
Biological Traits and Isoenzymatic Characterization of Triplophysa strauchii from the Turks River
 ,
,
Abstract

1. Introduction
2. Materials and Methods
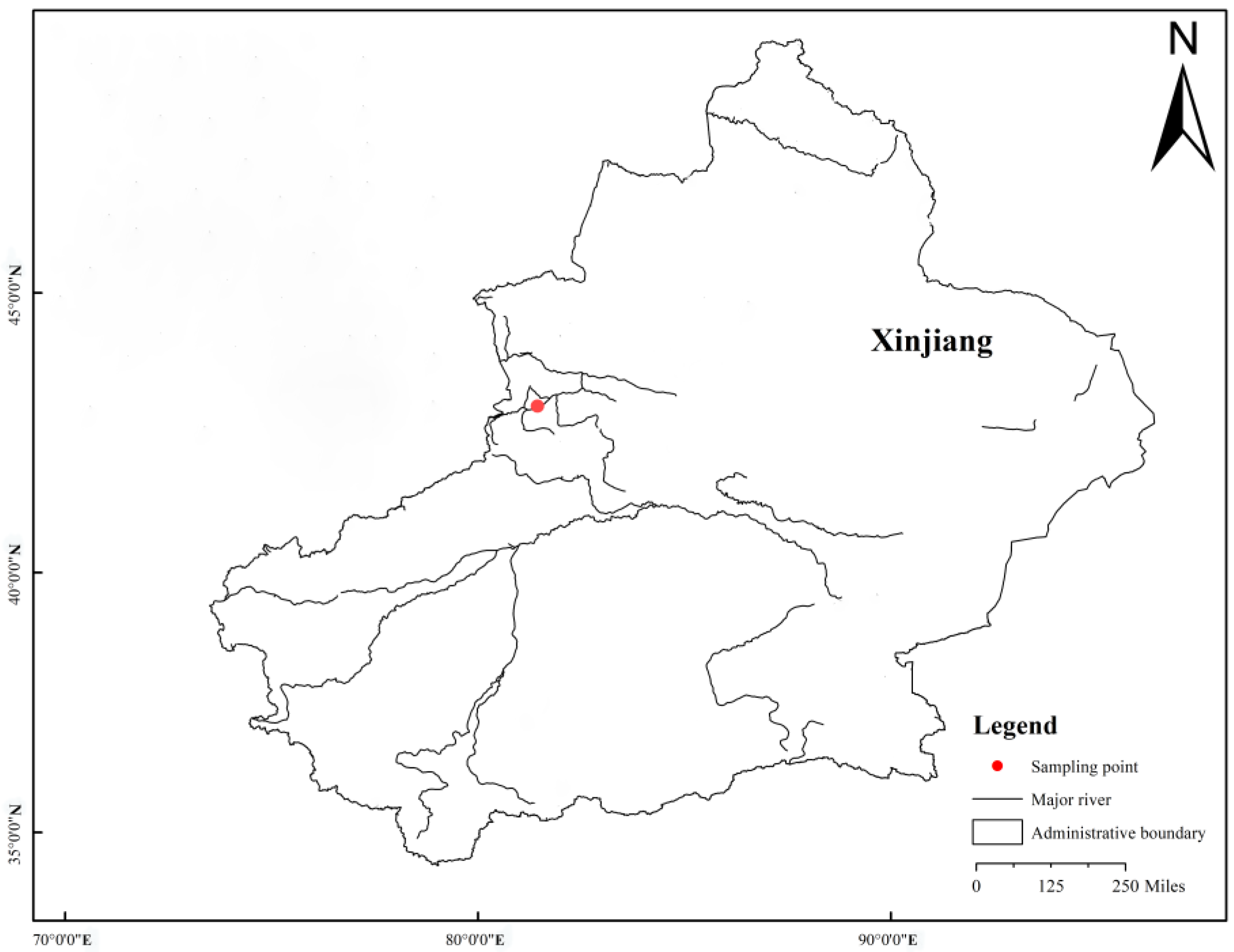
2.1. Sample Collection
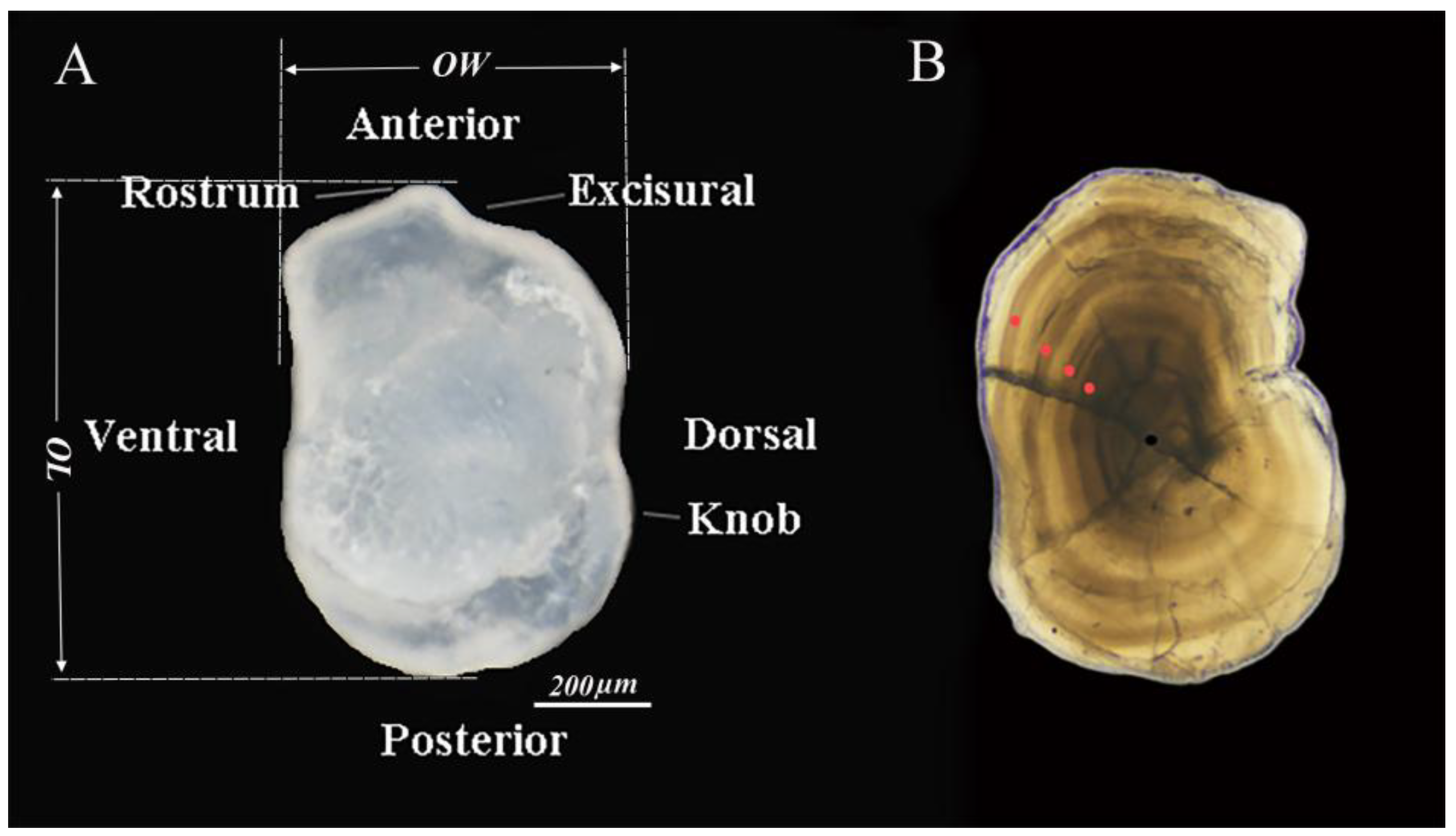
2.2. Otolith Morphological Characteristics
2.3. Age Determination Method
2.4. Growth Modeling
2.5. Gel Electrophoretic Analysis of Isoenzymes
2.6. Staining and Decolorization of Gels
2.7. Statistical Analysis
3. Results
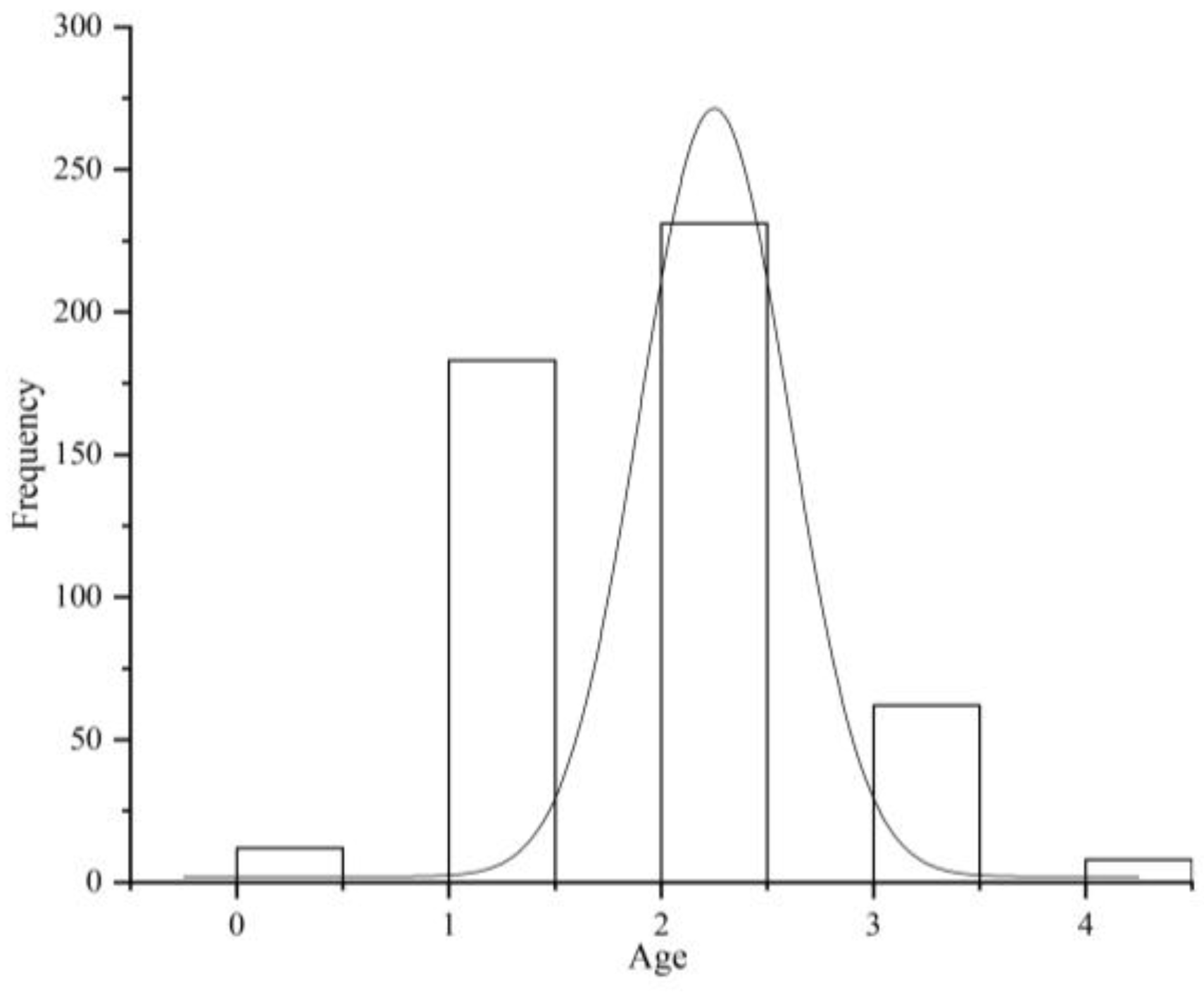
3.1. Age Composition of T. strauchii
3.1.1. Otolith Morphology
3.1.2. Age Determination
3.2. Growth Characteristics
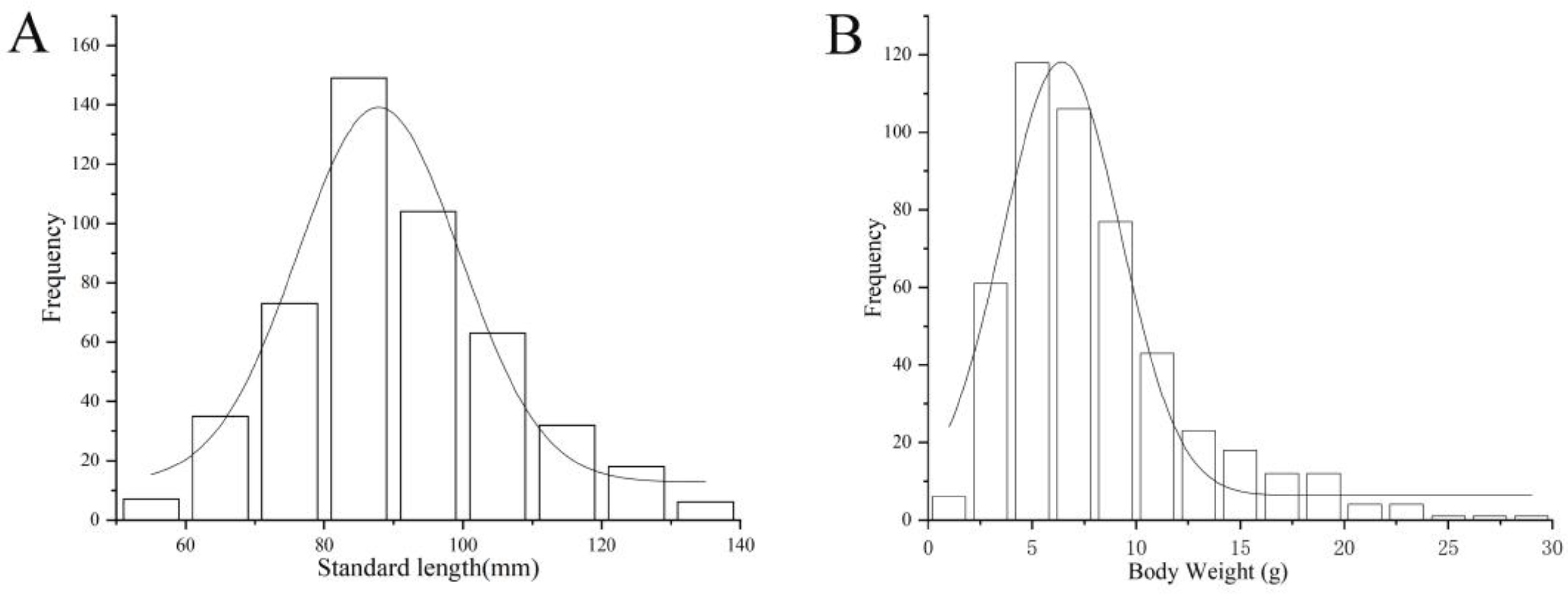
3.2.1. Distribution of SL (mm) and BW (g)
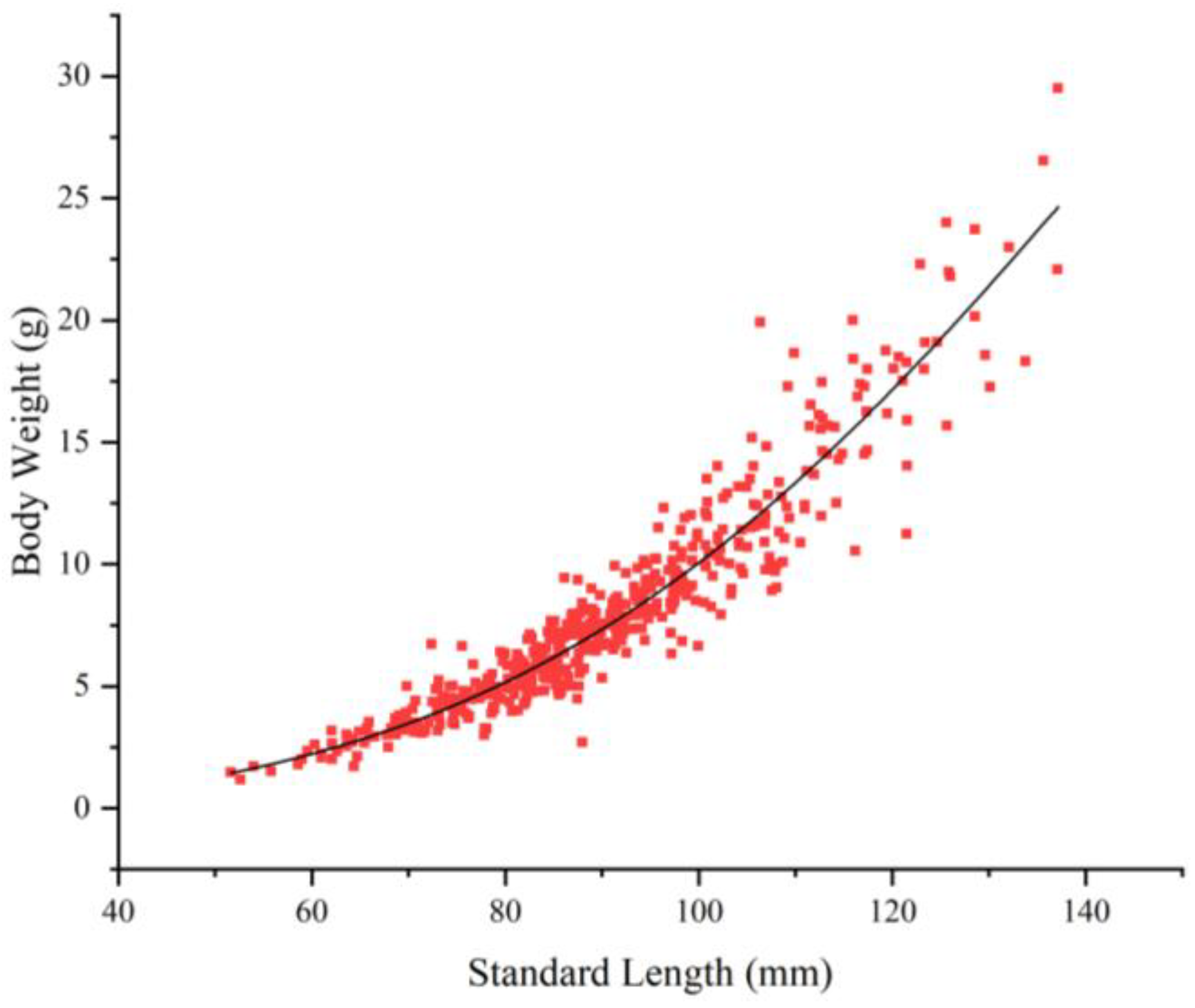
3.2.2. Relationship Between SL (mm) and BW (g)
3.2.3. Fulton’s Condition Index
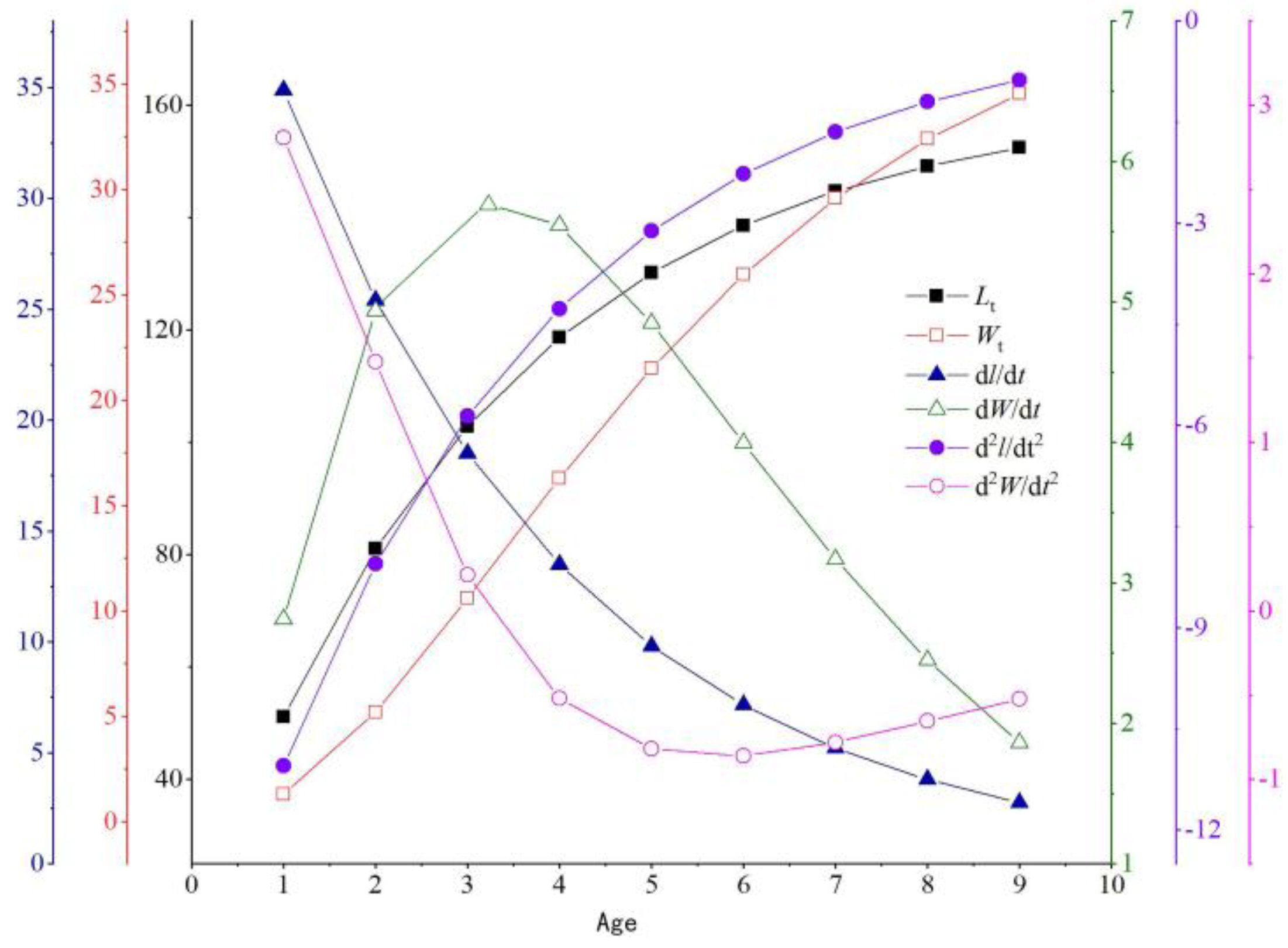
3.2.4. Growth Equation
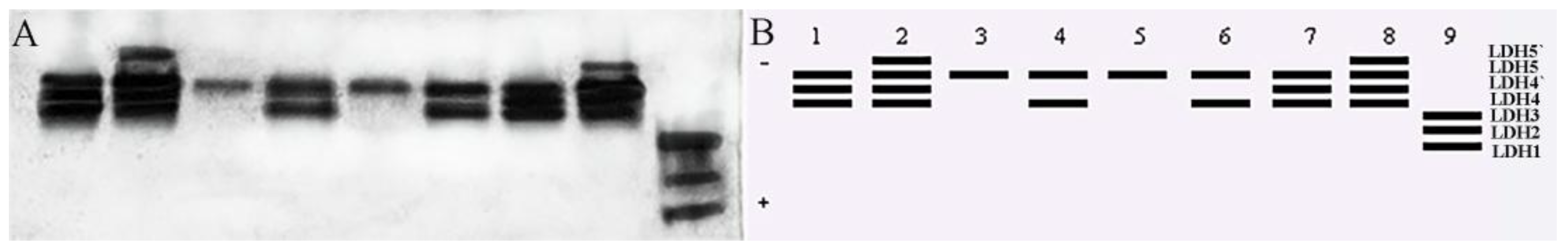
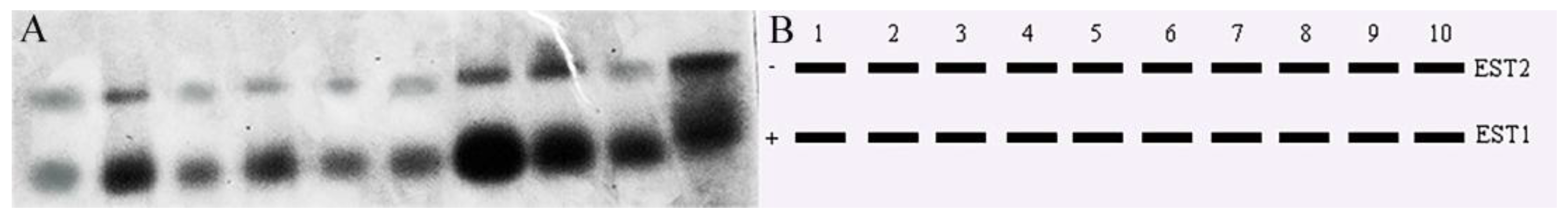
3.3. Tissue-Specific Expression of Isozymes in T. strauchii
4. Discussion
4.1. Biological Characteristics of T. strauchii in the Turks River
4.2. Isoenzymatic Characteristics of T. strauchii in the Turks River
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SL | Standard length |
| BW | Body weight |
| OA | Otolith area |
| Rmin | Minimum radius of otolith |
| Rmax | Maximum radius of otolith |
| OP | Otolith perimeter |
| OL | Otolith length |
| OW | Otolith width |
References
- Guo, Y.; Zhang, R.M.; Cai, L.G. Xinjiang of Fishery; Xinjiang Science and Technology Press: Urumqi, China, 2012. [Google Scholar]
- Cai, L.G.; Niu, J.G.; Liu, C.C.; Zou, M.; Xie, P.; Adakbek; Liu, J.; Li, H. Species Diversity and Dominant Fish Species in Different Reaches of the ILI River, XinJiang. Acta Hydrobiol. Sin. 2017, 41, 819–826. [Google Scholar]
- Guo, Y.; Cai, L.G.; Turxun. Study on the population structure of Triplophysa strauchii in Xinjiang in Sailimu Lake. Chin. J. Fish. 2002, 15, 1–5. [Google Scholar]
- Guo, Y.; Cai, L.; ADAKBEK. Biological characteristics of Triplophysa strauchii in Chawaopu Lake. Reserv. Fish. 2008, 3, 49–50+79. [Google Scholar]
- Kanu, U.C. The Complete mtDNA genome, Genetic diversity and population structure of Triplophysa strauchii (Cypriniformes: Cobitidae). Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2016. [Google Scholar]
- Shervette, V.R.; Rivera Hernández, J.M.; Nunoo, F.K.E. Age and growth of grey triggerfish Balistes capriscus from trans-Atlantic populations. J. Fish Biol. 2021, 98, 1120–1136. [Google Scholar] [CrossRef]
- Wang, X.Y.; Chen, S.A.; Wang, C.X.; Zi, F.Z.; Chang, D.S.; Xu, H.; Li, D.P. Otolith morphology and population discrimination of Triplophysa yarkandensis. Prog. Fish. Sci. 2023, 44, 201–211. [Google Scholar]
- Tian, N.; Yang, R.B.; Tan, B.Z.; Zeng, X.L.; He, L.Q.; Xu, Z.L.; Zhu, Z.; Liu, H.P.; Yang, X.F. Age, growth, and reproductive characteristics of Triplophysa stewarti in Lake Chugutso, Tibet. J. Fish. Sci. China 2022, 29, 1013–1021. [Google Scholar]
- Jin, S.S.; Wang, X.Y.; Lin, X.; Chen, S.A.; Liu, M.C.; Xie, C.X. Age and Growth of Triplophysa tenuis in Kaidu River, Xinjiang. Xinjiang Agric. Sci. 2020, 57, 181–189. [Google Scholar]
- Zhang, T.; Chen, P.; Sha, W.J.; Wang, R.X.; He, L.; Niu, J.G.; Gan, J.H. Electrophoretic Analysis on 3 Kinds of Isozymes in Gymnodiptychus dybowskii. Acta Agric. Boreali-Sin. 2023, 38 (Suppl. S1), 444–451. [Google Scholar]
- Silva, C.A.; Lima, R.C.; Teixeira, A.S. Isoenzyme electrophoretic patterns in discus fish (Symphysodon aequifasciatus Pellegrin, 1904 and Symphysodon discus Heckel, 1840) from the Central Amazon. Genet. Mol. Res. 2008, 7, 791–805. [Google Scholar] [CrossRef]
- Liu, J.Y. Study on Biology and Population Dynamics of Schizothorax macropogon in the Yarlung Tsangpo River. Master’s Thesis, Tarim University, Alar, China, 2016. [Google Scholar]
- Liu, Y.C.; Liu, S.Y.; Liu, H.P. Values of eight structures as age determination of Ptychobarbus dipogon, Tibet Autonomous Region. Acta Hydrobiol. Sin. 2019, 43, 579–588. [Google Scholar] [CrossRef]
- Yin, M.C. Fish Ecology; China Agriculture Press: Beijing, China, 1995; pp. 31–219, ISBN/7-109-03143-8. [Google Scholar]
- Ma, B.S. Study on the biology and population dynamics of Schizothorax oconnori. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2011. [Google Scholar] [CrossRef]
- Ren, B.; Ma, Y.W.; Tu, E.X.; Guo, Y.; Zhang, R.M.; Au, B.D.; Au, Z.Z.; Liu, Y. The study on the biology of Triplophysa (Hedinichthys) yarkandensis (Day) in Akesu River. Chin. J. Fish. 2004, 17, 46–52. [Google Scholar]
- Zang, L.; Gan, J.H.; He, L.; Zhang, T.; Zhou, J.G. Preliminary analysis on biochemical genetic characteristics and population genetic structure of Lota lota. Chin. Fish. Qual. Stand. 2022, 12, 45–50. [Google Scholar]
- Wei, Y.Z.; Yao, N.; Wang, S.; Xie, C.X.; Adak; Chen, S.A.; Zhou, F.R. Tissues specificity of three isozymes in Triplophysa yarkandensis. Heilongjiang Anim. Sci. Vet. Med. 2019, 1, 152–156. [Google Scholar]
- Chizinski, C.J.; Pope, K.L.; Wilde, G.R.; Strauss, R.E. Implications of stunting on morphology of freshwater fishes. J. Fish Biol. 2010, 76, 564–579. [Google Scholar] [CrossRef]
- Tokaç, A.; Herrmann, B.; Gökçe, G.; Krag, L.A.; Nezhad, D.S.; Lök, A.; Kaykaç, H.; Aydın, C.; Ulaş, A. Understanding the size selectivity of red mullet (Mullus barbatus) in Mediterranean trawl codends: A study based on fish morphology. Fish. Res. 2016, 174, 81–93. [Google Scholar] [CrossRef]
- Liu, L.; Zhou, Y.D.; Gao, T.X. A comparative study of different geographical groups of Sebastiscus marmoratus. J. Hydroecol. 2023, 44, 81–88. [Google Scholar]
- Campana, S.E.; Thorrold, S.R. Otoliths, increments, and elements: Keys to a comprehensive understanding of fish populations? Can. J. Fish. Aquat. Sci. 2001, 58, 30–38. [Google Scholar] [CrossRef]
- Cai, Z.L.; Duan, Y.J.; Xu, J.C.; He, Y.; Geng, J.Z.; Liu, X.; Han, M.X.; Liu, Y.H. Comparison of five structure materials in age determination of Amur ide Leuciscus waleckii in Dali Lake. J. Dalian Ocean. Univ. 2020, 35, 121–125. [Google Scholar]
- Chen, S.A. Study on Population Ecology of Triplophysa yarkandensis (Day) in Tarim river. Master’s Thesis, Huazhong Agricultural University, Hubei, China, 2012. [Google Scholar]
- Zeng, L.; Tang, W.H. Age, Body Growth and Reproductive Characteristics of Triplophysa yarkandensis. Chin. J. Zool. 2010, 45, 29–38. [Google Scholar]
- Munro, J.L.; Pauly, D. A simple method for comparing the growth of fishes and invertebrates. Fishbyte 2012, 1, 5–6. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretical Approach; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Kim, J.; Lee, S.; Nguyen, P.T.; Han, D.W.; Kim, I.C.; Kim, J.H. Length-weight relationships and condition factors of six notothenioid fish species occurring off King George Island and Northern Victoria Land (Antarctica). Polar Biol. 2023, 46, 1145–1150. [Google Scholar] [CrossRef]
- Bhattacharya, P.; Banik, S. Length-weight relationship and condition factor of the pabo catfish Ompok pabo (Hamilton, 1822) from Tripura, India. Indian J. Fish. 2012, 59, 141–146. [Google Scholar]
- Özcan, G.; Altun, A. Length-Weight Relationship and Condition Factor of Three Endemic and Threatened Freshwater Fishes from Orontes River. Pak. J. Zool. 2015, 47, 1637–1643. [Google Scholar]
- Ricker, W.E. Computation and interpretation of biological statistics of fish populations. Bull. Fish. Res. Board Can. 1975, 191, 382–385. [Google Scholar]
- Yang, L.; Lu, H.; Fu, M.; Bai, Y.P.; Su, S.Q.; Yao, W.Z. Age and growth characteristics of Coilia nasus in the Three Gorges Reservoir Region. Acta Hydrobiol. Sin. 2022, 46, 17–28. [Google Scholar]
- Russell, D.J.; Thomson, F.E.; Thuesen, P.A. Age and growth of two newly established invasive populations of Tilapia mariae in northern Australia. Fish Biol. 2013, 82, 1211–1225. [Google Scholar] [CrossRef]
- Chen, F.; Yuan, Y.J.; Wei, W.S.; Yu, S.L.; Fan, Z.A.; Zhang, R.B.; Zhang, T.W.; Shang, H.M. Variation and prediction trend of precipitation series for the Turks river basin during the last 236 years. J. Mt. Sci. 2010, 28, 545–551. [Google Scholar] [CrossRef]
- Niu, Y.J.; Ren, D.Q.; Chen, S.A.; Cai, L.G.; Niu, J.G.; Xie, C.X. Growth characteristics of Gymnodipty chusdybowskii Kessler in three tributaries of the Ili river in Xinjiang, China. J. Hydroecol. 2015, 36, 59–65. [Google Scholar] [CrossRef]
- Lester, N.P.; Shuter, B.J.; Abrams, P.A. Interpreting the von Bertalanffy model of somatic growth in fishes: The cost of reproduction. Proc. R. Soc. B-Biol. Sci. 2004, 271, 1625–1631. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Maravelias, C.D. Modelling fish growth: Multi-model inference as a better alternative to a priori using von Bertalanffy equation. Fish Fish. 2008, 9, 178–187. [Google Scholar] [CrossRef]
- Schofield, M.R.; Barker, R.J.; Taylor, P. Modeling Individual Specific Fish Length from Capture-Recapture Data using the von Bertalanffy Growth Curve. Biometrics 2013, 69, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y. Research on the age, growth, reproduction and population discrimination of Triplophysa yarkandensis. Master’s Thesis, Tarim University, Alar, China, 2023. [Google Scholar]
- Champasri, C.; Phetlum, S.; Pornchoo, C. Diverse activities and biochemical properties of amylase and proteases from six freshwater fish species. Sci Rep. 2021, 11, 5727. [Google Scholar] [CrossRef] [PubMed]
- Falcón, H.B.; Forrellat, B.A.; Carrillo, F.O.; Ubieta, H.K. Digestive enzymes of two freshwater fishes (Limia vittata and Gambusia punctata) with different dietary preferences at three developmental stages. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2011, 158, 136–141. [Google Scholar] [CrossRef]
- Ji, H.; Sun, H.T.; Xiong, D.M. Studies on activity, distribution, and zymogram of protease, α-amylase, and lipase in the paddlefish Polyodon spathula. Fish Physiol. Biochem. 2012, 38, 603–613. [Google Scholar] [CrossRef]
- Kolesnikova, E.E.; Soldatov, A.A.; Golovina, I.V.; Sysoeva, I.V.; Sysoev, A.A. Effect of acute hypoxia on the brain energy metabolism of the scorpionfish Scorpaena porcus Linnaeus, 1758: The pattern of oxidoreductase activity and adenylate system. Fish Physiol. Biochem. 2022, 48, 1105–1115. [Google Scholar] [CrossRef]
- He, G.; He, L.; Xu, Y.F.; Wang, X.G.; Li, X.L.; Zhang, Z. Isoenzyme analysis of Triplophysa xiangxiensis population. Chin. J. Ecol. 2009, 28, 2037–2041. [Google Scholar]
- Prakash, P.; Kumar, S.; Sardar, P.; Munilkumar, S.; Sahoo, S.; Satheesh, M.; Reena, H.; Mannur, V.; Patel, A. Optimization of weaning strategy in the climbing perch (Anabas testudineus, Bloch 1792) larvae on growth, survival, digestive, metabolic and stress responses. Fish Physiol. Biochem. 2023, 49, 1151–1169. [Google Scholar] [CrossRef]
- Vila, G.; Segura, J.A.; Ludemann, V.; Pose, G.N. Surface mycobiota of home-made dry cured sausages from the main producing regions of Argentina and morphological and biochemical characterization of Penicillium nalgiovense populations. Int. J. Food Microbiol. 2019, 309, 108312. [Google Scholar] [CrossRef]
- José, A.A.; Alvarez, M.C. Genetic identification of sparid species by isozyme markers: Application to interspecific hybrids. Aquaculture 1999, 173, 1–103. [Google Scholar]
- Xu, X.D.; Wang, H.H.; Li, Y.H.; Wu, B.; Ma, B.H.; Zeng, Q.X. Investigation on LDH and EST Isozymes in Different Tissues of Mastacembelus armatus. Hunan Arricultural Sci. 2020, 9, 76–78. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isoenzyme | Tris-HCl/mL | NAD/mg | NBT/mg | PMS/mg | Other Reagents |
|---|---|---|---|---|---|
| LDH | 1.5 mol/L Tris-HCl, pH 9.5, 15 mL | 30 | 30 | 10.5 | 1 mol/L sodium lactate, 15 mL. |
| MDH | 1.5 mol/L Tris-HCl, pH 9.5, 15 mL | 30 | 30 | 10.5 | 1 mol/L sodium malate, 15 mL; distilled water, 90 mL. |
| EST | 0.1 mol/L Tris-HCl, pH 8.0, 140 mL | - | - | - | α-Naphthyl acetate, 20 mg; β-naphthyl acetate, 20 mg; Solid Blue RR salt, 100 mg. |
| Parameter | Range | Mean ± S.D. |
|---|---|---|
| Otolith area, OA/mm2 | 0.32–1.13 | 0.66 + 0.12 |
| Minimum radius of otolith, Rmin/mm | 0.58–1.11 | 0.79 + 0.08 |
| Maximum radius of otolith, Rmax/mm | 0.72–1.46 | 1.07 + 0.11 |
| Otolith perimeter, OP/mm | 2.18–4.23 | 3.10 + 0.31 |
| Otolith length, OL/mm | 0.32–0.68 | 0.5 + 0.05 |
| Otolith width, OW/mm | 0.29–0.61 | 0.42 + 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serekbol, G.; Wang, C.; Yang, L.; Wang, H.; Liu, J.; Huo, Q.; Xu, H.; Song, Y.; Ren, D.; Chen, S. Biological Traits and Isoenzymatic Characterization of Triplophysa strauchii from the Turks River. Fishes 2025, 10, 239. https://doi.org/10.3390/fishes10050239
Serekbol G, Wang C, Yang L, Wang H, Liu J, Huo Q, Xu H, Song Y, Ren D, Chen S. Biological Traits and Isoenzymatic Characterization of Triplophysa strauchii from the Turks River. Fishes. 2025; 10(5):239. https://doi.org/10.3390/fishes10050239
Chicago/Turabian StyleSerekbol, Gulden, Chengxin Wang, Liting Yang, Huanhuan Wang, Jiaxuan Liu, Qiang Huo, Hao Xu, Yong Song, Daoquan Ren, and Shengao Chen. 2025. "Biological Traits and Isoenzymatic Characterization of Triplophysa strauchii from the Turks River" Fishes 10, no. 5: 239. https://doi.org/10.3390/fishes10050239
APA StyleSerekbol, G., Wang, C., Yang, L., Wang, H., Liu, J., Huo, Q., Xu, H., Song, Y., Ren, D., & Chen, S. (2025). Biological Traits and Isoenzymatic Characterization of Triplophysa strauchii from the Turks River. Fishes, 10(5), 239. https://doi.org/10.3390/fishes10050239