Abstract
The onion (Allium cepa L.) belongs to the family Alliaceae and has paramount importance among vegetable crops around the globe. Salinity stress and the soil-borne fungus Fusarium oxysporum f. sp. Cepa (FOC), which is the cause of basal rot, are potentially hazardous factors related to the yield of onion crops. The present study was conducted in CRD using three replicates in in vitro conditions and in a greenhouse to investigate the impact of stress factors (salinity levels and FOC) (T1 = Negative control; T2 = Positive control (FOC); T3 = EC 2.5 dS m−1; T4 = EC 3.5 dS m−1; T5 = EC 4.5 dS m−1; T6 = EC 2.5 dS m−1 + FOC; T7 = EC 3.5 dS m−1 + FOC; and T8 = EC 4.5 dS m−1 + FOC) on the morphological, physiological, and yield parameters of onions. Fungal biomass, in the in vitro condition, increased at a salinity level of 2–8 dS m−1. In the greenhouse, the application of salinity levels, viz. 2.5, 3.5, and 4.5 dS m−1, and fungal pathogens exhibited a more severe impact on the severity of the disease, onion growth, and bulb compared to sole stress. Overall, the decreased levels of osmotic potential, total chlorophyll content, membrane stability index, and total protein were assessed in onion leaves, whereas total phenolics were enhanced in the mentioned stress conditions. Sugar contents were reduced due to the sole application of pathogens, whereas they were enhanced in the sole application of salinity stress and increased due to a combination of FOC and salinity level applications. Polyphenol oxidase (PPO) activity was not altered as a result of pathogen infection, while peroxidase (PO) and phenylalanine ammonia-lyase (PAL) contents were enhanced, and the activity of catalase (CAT) was reduced. Sole stress along with the combined application of salinity levels with fungal pathogens exhibited enhanced PO and PPO activity; in contrast, CAT and PAL activity decreased. In summary, onion crop cultivation in saline soil conditions increases the disease’s severity, which is caused by FOC. Furthermore, it is crucial to carry out field trials while considering these outcomes in order to support a strong strategy for reducing the tested stresses within a wider range of environmental conditions.
1. Introduction
Soil salinization poses a key abiotic threat to agriculture by minimizing the growth of plants, and it exerts a negative impact on productivity around the globe [1]. Salinity adversely affected a large area (around 10 million ha) of Pakistan, which comprises 12.9% of the country’s mainland, the main cause of which is insufficient drainage conditions [2,3]. The nutrition and yield of various crops, viz. vegetables, are severely affected due to soil salinity as vegetables are more sensitivity toward salt stress [4]. Various morphological, physiological, biochemical, growth, and yield attributes of plants were impacted negatively in salt-stress conditions [1]. Moreover, in vegetables, a reduction in biomass has also been documented in salt-stress environments [5]. Along with its impact on plants, salinity stress also increases the susceptibility towards a variety of pathogens, which aggrevates plant diseases due to various soil-borne fungi [6]. Thus, salinity stress could be a vital aspect with respect to the increase in disease incidence in various crops. The onion (Allium cepa L.) has paramount importance among vegetable crops and occupies approximately 5.2 million hectares worldwide, contributing 100 million tons in terms of production [7]. In Pakistan, onions are cultivated within an area measuring 141 thousand hectares, contributing 2108.8 thousand tons in terms of production [8]. Onion is widely affected by the soil-borne fungus Fusarium oxysporum f. sp. Cepa (fusarium basal rot) [9] because it can remain viable in the soil for a long period of time in the absence of a host [10] and can cause severe losses to onion crops [11]. Onion plants infected with FBR exhibit disease symptoms at all growth stages, viz. seedling damping-off, leaf chlorosis, and necrosis with discoloration of basal plate tissues [12].
The plant exhibits various peculiar behaviors towards different biotic and abiotic stress inputs, alone and in combination, and other general reactions [13]. The induction of reactive oxygen species (ROS) plays a vital role against various abiotic and biotic factors that trigger various responses in plants, viz., PR proteins, hypersensitive response (HR), secondary metabolite, and enzyme production as well as programmed cell death (PCD) [14]. An increase in enzyme activities, viz. peroxidase (PO), superoxide dismutase (SOD), and catalase (CAT), is correlated with the endurance of salt in a variety of plants [15]. Furthermore, these enzymatic responses differ in terms of the intensity, time period, and collective impact of abiotic and biotic factors [15]. Considering the response of plants towards concurrent stresses is of paramount importance, it could be helpful for researchers in evaluating sole stress and in combining the effect of plant pathogens and salinity with respect to the performance of onions. Consequently, the primary objective of the proposed investigation was to evaluate the exclusive and combined effects of FOC and salinity on the morphology, growth, biochemistry, and yield of the onion.
2. Materials and Methods
2.1. Impact of Salinity on FOC Performance in Laboratory
In vitro experiments were conducted to evaluate the impact of different saline solutions on F. oxysporum growth. Calcium chloride, sodium chloride, magnesium sulphate, and sodium sulphate were dissolved in double-distilled water (100 mL) to prepare saline solutions with altered range of EC (2–8 dS m−1) using Digital LCD EDS EC meter temperature tester (Table 1).

Table 1.
Various salt concentrations (g 100 mL−1) used to prepare saline solutions.
Preparations were conducted in sterilized 250 mL flask having mixture of saline solutions of different EC 2 to 8 dS m−1 and 2 g ME (malt extract) 100 mL−1. Inoculation of flasks was achieved with FOC (2 nm disc) and kept in an incubator at 28 ± 2 °C for 7 days. Dried fungal biomass was obtained using oven at 60 °C for 24 h.
2.2. Analysis of Physio-Chemical Behavior of Soil
Loamy soil (42.5%:32.3%:25.2%) was used at 7.6 pH, 36% saturation level, 0.36% organic matter, and 1.5 dS m−1 EC [16]; 3 sodium absorption ratio (SAR); 8 meq L−1 calcium and magnesium (Ca++, Mg++); 6 meq L−1 sodium (Na++); 5 meq L−1 carbonate (CO3−); 5.5 meq L−1 sulphate (SO4−2); 4.5 meq L−1 bicarbonate (HCO3−); and 3 meq L−1 chloride (Cl−) [17]. Magnesium + Calcium was estimated by EDTA titration, Sodium in the soil extract was measured using flame photometer. Sulphate was measured using the barium sulfate precipitation method. Carbonate and bicarbonate were determined by titrating soil extract against 0.01 N H2SO4.
2.3. Preparation of Saline Soil
Earthen pots (30 cm length and 22 cm diameter) were used for experimentation. Known concentrations of salts were added to change soil salinity. Quadratic formula and SAR were applied to determine the quantity of salt required to amend the soil [18]. A total of 12 L of double distilled water with known concentrations of various salts were supplemented in 50 kg of soil for making soil with EC 2.5 dS m−1, viz. sodium chloride (1.37 g), calcium chloride (2.19 g), sodium sulphate (3.41 g), and magnesium sulphate (1.36 g); 3.5 dS m−1, viz. sodium chloride (1.32 g), calcium chloride (6.84 g), sodium sulphate (5.94 g), magnesium sulphate (3.36 g); and 4.5 dS m−1, viz. sodium chloride (6.31 g), calcium chloride (10.92 g), sodium sulphate (9.11 g), and magnesium sulphate (5.82 g). Successive irrigations with distilled water were performed for a period of four months at regular intervals (20–25 days) to obtain salt equilibrium in soil.
To evaluate the synergistic effect of pathogen (FOC) and salinity, soil with all three salinity levels were inoculated with fungal suspension (40 mL). Onion seedlings (VARIO-2), 40 days old, were transferred into pots. Eight treatments, viz. T1 = Negative control, T2 = Positive control (FOC), T3 = EC 2.5 dS m−1, T4 = EC 3.5 dS m−1, T5 = EC 4.5 dS m−1, T6 = EC 2.5 dS m−1 + FOC, T7 = EC 3.5 dS m−1 + FOC, and T8 = EC 4.5 dS m−1 + FOC were used. The experiments were terminated after 50 days. Three replications were conducted. Treated pots were positioned in a controlled environment (greenhouse with RH 65% and temperature at 27 °C ± 3). Disease symptoms of fusarium basal rot (FBR) were evaluated using a 1:5 disease rating scale [19].
Impact of used treatments, after 35 days of inoculation, were analyzed by considering the growth, morphology, physio-chemical properties, and yield attributes.
2.4. Physiological Assays
The osmotic potential of the cell sap of onion leaves was calculated using a vapor pressure osmometer (Wescor model VAPRO 5520) according to the method described by Capell and Dorffling [20]. Carotenoids and total chlorophyll content were quantified as illustrated by Lichtenthaler and Wellburn [21]. Protein content in the samples were assessed according to Lowry et al. [22]. Total sugar content was assessed according to Nelson [23] to find out the absorbance of the sample at 620 nm. The membrane stability index (MSI) was evaluated using the method of Saneoka et al. [24].
2.5. Antioxidant Enzymes Assays
A homogenized mixture of onion leaves (0.5 g) and 10 mL of Na2SO4 buffer solution (pH 6.5) was centrifuged (10,000 rpm) for a period of 10 min. Peroxidase (PO) activity was measured following the protocol of Urbanek et al. [25]. Polyphenol oxidase (PPO) was measured according to the method of Gauillard et al. [26]. Phenylalanine ammonia-lyase (PAL) activity was estimated using the technique given by Nagarathna et al. [27]. The assessment of phenolics was conducted according to Malik and Singh [28]. The conversion of L-phenylalanine to transcinnamic acid was measured at 290 nm [29]. Catalase (CAT) activity was assessed using the method described by Maehly and Chance [30].
2.6. Growth Assays
Plant growth attributes (length and weight) and bulb growth parameters (weight and diameter) were detected after 60 days of inoculation. The dry weight of the root and shoot were recorded by drying in an oven at 80 °C for 24 h.
2.7. Statistical Analysis
Collected data were statistically analyzed using Statistix 8.01 software by applying Fisher’s analysis of variance. Statistical significant differences are indicated in descending order (a, b, c, etc.) at p ≤ 0.05 (Tukey’s HSD test).
3. Results
3.1. Impact of Saline Solution on Fungal (FOC) Growth in Controlled Conditions
The growth of fungus was significantly increased (p ≤ 0.05) with increasing EC levels compared to the control. Fungal dry biomass showed the maximum value (289.43 mg) at 4 ds m−1 EC in comparison to the control group (160.73 mg). The increase in fungal dry biomass was 45–80% in the case of 2–4 ds m−1 EC, whereas 32–37% dry biomass of fungus was observed at EC levels between 5 and 8 ds m−1 (Figure 1).
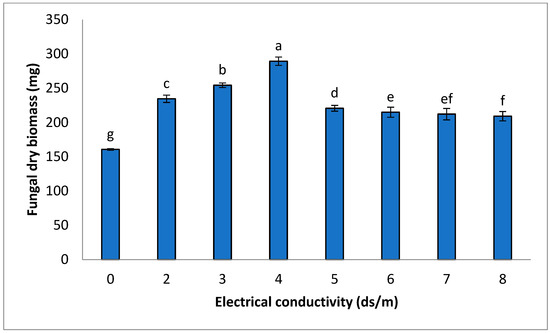
Figure 1.
Impact of various EC levels on FOC biomass, cultured on malt extract medium (MEA), after 7 days of incubation. Statistical significant differences (p ≤ 0.05) are shown with different letters in descending order (a, b, c, etc.).
3.2. Impact of FOC on Growth Attributes of Onions under Stressed Condition
Sole application of FOC (T2) exhibited 55% disease severity over T1 and placed in the fourth rank according to the disease rating scale. Moreover, the infected bulbs showed disease symptoms, viz. uneven shape, softness, and basal plate discoloration. The combination of salinity levels with FOC (T6–T8) showed higher disease severity (76–100%), ranked fifth according to the disease rating scale, and disintegrated the bulbs to a greater extent than sole application of salinity or FOC (Figure 2).
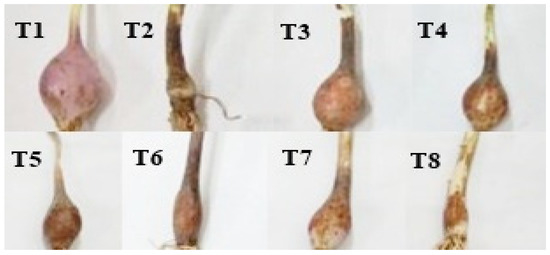
Figure 2.
Impact of various salinity levels and FOC on onion bulb.
Various level of salinity, viz. 2.5, 3.5, and 4.5 ds m−1 (EC), alone and in combination with FOC, significantly (p ≤ 0.05) impacted the bulb diameter, bulb weight, shoot length, shoot dry weight, root length, and root dry weight (Figure 3). In control treatments, maximum shoot length (cm) was observed in T1 (23.56) (+ve) compared to T2 (11.63) (-ve). Regarding salinity levels alone, a decreasing trend, viz. T3 (19.16) > T4 (14.2) > T5 (11.16), was observed. A combination of salinity and FOC showed that shoot length was decreased in T6 followed by T7 and T8 (Figure 3). Shoot dry weight (g) was significantly (p ≤ 0.05) higher in T1 (0.66) compared to to T2 (0.22). Shoot dry weight in T3–T5 salinity levels (2.5–4.5 ds m−1) was reduced between 0.44 and 0.23 g. Regarding the combination of salinity levels with FOC viz. T6–T8 shoot dry weight was decreased from 0.42 to 0.14 g (Figure 3). Root length (cm) was significantly increased (p ≤ 0.05) in the negative control (4.53) compared to the positive control (2.23). Sole application of salinity levels (T3–T5) decreased the root length between 3.43 and 2.24 cm, whereas T6–T8 exhibited a greater decreased in root length of between 2.33 and 1.5 cm (Figure 3). The negative control (T1) exhibited maximum (0.07) root dry weight (g) compared to the positive control (0.02) and other used treatments (T3–T8). Sole application of various salinity levels (T3–T5) resulted in root dry weight ranging between 0.05 and 0.03 g, while T6–T8 exhibited a root dry weight between 0.04 and 0.02 g (Figure 3). Bulb diameter (cm) of the onions was significantly (p ≤ 0.05) higher (3.80) in T1 compared to T2 (3.33). Salinity levels (T3–T5) resulted in a bulb diameter of the onions between 3.43 and 3.10 cm, while combined application of salinity levels with FOC (T6–T8) resulted in a bulb diameter ranging between 1.9 and 2.6 cm (Figure 4). The negative control (T1) exhibited significantly (p ≤ 0.05) higher (6.4) bulb weight (g) of the onions compared to the positive control (T2) (2.23). Sole application of salinity levels (T3–T5) resulted in bulb weight between 2.23 and 1.70 g, while T6–T8 exhibited bulb weight of 1.80–1.56 g (Figure 4).
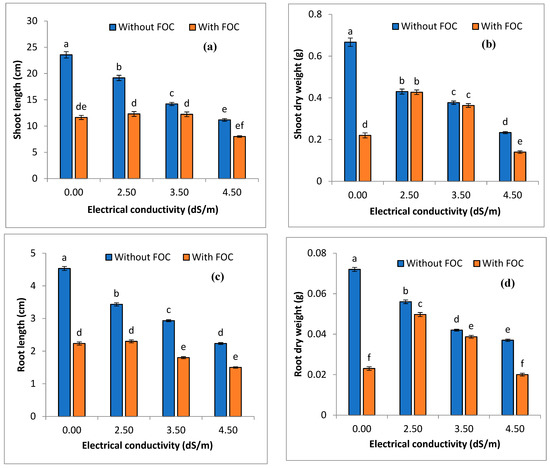
Figure 3.
Impact of various salinity levels and FOC on shoot length (a), shoot dry weight (b), root length (c), and root dry weight (d) of onions on the 50th day of application. Statistical significant differences (p ≤ 0.05) are shown with different letters in descending order (a, b, c, etc.).
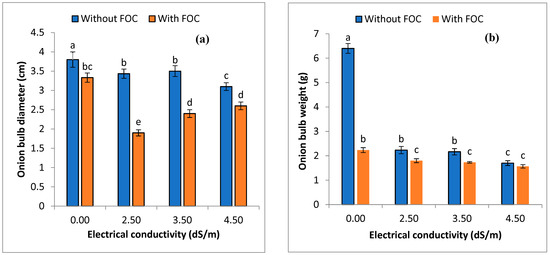
Figure 4.
Impact of various salinity levels and FOC on bulb diameter (a) and bulb weight (b) of onions on the 50th day of application. Statistical significant differences (p ≤ 0.05) are shown with different letters in descending order (a, b, c, etc.).
3.3. Impact of Stress Conditions on Physiological Attributes of Onions
All salinity levels (T3–T5), including the positive control stress, and the combination with FOC (T6–T8) significantly (p ≤ 0.05) impacted all physiological attributes. Sole application of salinity levels (T3–T5) negatively impacted the physiological parameters as osmotic potential (0.93–0.53 MPa), total chlorophyll content (0.91–0.80 mg/g), membrane stability index (58.33–38.00%) and total protein content (1.37–0.61 mg/g), whereas, total sugar content as well as total phenolics increased to 1.47–2.47 mg/g and 6.50–5.39 mg/g, respectively. The combination of salinity levels with FOC (T6–T8) impacted the physiological aspects, such as osmotic potential (1.08–0.74 MPa), total chlorophyll content (0.77–0.74 mg/g), membrane stability index (55.00–47.33%), and total protein content (1.62–0.67 mg/g), whereas total sugar content (5.04 mg/g) as well as total phenolics (6.63 mg/g) levels were enhanced. This increase became less pronounced with increasing soil salinity (2.5–4.5 dS m−1). Total chlorophyll content, membrane stability index, osmotic potential, and total protein content were higher in the negative control (T1) than in the positive control (T2) (Table 2).
Table 2.
Impact of various salinity levels and FOC on the physiological attributes of onions on 35th day of inoculation.
3.4. Impact of Stress Conditions on Biochemical Attributes of Onions
Enzymatic activity (units/min/mg of protein) (PPO) was insignificantly impacted, while PO and PAL significantly increased (p ≤ 0.05). In contrast, catalase (CAT) activity was reduced with respect to T1. Sole application of salinity levels (T3–T5) impacted enzymatic activity, such as PO (790–2020), PPO (8.26–4.33), PAL (7.56–4.80) and catalase (1507–541.30), whereas the combined application of salinity levels with FOC (T6–T8) influenced the enzymatic activity, such as PO (999–2177.3), PPO (5.80–4.20), PAL (6.56–2.83), and catalase (1360–502.70) (Figure 5).
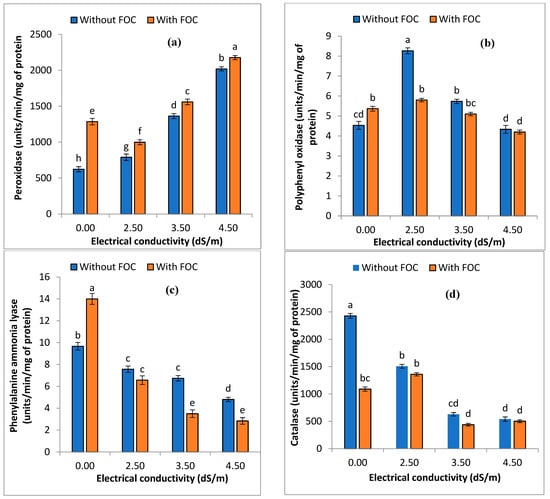
Figure 5.
Impact of various salinity levels and FOC on peroxidase (a), polyphenyl oxidase (b), phenylalanine ammonia lyase (c), and catalase (d) of onions on the 35th day of application. Statistical significant differences (p ≤ 0.05) are shown with different letters in descending order (a, b, c, etc.).
3.5. Correlation and Principal Component Analysis
In order to establish the credibility of the observations in onion plants, an examination was conducted on growth, yield, and the physio-chemical parameters to ascertain negative and positive correlations (Table 3). All attributes, viz. PO, TPh, TSC, and OP, are negatively correlated with BD, BW, CAT and MSI, whereas PPO, RL, RW, SL, SW, TChl, TPrC were negatively correlated with OP. PO was negatively correlated with RL, RW, SL, SW, TChl, and TPrC. On the other hand, RL and RW were positively correlated with RW, SL, SW, TChl, and TPrC. Onions cultivated in various stress levels (T1–T8) exhibited connection between the growth, yield, and various physio-chemical attributes using a PCA (principal component analysis) plot (Figure 6). The primary components, viz. Dim1 and Dim2, made the highest percentage and account for 77.2% of total database. Dim1 comprises 60.7%, whereas Dim2 comprises 16.5%, of the database.
Table 3.
Correlation matrix of growth, yield and physio-chemical attributes of onions in response to different stress levels of salinity and FOC.
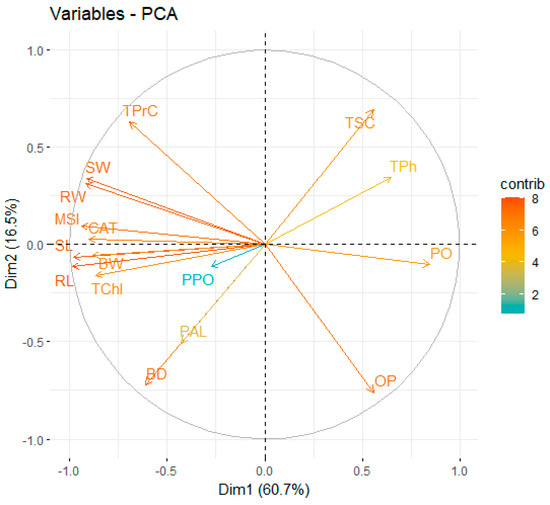
Figure 6.
Principal component analysis plot representing correlations among variables for various stress levels in onions. BD = Bulb diameter, BW = Bulb weight, CAT = Catalase, MSI = Membrane Stability Index, OP = Osmotic potential, PAL = Phenylalanine ammonia lyase, PO = Peroxidase, PPO = Polyphenyl oxidase, RL = Root length, RW = Root dry weight, SL = Shoot length, SW = Shoot dry weight, TChl = Total chlorophyll content, TPh = Total phenolics, TPrC = Total protein content, TSC = Total sugar content.
4. Discussion
The present findings showed 45–80% enhancement in FOC biomass at 2–4 dS m−1 and 32–37% enhancement at 5–8 dS m−1 compared to the control. Boumaaza et al. [31] indicated similar results in a way, as various levels of salinity (NaCl) enhanced the production of conidia in Fusarium oxysporum and exhibited a slight impact, at higher concentrations, on the germination of conidia. Tawfik et al. [32] revealed that sodium salts at higher concentrations inhibit the growth of Fusarium semitectum. Boumaaza et al. [33] documented that NaCl had a stimulatory impact at 150 ppm on fungal mycelial growth, conidial production, and germination in various fungi, viz. Botrytis cinerea and Verticillium dahlia. Microorganisms cope with higher salt stress by adopting osmotic and ionic potential mechanisms [34]. Microbes acclimatize to salinity stress by accruing various inorganic and organic osmolytes [35] which involve higher energy levels [36]; and ultimately causing a reduction in activity and growth. Hence, a boost in biomass of FOC at 2–4 dS m−1 EC levels might exhibit selective accumulation of solute indispensable for metabolism by counteracting osmotic pressure enhancement, whereasan increase in EC level (5–8 dS m−1) caused the enhancement of the solute level with a negative increase in osmotic potential, resulting in the leakage of cells’ water, which can affect the microorganisms by plasmolysis [32].
In the current study, the infection caused by the tested pathogen solely or in combination with various salinity levels exhibited a negative impact on growth parameters. Additionally, there was an obvious increase in fusarium basal rot severity and disease symptoms viz. uneven shape, softness, and basal plate discoloration [9]. These results were in harmony with Shoaib et al. [15], who observed similar disease symptoms on onions. Plants infected with FOC showed stunted growth, succulent leaves, rotted bulbs, and discoloration of the base plate [37]. Salinity stress negatively impacted onions due to restriction in water absorption, intake of minerals, malfunction of the membrane, and metabolic disturbance [38].
Physiological attributes measured in the current study exhibited a declining trend for both the sole and combined treatments. This might be due to the mycotoxin (fusaric acid), which lowers the stem hydraulic conductance and leaf water potential, which cause membrane injury and the leakage of water [15]. Wang et al. [39] found that photosynthesis reduction closes the stomata, which causes a metabolic disturbance while reducing the accumulation of sugar content. Osmotic pressure decreased with increasing the salinity level [40]. Hence, the photosynthetic reduction and the changing of metabolic processes may occur due to the accumulation of sugar that could be ascribed to salinity [15].
Regarding the biochemicals, phenolic activity was increased, but not at all levels, to resist the invading fungi. Furthermore, The activities of PO and PAL were increased. On the other hand, CAT activity was decreased, whereas no significant enhancement was observed in case of PPO. The decrease in CAT activity could be attributed to the high proteolysis in response to stress [41]. Total phenolics were higher in the case of salinity stress (T3) over other salinity levels. Salt stress increased the PO and PPO activity, whereas the activity of CAT and PAL decreased with the increasing salinity levels [42]. A combination of FOC and salinity levels exhibited the general behavior of increase/decrease regarding physio-chemical attributes, which were in line with the sole application of salinity or FOC. The oxidative burst of cellular components could be accredited to the increase in production of PPO and PO, whereas it negatively impacted the that of PAL and CAT [15]. Mansha et al. [43] also documented a similar increase/decrease in biochemical attributes in onions under stress conditions. The results of current study revealed that the enzymatic activation under stress conditions is relatively inconstant. Additionally, peroxidase could be a part of the paramount importance in the induction of antioxidants in onions.
5. Conclusions
In this study, a combination of salinity levels and a fungal pathogen exhibited severe damage on the growth and production of onions when compared to sole stress, while crop damage (severity) is correlated with salinity levels. Under salt stress, the physiology of onion plants is disturbed, and as a result, the plant becomes weak and more prone to virulent pathogens. Furthermore, field trials should be conducted as a continuity of this work to maintain the efficient strategy in the mitigation of stresses under more diverse environmental conditions.
Author Contributions
Conceptualization, M.Z.M., H.M.A., K.I., C.M.S.H. and A.S.; formal analysis, R.I. and Q.u.Z.; investigation, S.M.A.-Q.; methodology, N.A.A.-H., M.F.M.I. and W.A.O.; resources, M.Z.M., software, H.M.A.; supervision, K.I.; validation, A.S. and S.M.A.-Q.; visualization, N.A.A.-H.; writing—original draft, W.A.O.; writing—review and editing, M.F.M.I. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Deanship of Scientific Research, Taif University.
Data Availability Statement
All concerned data are available in the form of tables and figures.
Acknowledgments
The researchers would like to acknowledge the Deanship of Scientific Research, Taif University, for funding this work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kibria, M.G.; Hoque, M.A. A Review on Plant Responses to Soil Salinity and Amelioration Strategies. Open J. Soil Sci. 2019, 9, 219–231. [Google Scholar] [CrossRef]
- Syed, A.; Sarwar, G.; Shah, S.H.; Muhammad, S. Soil Salinity Research in 21st Century in Pakistan: Its Impact on Availability of Plant Nutrients, Growth and Yield of Crops. Commun. Soil Sci. Plant Anal. 2021, 52, 183–200. [Google Scholar] [CrossRef]
- Mahboob, W.; Khan, A.; Shirazi, M.U. Induction of Salt Tolerance in Wheat (Triticum aestivum L.) Seedlings through Exogenous Application of Proline. Pak. J. Bot. 2016, 48, 861–867. [Google Scholar]
- Machado, R.M.A.; Serralheiro, R.P. Soil Salinity: Effect on Vegetable Crop Growth. Management Practices to Prevent and Mitigate Soil Salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- Chourasia, K.N.; More, S.J.; Kumar, A.; Kumar, D.; Singh, B.; Bhardwaj, V.; Kumar, A.; Das, S.K.; Singh, R.K.; Zinta, G.; et al. Salinity responses and tolerance mechanisms in underground vegetable crops: An integrative review. Planta 2021, 255, 68. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D.; Davranov, K.; Wirth, S.; Hashem, A.; Abd_Allah, E.F. Impact of soil salinity on the plant-growth–promoting and biological control abilities of root associated bacteria. Saudi J. Biol. Sci. 2017, 24, 1601–1608. [Google Scholar] [CrossRef]
- FAO. Crop Water Information; Food and Agriculture Organization of the United Nations. 2021. Available online: https://www.fao.org/land-water/databases-and-software/crop-information/en/ (accessed on 7 November 2021).
- Economic Survey of Pakistan (ESP), Government of Pakistan, Ministry of Food, Agriculture and Livestock; Federal Bureau of Statistics: Islamabad, Pakistan, 2021.
- Le, D.; Audenaert, K.; Haesaert, G. Fusarium basal rot: Profile of an increasingly important disease in Allium spp. Trop. Plant Pathol. 2021, 46, 241–253. [Google Scholar] [CrossRef]
- Afzal, M.M.; Sahi, S.T.; Habib, A.; Ashraf, W.; Zeshan, M.A.; Raheel, M.; Shakeel, Q. Management of Fusarium Basal Rot of Onion caused by Fusarium oxysporum f.sp. cepae through Desert Plants Extracts. Sarhad J. Agric. 2021, 37, 1377–1383. [Google Scholar] [CrossRef]
- Shamyuktha, J.; Sheela, J.; Rajinimala, N.; Jeberlinprabina, B.; Ravindran, C. Survey on Onion Basal Rot Disease Incidence and Evaluation of Aggregatum Onion (Allium cepa L. Var. Aggregatum Don.) Genotypes Against Fusarium oxysporum f. sp. Cepae. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 529–536. [Google Scholar] [CrossRef]
- Cramer, C.S.; Mandal, S.; Sharma, S.; Nourbakhsh, S.S.; Goldman, I.; Guzman, I. Recent Advances in Onion Genetic Improvement. Agronomy 2021, 11, 482. [Google Scholar] [CrossRef]
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; Senthil-Kumar, M. Impact of Combined Abiotic and Biotic Stresses on Plant Growth and Avenues for Crop Improvement by Exploiting Physio-morphological Traits. Front. Plant Sci. 2017, 8, 537. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Cheng, Z.; Ahmad, H.; Hayat, S. Reactive oxygen species (ROS) as defenses against a broad range of plant fungal infections and case study on ROS employed by crops against Verticillium dahliae wilts. J. Plant Interact. 2018, 13, 353–363. [Google Scholar] [CrossRef]
- Shoaib, A.; Meraj, S.; Nafisa; Khan, K.A.; Javaid, M.A. Influence of salinity and Fusarium oxysporum as the stress factors on morpho-physiological and yield attributes in onion. Physiol. Mol. Biol. Plants 2018, 24, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- McLean, E.O. Soil pH and Lime Requirement. In Methods of Soil Analysis, Part 2. Chemical and Microbiological Properties; Page, A.L., Ed.; American Society of Agronomy, Soil Science Society of America J.: Madison, AL, USA, 1982; pp. 199–223. [Google Scholar]
- Richards, L.A. Diagnosis and Improvement of Saline and Alkali Soils; Agric. Handbook No 60; US Government Printing Office: Washington, DC, USA, 1954; Volume 78, p. 154.
- Pang, H.-C.; Li, Y.-Y.; Yang, J.-S.; Liang, Y.-S. Effect of brackish water irrigation and straw mulching on soil salinity and crop yields under monsoonal climatic conditions. Agric. Water Manag. 2010, 97, 1971–1977. [Google Scholar] [CrossRef]
- Coşkuntuna, A.; Özer, N. Biological control of onion basal rot disease using Trichoderma harzianum and induction of antifungal compounds in onion set following seed treatment. Crop Prot. 2008, 27, 330–336. [Google Scholar] [CrossRef]
- Capell, B.; Dorffling, K. Genotype-specific differences in chilling tolerance of maize in relation to chilling-induced changes in water status and abscisic acid accumulation. Physiol. Plant. 1993, 88, 638–646. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, K.; Welburn, A.R. Determination of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Nelson, N. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem. 1944, 153, 375–380. [Google Scholar] [CrossRef]
- Saneoka, H.; Moghaieb, R.E.A.; Premachandra, G.S.; Fujita, K. Nitrogen nutrition and water stress effects on cell membrane stability and leaf water relations in Agrostis palustris Huds. Environ. Exp. Bot. 2004, 52, 131–138. [Google Scholar] [CrossRef]
- Urbanek, H.; Kuzniak-Gebarowska, E.; Herka, K. Elicitation of Defence Responses in Bean Leaves by Botrytis cinerea Polygalacturonase. Acta Physiol. Plant. 1991, 13, 43–50. [Google Scholar]
- Gauillard, F.; Richardforget, F.; Nicolas, J. New Spectrophotometric Assay for Polyphenol Oxidase Activity. Anal. Biochem. 1993, 215, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Nagarathna, K.C.; Shetty, S.A.; Shetty, H.S. Phenylalanine Ammonia Lyase Activity in Pearl Millet Seedlings and its Relation to Downy Mildew Disease Resistance. J. Exp. Bot. 1993, 265, 1291–1296. [Google Scholar] [CrossRef]
- Mallick, C.P.; Singh, M.B. Phenolics. In Plant Enzymology and Histo Enzymology; Kalyani Publishers: New Delhi, India, 1980. [Google Scholar]
- Dickerson, D.P.; Pascholati, S.E.; Hagerman, A.E.; Butler, L.G.; Nicholson, R.L. Phenylalanine Ammonia-Lyase and Hydroxycinnamate: Mesocotyls Inoculated with Helminthosporium maydis or Helminthosporium carbonum. Physiol. Plant Pathol. 1984, 25, 111–123. [Google Scholar] [CrossRef]
- Maehly, A.C.; Chance, B. The Assay of Catalases and Peroxidases. Methods Biochem. Anal. 1954, 1, 357–424. [Google Scholar]
- Boumaaza, B.; Benada, M.H.; Boudalia, S.; Benzohra, I.E.; Gacemi, A.; Khaladi, O.; Benkhelifa, M. Effect of salinity levels on antifungal activity of essential oil from Thymus against Fusarium oxysporum. Rev. Fac. Agron. (LUZ) 2022, 39, e223941. [Google Scholar] [CrossRef]
- Tawfik, A.; Koriem, A.; Younis, S.; Elian, M. Effect of some salts on the mycelial growth and spore germination of fungi caused fruit rot of sweet pepper post-harvest diseases pathogens. J. Prod. Dev. 2021, 26, 433–446. [Google Scholar] [CrossRef]
- Boumaaza, B.; Benkhelifa, M.; Belkhoudja, M. Effects of two salts compounds on mycelial growth, sporulation, and spore germination of six isolates of Botrytis cinerea in the Western North of Algeria. Int. J. Microbiol. 2015, 2015, 572626. [Google Scholar] [CrossRef]
- Gunde-Cimerman, N.; Plemenitaš, A.; Oren, A. Strategies of adaptation of microorganisms of the three domains of life to high salt concentrations. FEMS Microbiol. Rev. 2018, 42, 353–375. [Google Scholar] [CrossRef]
- Nath, A. Insights into the sequence parameters for halophilic adaptation. Amino Acids 2016, 48, 751–762. [Google Scholar] [CrossRef]
- Elmajdoub, B.; Marschner, P. Responses of Soil Microbial Activity and Biomass to Salinity After Repeated Additions of Plant Residues. Pedosphere 2015, 25, 177–185. [Google Scholar] [CrossRef]
- Sasaki, K.; Nakahara, K.; Tanaka, S.; Shigyo, M.; Ito, S.I. Genetic and pathogenic variability of Fusarium oxysporum f. sp. cepae Isolated from onion and Welsh onion in JP. Phytopathology 2015, 105, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Gull, A.; Lone, A.; Wani, N.I. Biotic and Abiotic Stresses in Plants. Abiotic Biot. Stress Plants 2019, 7, 1–9. [Google Scholar]
- Gad, N.; Hassan, N.M.; Sayed, S.A.A.E. Influence of Cobalt on Tolerating Climatic Change (Salinity) in Onion Plant with Reference to Physiological and Chemical Approach. Plant Arch. 2020, 20, 1496–1500. [Google Scholar]
- Palma, J.M.; Sandalio, L.M.; Corpas, F.J.; Romero-Puertas, M.C.; McCarthy, I.; del Río, L.A. Plant proteases, protein degradation, and oxidative stress: Role of peroxisomes. Plant Physiol. Biochem. 2002, 40, 521–530. [Google Scholar] [CrossRef]
- Mansha, M.Z.; Habib, A.; Ashraf, W.; Shakeel, Q.; Raheel, M.; Zaman, Q.; Aatif, H.M.; Tahir, M. Impact of Resistance Inducers on Biochemical Attributes of Onion Leaves against Purple Blotch (Alternaria porri). Appl. Ecol. Environ. Res. 2019, 17, 9773–9784. [Google Scholar] [CrossRef]
- Pan, C.; Yang, K.; Erhunmwunsee, F.; Li, Y.; Liu, M.; Pan, S.; Yang, D.; Lu, G.; Ma, D.; Tian, J. Inhibitory effect of cinnamaldehyde on Fusarium solani and its application in postharvest preservation of sweet potato. Food Chem. 2023, 408, 135213. [Google Scholar] [CrossRef]
- Wang, L.; Li, X.; Gao, F.; Liu, Y.; Lang, S.; Wang, C.; Zhang, D. Effect of ultrasound combined with exogenous GABA treatment on polyphenolic metabolites and antioxidant activity of mung bean during germination. Ultrason. Sonochemistry 2023, 94, 106311. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).