrossRef]
- Reverter, M.; Tapissier-Bontemps, N.; Sarter, S.; Sasal, P.; Caruso, D. Moving towards more sustainable aquaculture practices: A meta-analysis on the potential of plant-enriched diets to improve fish growth, immunity and disease resistance. Rev. Aquac. 2021, 13, 537–555. [Google Scholar] [CrossRef]
- Khoobchandani, M.; Ojeswi, B.K.; Sharma, B.; Srivastava, M.M. Chenopodium album prevents progression of cell growth and enhances cell toxicity in human breast cancer cell lines. Oxid. Med. Cell. Longev. 2009, 2, 160–165. [Google Scholar] [CrossRef]
- Antonisamy, P.; Dhanasekaran, M.; Kim, H.R.; Jo, S.G.; Agastian, P.; Kwon, K.B. Anti-inflammatory and analgesic activity of ononitol monohydrate isolated from Cassia tora L. in animal models. Saudi J. Biol. Sci. 2017, 24, 1933–1938. [Google Scholar] [CrossRef]
- Kumar, R.S.; Narasingappa, R.B.; Joshi, C.G.; Girish, T.K.; Rao, U.J.P.; Danagoudar, A. Evaluation of Cassia tora Linn. against oxidative stress-induced DNA and cell membrane damage. J. Pharm. Bioallied Sci. 2017, 9, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Shin, G.R.; Lee, S.; Lee, S.; Do, S.G.; Shin, E.; Lee, C.H. Maturity stage-specific metabolite profiling of Cudrania tricuspidata and its correlation with antioxidant activity. Ind. Crops Prod. 2015, 70, 322–331. [Google Scholar] [CrossRef]
- Ko, W.; Kim, N.; Lee, H.; Woo, E.R.; Kim, Y.C.; Oh, H.; Lee, D.S. Anti-inflammatory effects of compounds from Cudrania tricuspidata in HaCaT human keratinocytes. Int. J. Mol. Sci. 2021, 22, 7472. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhao, S.; Tao, S.; Hou, G.; Zhao, F.; Tan, S.; Meng, Q. Dioscorea spp.: Bioactive compounds and potential for the treatment of inflammatory and metabolic diseases. Molecules 2023, 28, 2878. [Google Scholar] [CrossRef]
- Park, S.Y.; Truong, V.L.; Jeon, S.G.; Choe, S.Y.; Rarison, R.H.; Yoon, B.H.; Park, J.W.; Jeong, H.J.; Jeong, W.S. Anti-inflammatory and prebiotic potential of ethanol extracts and mucilage polysaccharides from Korean yams (Dioscorea polystachya and Dioscorea bulbifera). Foods 2025, 14, 173. [Google Scholar] [CrossRef]
- Tang, X.; Liu, X.; Zhong, J.; Fang, R. Potential application of Lonicera japonica extracts in animal production: From the perspective of intestinal health. Front. Microbiol. 2021, 12, 719877. [Google Scholar] [CrossRef]
- Jeong, S.H.; Park, M.Y.; Bhosale, P.B.; Abusaliya, A.; Won, C.K.; Park, K.I.; Kim, E.; Heo, J.D.; Kim, H.W.; Ahn, M.; et al. Potential antioxidant and anti-inflammatory effects of Lonicera japonica and Citri reticulatae pericarpium polyphenolic extract (LCPE). Antioxidants 2023, 12, 1582. [Google Scholar] [CrossRef]
- Malićanin, M.; Karabegović, I.; Đorđević, N.; Mančić, S.; Stojanović, S.S.; Brković, D.; Danilović, B. Influence of the extraction method on the biological potential of Solidago virgaurea L. essential oil and hydrolates. Plants 2024, 13, 2187. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.S.; Kim, H.Y.; Zuo, G.; Lee, E.H.; Kang, S.K.; Lim, S.S. Constituents from Solidago virgaurea var. gigantea and their inhibitory effect on lipid accumulation. Fitoterapia 2020, 146, 104683. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Zhao, J.; Chen, X.; Liu, M.; Ren, G.; Lu, T.; Shao, Y.; Xu, L. Autophagy induced by infectious pancreatic necrosis virus promotes its multiplication in the Chinook salmon embryo cell line CHSE-214. Fish Shellfish Immunol. 2020, 97, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xu, L.; Brandsma, J.H.; Wang, Y.; Hakim, M.S.; Zhou, X.; Pan, Q. Convergent transcription of interferon-stimulated genes by TNF-α and IFN-α augments antiviral activity against HCV and HEV. Sci. Rep. 2016, 6, 25482. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.Y.; Wang, K.L.; Li, L.; Li, B.; Wu, X.Y.; Zhang, Z.W.; Li, N.; Liu, L.H.; Nie, P.; Chen, S.N. Transcription of NOD1 and NOD2 and their interaction with CARD9 and RIPK2 in IFN signaling in a perciform fish, the Chinese perch, Siniperca chuatsi. Front. Immunol. 2024, 15, 1374368. [Google Scholar] [CrossRef]
- Tranquilino-Rodríguez, E.; Martínez-Flores, H.E. Ultrasound-assisted extraction of phenolic compounds from Moringa oleifera leaves by response surface methodology. Bioact. Compd. Health Dis. 2023, 6, 325–337. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 11th ed.; CLSI Standard M07; CLSI: Wayne, PA, USA, 2018. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Holen, E.; Austgulen, M.H.; Espe, M. RNA form baker’s yeast cultured with and without lipopolysaccharide (LPS) modulates gene transcription in an intestinal epithelial cell model, RTgutGC from rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2021, 119, 397–408. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhao, J.Z.; Xu, L.M.; Ren, G.M.; Shao, Y.Z.; Liu, Q.; Teng, C.B.; Lu, T.Y. Comparative transcriptome analysis of rainbow trout gonadal cells (RTG-2) infected with U and J genogroup infectious hematopoietic necrosis virus. Front. Microbiol. 2023, 13, 1109606. [Google Scholar] [CrossRef]
- Della Torre, C.; Monti, M.; Focardi, S.; Corsi, I. Time-dependent modulation of cyp1a gene transcription and EROD activity by musk xylene in PLHC-1 and RTG-2 fish cell lines. Toxicol. Vitr. 2011, 25, 1575–1580. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.W.; Kang, N.S.; Park, K.H. Isolation of antibacterial prenylated flavonoids from Cudrania tricuspidata. Appl. Biol. Chem. 2004, 47, 270–273. [Google Scholar]
- Chamkhi, I.; Charfi, S.; El Hachlafi, N.; Mechchate, H.; Guaouguaou, F.E.; El Omari, N.; Bakrim, S.; Balahbib, A.; Zengin, G.; Bouyahya, A. Genetic diversity, antimicrobial, nutritional, and phytochemical properties of Chenopodium album: A comprehensive review. Food Res. Int. 2022, 154, 110979. [Google Scholar] [CrossRef] [PubMed]
- Huo, L.C.; Liu, N.Y.; Wang, C.J.; Luo, Y.; Liu, J.X. Lonicera japonica protects Pelodiscus sinensis by inhibiting the biofilm formation of Aeromonas hydrophila. Appl. Microbiol. Biotechnol. 2024, 108, 67. [Google Scholar] [CrossRef] [PubMed]
- Cushnie, T.T.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Mukwaya, E.; Wong, M.S.; Zhang, Y. A systematic review on biological activities of prenylated flavonoids. Pharm. Biol. 2014, 52, 655–660. [Google Scholar] [CrossRef]
- Nazir, R.; Kumar, V.; Gupta, S.; Dwivedi, P.; Pandey, D.K.; Dey, A. Biotechnological strategies for the sustainable production of diosgenin from Dioscorea spp. Appl. Microbiol. Biotechnol. 2021, 105, 569–585. [Google Scholar] [CrossRef]
- Guan, R.; Guo, F.; Guo, R.; Wang, S.; Sun, X.; Zhao, Q.; Zhang, Y.; Lin, J. Integrated metabolic profiling and transcriptome analysis of Lonicera japonica flowers for chlorogenic acid, luteolin and endogenous hormone syntheses. Gene 2023, 888, 147739. [Google Scholar] [CrossRef]
- Kim, D.C.; Yoon, C.S.; Quang, T.H.; Ko, W.; Kim, J.S.; Oh, H.; Kim, Y.C. Prenylated flavonoids from Cudrania tricuspidata suppress lipopolysaccharide-induced neuroinflammatory activities in BV2 microglial cells. Int. J. Mol. Sci. 2016, 17, 255. [Google Scholar] [CrossRef]
- Lee, B.W.; Lee, J.H.; Lee, S.T.; Lee, H.S.; Lee, W.S.; Jeong, T.S.; Park, K.H. Antioxidant and cytotoxic activities of xanthones from Cudrania tricuspidata. Bioorg. Med. Chem. Lett. 2005, 15, 5548–5552. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Chen, S.H.; Chen, C.H.; Chou, P.Y.; Yang, C.C.; Lin, F.H. Polysaccharide extracted from Bletilla striata promotes proliferation and migration of human tenocytes. Polymers 2020, 12, 2567. [Google Scholar] [CrossRef]
- Budzianowski, J.; Thiem, B.; Kikowska, M. Solidago virgaurea L.—Chemical composition, traditional and medicinal use, pharmaceutical properties, potential applications, and biotechnological studies—A review. In Medicinal Plants; Ekiert, H.M., Ramawat, K.G., Arora, J., Eds.; Sustainable Development and Biodiversity; Springer: Cham, Switzerland, 2021; Volume 28, pp. 569–602. [Google Scholar] [CrossRef]
- Choi, S.Z.; Choi, S.U.; Lee, K.R. Pytochemical constituents of the aerial parts from Solidago virga-aurea var. gigantea. Arch. Pharm. Res. 2004, 27, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Chan, L.P.; Liu, C.; Chiang, F.Y.; Wang, L.F.; Lee, K.W.; Chen, W.T.; Liang, C.H. IL-8 promotes inflammatory mediators and stimulates activation of p38 MAPK/ERK-NF-κB pathway and reduction of JNK in HNSCC. Oncotarget 2017, 8, 56375. [Google Scholar] [CrossRef]
- Zhou, Q.; Sun, H.J.; Zhang, X.W. Total saponin fraction of Dioscorea nipponica makino improves gouty arthritis symptoms in rats via M1/M2 polarization of monocytes and macrophages mediated by arachidonic acid signaling. Chin. J. Integr. Med. 2023, 29, 1007–1017. [Google Scholar] [CrossRef]
- Gao, F.Y.; Pang, J.C.; Lu, M.X.; Yang, X.L.; Zhu, H.P.; Ke, X.L.; Liu, Z.G.; Cao, J.M.; Wang, M. Molecular characterization, expression and functional analysis of NOD1, NOD2 and NLRC3 in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2018, 73, 207–219. [Google Scholar] [CrossRef]
- Yuan, Y.; Wang, Z.; Jiang, C.; Wang, X.; Huang, L. Exploiting genes and functional diversity of chlorogenic acid and luteolin biosyntheses in Lonicera japonica and their substitutes. Gene 2014, 534, 408–416. [Google Scholar] [CrossRef]
- Lou, L.; Liu, Y.; Zhou, J.; Wei, Y.; Deng, J.; Dong, B.; Chai, L. Chlorogenic acid and luteolin synergistically inhibit the proliferation of interleukin-1 β-induced fibroblast-like synoviocytes through regulating the activation of NF-κB and JAK/STAT-signaling pathways. Immunopharmacol. Immunotoxicol. 2015, 37, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Feng, Z.; Wang, L.; Ma, X.; Wang, L.; Liu, K.; Geng, X.; Peng, C. Chlorogenic acid alleviates hepatic ischemia–reperfusion injury by inhibiting oxidative stress, inflammation, and mitochondria-mediated apoptosis in vivo and in vitro. Inflammation 2023, 46, 1061–1076. [Google Scholar] [CrossRef]
- Fahmideh, H.; Shapourian, H.; Moltafeti, R.; Tavakol, C.; Forghaniesfidvajani, R.; Zalpoor, H.; Nabi-Afjadi, M. The role of natural products as inhibitors of JAK/STAT signaling pathways in glioblastoma treatment. Oxid. Med. Cell. Longev. 2022, 1, 7838583. [Google Scholar] [CrossRef]
- Abdel-Aziz, M.S.; Shaheen, M.S.; El-Nekeety, A.A.; Abdel-Wahhab, M.A. Antioxidant and antibacterial activity of silver nanoparticles biosynthesized using Chenopodium murale leaf extract. J. Saudi Chem. Soc. 2014, 18, 356–363. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Common Name | Plant Part | Extraction Method | Solvent | Extraction Conditions * | Abbreviation | Yield (g) |
|---|---|---|---|---|---|---|---|
| Chenopodium album | Lamb’s quarters | Leaf | Maceration | 70% ethanol | 80 °C, 3 h, 1:50 (w/v) | CA | 2.21 |
| Cassiatora | Sickle senna | Seed | Maceration | 70% ethanol | 80 °C, 3 h, 1:50 (w/v) | CTO | 0.29 |
| Cudrania tricuspidata | Chinese mulberry | Root | Maceration | 70% ethanol | 80 °C, 3 h, 1:50 (w/v) | CTR | 3.25 |
| Dioscorea polystachya | Chinese yam | Tuber | Maceration | 70% ethanol | 80 °C, 3 h, 1:50 (w/v) | DP | 1.37 |
| Lonicera japonica | Japanese honeysuckle | Flower | Maceration | 70% ethanol | 80 °C, 3 h, 1:50 (w/v) | LJ | 1.22 |
| Solidago virgaurea subsp. gigantea | Giant goldenrod | Aerial parts | Maceration | 70% ethanol | 80 °C, 3 h, 1:50 (w/v) | SV | 2.87 |
| Gene | Accession Number | Sequence (5′−3′) | Efficiency (%) | Reference |
|---|---|---|---|---|
| TNF-α | NM_001124357.1 | F: GTCAGCATGGAAGACCGTCA | 98 | This study |
| R: CCGGCAATCTGCTTCAATGT | ||||
| IL-8 | NM_001140710.2 | F: GGCCCTCCTGACCATTACT | 102 | [31] |
| R: ATGAGTCTACCAATTCGTCTGC | ||||
| RIPK2 | LOC110526239 | F: CTGATGCAATGACAAGGGCTAC | 100 | [33] |
| R: TGCTGGGACTGGTGGAAG | ||||
| IRF1 | LOC110533376 | F: GGCTGGAGGATAAGATTGA | 102 | |
| R: GTGTAGTCTCGCCTTGTATGA | ||||
| GAPDH | LOC100135863 | F: TGTTCGTCATGGGAGTCAACGA | 105 | [34] |
| R: ATGCCGAAGTTGTCGTGGATGA |
| Bacteria | Aeromonas hydrophila | Aeromonas salmonicida | Aeromonas sobria | Photobacterium damselae subsp. damselae | Streptococcus parauberis | Vibrio anguillarum | |
|---|---|---|---|---|---|---|---|
| Extract | |||||||
| CA | ≤10 | ≤10 | ≤10 | ≤10 | 10,000 | ≤10 | |
| CTO | ≤10 | ≤10 | ≤10 | ≤10 | 10,000 | ≤10 | |
| CTR | ≤10 | ≤10 | ≤10 | ≤10 | 10,000 | 750 | |
| DP | ≤10 | ≤10 | ≤10 | ≤10 | 10,000 | 10,000 | |
| LJ | ≤10 | ≤10 | ≤10 | ≤10 | 10,000 | ≤10 | |
| SV | ≤10 | ≤10 | ≤10 | ≤10 | 10,000 | ≤10 | |
| Extract | DPPH (μg/mL) | ABTS (μg/mL) | TPC (mg GAE/g) |
|---|---|---|---|
| IC50 | IC50 | ||
| CA | 2378 ± 11.4 | 4637 ± 48.7 | 1287 ± 12.5 |
| CTO | 1324 ± 10.2 | 227 ± 12.3 | 1409 ± 10.5 |
| CTR | 1292 ± 17.7 | 712 ± 41.5 | 1498 ± 11.2 |
| DP | 5695 ± 42.3 | 5109 ± 63.3 | 1214 ± 18.3 |
| LJ | 1394 ± 21.7 | 315 ± 22.4 | 1428 ± 12.2 |
| SV | 1647 ± 28.6 | 811 ± 35.3 | 1330 ± 13.8 |
| Trolox | 61 | - | - |
| Extract | Concentration (μg/mL) | IC50 | |||||
|---|---|---|---|---|---|---|---|
| 0 | 10 | 50 | 100 | 200 | 1000 | ||
| CA | 100 ± 2.8 | 99.6 ± 3.2 | 95.3 ± 2.1 | 86.7 ± 4.1 * | 76.2 ± 0.2 * | 132.7 ± 1.1 | ND |
| CTO | 100 ± 3.2 | 96.6 ± 5.0 | 113.3 ± 9.1 | 98.5 ± 8.8 | 95.6 ± 1.5 | 86.1 ± 2.1 * | ND |
| CTR | 100 ± 2.5 | 97.5 ± 2.1 | 88.6 ± 5.3 * | 86.91 ± 3.5 * | 82.2 ± 5.3 * | 45.6 ± 3.0 ** | 904.2 ± 20.5 |
| DP | 100 ± 3.8 | 100.5 ± 5.9 | 105.7 ± 11.6 | 107.9 ± 3.9 | 116.0 ± 5.2 | 105.1 ± 10.9 | ND |
| LJ | 100 ± 2.7 | 97.3 ± 3.2 | 98.0 ± 2.9 | 98.3 ± 0.5 | 104.6 ± 4.5 | 97.6 ± 1.5 | ND |
| SV | 100 ± 3.5 | 109.9 ± 6.4 | 115.8 ± 3.8 | 118.0 ± 9.7 | 110.1 ± 1.4 | 58.3 ± 0.8 ** | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woo, S.-J.; Kim, S.-S.; Jeon, E.-J.; Lee, D.-S.; Kim, N.-Y. Antibacterial Activity, Antioxidant Capacity and Immune Gene Modulation of Six Medicinal Plants in CHSE-214 Cells. Fishes 2025, 10, 313. https://doi.org/10.3390/fishes10070313
Woo S-J, Kim S-S, Jeon E-J, Lee D-S, Kim N-Y. Antibacterial Activity, Antioxidant Capacity and Immune Gene Modulation of Six Medicinal Plants in CHSE-214 Cells. Fishes. 2025; 10(7):313. https://doi.org/10.3390/fishes10070313
Chicago/Turabian StyleWoo, Soo-Ji, So-Sun Kim, Eun-Ji Jeon, Dong-Sung Lee, and Na-Young Kim. 2025. "Antibacterial Activity, Antioxidant Capacity and Immune Gene Modulation of Six Medicinal Plants in CHSE-214 Cells" Fishes 10, no. 7: 313. https://doi.org/10.3390/fishes10070313
APA StyleWoo, S.-J., Kim, S.-S., Jeon, E.-J., Lee, D.-S., & Kim, N.-Y. (2025). Antibacterial Activity, Antioxidant Capacity and Immune Gene Modulation of Six Medicinal Plants in CHSE-214 Cells. Fishes, 10(7), 313. https://doi.org/10.3390/fishes10070313