Abstract
Gibberellins (GAs) are plant hormones indispensable in regulating the growth and development of fruits. Recent studies have shown that GAs play important roles in delaying horticultural crop ripening and senescence, enhancing the internal and external quality of horticultural crops and resistance to stress and disease. We reviewed the role of GAs in the postharvest physiology of fruits in recent years. GAs are closely related to their ability to retard fruit senescence. GAs could effectively improve fruit storage quality and significantly increase flesh hardness, reduce respiration intensity, inhibit the release of endogenous ethylene, and effectively inhibit fruit softening and ripening. It can also improve the intrinsic and extrinsic quality of fruit storage by improving fruit shape, regulating color, delaying the reduction of soluble solids, promoting sugar accumulation, and delaying vitamin loss. GAs also play a role in postharvest biotic and abiotic stress resistance. The GA treatment effectively reduces the cold damage index, reduces the production and accumulation of superoxide anion(O2−), improves the antioxidant capacity of fruits, and maintains the integrity of cell membranes during low-temperature storage. Moreover, GAs could effectively control some postharvest fruit diseases. In conclusion, GAs play an important role in the physiological regulation of postharvest fruits and have important application prospects in postharvest fruits.
1. Introduction
Bioactive gibberellin (GA) is one kind of phytohormone that regulates plant growth and development. GAs are involved in plant reproductive development, promoting organ growth (enhancing cell elongation and cell division), and activating developmental processes such as seed germination, maturation, and induction of flowering [1]. In recent years, the regulation mechanism of GAs and the application of exogenous plant hormones in the postharvest storage quality of horticultural crops has been studied. GAs were first found in the metabolites of the fungus Gibberella fujikuroi and later reclassified as Fusarium fujikuroi. GA was discovered as a natural plant hormone in the late 1950s that promoted growth in higherplants [2]. GAs are in a class of phytohormones with kaurene as their backbone. GAs contain a large group of diterpenoid carboxylic acids, which are classified based on their structure.
The biologically active GAs mainly include GA1, GA3, GA4, and GA7, which play direct or relative roles in plants. Among them, GA1 is the most widespread and is present in most plants [3], while GA4 is mainly found in Arabidopsis thaliana and some cucurbits. As for the relative roles of GA1 and GA4 or GA3 and GA7, further studies are needed to clarify them. At the same time, other types of GAs that are not biologically active in plants can often be converted into biologically active GAs by certain means. For example, GA12 is converted to GA4 by GA 20-oxidase (GA20ox) and GA3ox, and it can also be converted to intermediate GA53 by GA13ox and then to biologically active GA1, GA3, and GA6 by different pathways in the presence of GA20ox and GA3ox [4].
The current understanding of the physiological roles of gibberellins in postharvest horticulture crops was outlined and discussed. The main content is as follows: (1) GAs are closely related to their ability to retard fruit senescence. GA treatment could improve fruit storage quality and significantly increase flesh hardness, reduce respiration intensity, inhibit the release of endogenous ethylene, and effectively inhibit fruit softening and ripening. (2) They can also improve the intrinsic and extrinsic quality of fruit storage by improving fruit shape, regulating color, delaying the reduction of soluble solids, promoting sugar accumulation, and delaying vitamin C loss. (3) GAs improve fruit chilling resistance through effective control of postharvest chilling damage. The treatment of GAS effectively reduces the cold damage index, reducing the production and accumulation of superoxide anion (O2−), improving the antioxidant capacity of fruits, and maintaining the integrity of cell membranes during low-temperature storage. (4) GAs also a play role in postharvest disease control. Overall, GAs, as an important class of plant hormone, play an essential role in the postharvest physiological regulation of fruits. Their application in postharvest could provide valid biotechnology for extending the storage period of fruits.
2. Gibberellins for Regulating the Ripening and Senescence Process
Gibberellins could influence the ripening of fruit by affecting their size, color, flavor, and nutritional value (Table 1). Ripening is generally accomplished when fruits’ internal and external components reach optimal states. It has been found that GA3 affected the size, metabolic contents, and color of fruits during tomato softening [5,6]. Meanwhile, exogenous GA treatment could improve the nutritional traits of bulbs in most garlic cultivars, and it regulates starch synthesis and degradation by affecting related enzyme activities, promoting sucrose accumulation and inhibiting starch regeneration to accumulate soluble sugars in potato tubers [7,8]. Moreover, it has been found that applying exogenous GA3 and paclobutrazol (PAC, an inhibiter of gibberellin biosynthesis) could prolong and shorten fruit ripening time, respectively [9]. Those results suggested that GAs may exert a delayed influence on fruit ripening.

Table 1.
Effect of GAs for regulating the ripening and senescence of horticultural crops.
Regarding the molecular mechanisms of exogenous gibberellins in fruit ripening, studies have identified the effects of GAs on ripening characteristics and the expression of ethylene biosynthesis and signaling genes in tomato fruit (Figure 1). It was found that the exogenous GA treatment could effectively increase the expression level of gibberellin-stimulated transcript 1 (GAST1), slow down (or inhibit) the decrease of endogenous GA concentration, inhibit the expression of fruit ripening regulators RIPENING INHIBITOR (RIN), NON-RIPENING (NOR), and COLORLESS NON-RIPENING (CNR), attenuate the expression levels of crucial genes in ethylene synthesis (ACS2, ACS4, and ACO1), and also reduce the expression of key ethylene receptor genes EYR3 (NR) and ETR4. The altered expression of these maturation regulators also suppressed the expression of the major ethylene signaling gene EIN2 to a certain extent [6]. GAs were demonstrated to play a negative role in tomato fruit ripening by regulating the ethylene-related pathways.
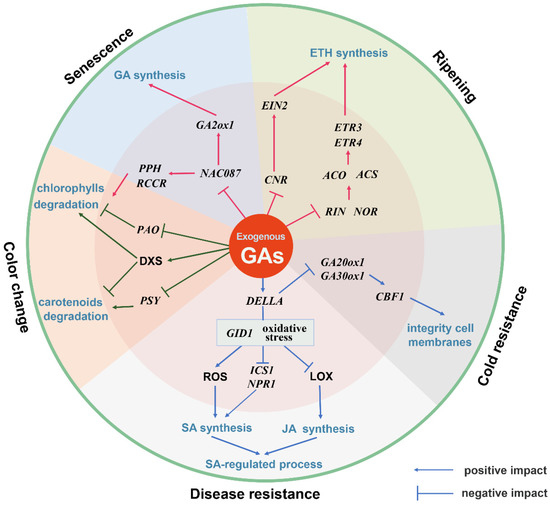
Figure 1.
Molecular mechanisms of gibberellins related to regulating postharvest physiology. (1) GAs regulated the ripening process (modeling based on tomato): GAs affect the expression of EYR3, ETR4, and EIN2 by regulating RIN, NOR, and CNR. (2) GAs regulated the senescence process (modeling based on cabbage): GAs affect the expression levels of PPH, RCCR, and GA2ox1 by regulating the expression of BrNAC087. (3) GAs regulated the color change of fruit (modeling based on citrus): the use of GAs can regulate DXS activity and gene expression of PAO and PSY. (4) GAs regulated the cold resistance of fruit (modeling based on tomato): GAs regulate GA20ox1 and GA3ox1 expression by affecting the DELLA gene expression, which in turn incrementally affects CFB1 expression. (5) GAs regulated the disease resistance of fruit (modeling based on tomato): GAs can affect DELLA gene expression through both binding to GID1 and induced oxidative stress, which in turn regulates the levels of ROS, LOX, and ICS1, NPR1 to affect SA and JA biosynthesis and thus the SAR process. Abbreviations: ACO (1-aminocyclopropane-1-carboxylic acid oxidase), ACS (1-aminocyclopropane-1-carboxylic acid synthase), CBF1 (C-repeat binding transcription factor 1), CNR (COLORLESS NON-RIPENING), DXS (1-deoxy-D-xylulose 5-phosphate synthase), EIN2 (ETHYLENE INSENSITIVE 2), ETH(ethylene content), ETR (ETHYLENE RESPONSE), GA (gibberellin), GID1 (GIBBERELLIN INSENSITIVE DWARF 1), ICS1 (isochorismate synthase 1), JA (jasmonate), LOX (lysyl oxidase), NOR (NON-RIPENING), NPR1 (nonexpressor of pathogenesis-related genes 1), PAO (pheophorbidea oxygenase), PPH (pheophytinase), PSY (phytoene synthase), RIN (RIPENING INHIBITOR), SA (salicylic acid), RCCR (red chlorophyll catabolite reductase), ROS (reactive oxygen species).
Gibberellins had important roles in regulating fruit senescence. Aging fruit refers to the gradual loss of water, flavor, and nutrition; it also reduces enzyme activity, causes cell aging, and occurs with decay and spoilage. The physiological indicators of fruit aging include enzyme activities of superoxide dismutase (SOD), the function of scavenging reactive oxygen species and is a protective enzyme for plant cells [12], peroxidase (POD), and catalase (CAT), the function of decomposing H2O2 [13], polyphenol oxidase (PPO), trigger fruit browning phenolic substances [14], and malonaldehyde (MDA), and are considered to measure the rate of lipid peroxidation in the cell membrane and be an indicator of the amount of senescence and stress [15]. It was found that GA3 treatment could significantly reduce the accumulation of MDA and H2O2, enhance the activity of CAT and SOD, and reduce the activity of POD and PPO in the shoots of Toona sinensis, which showed that GA treatment could effectively delay the aging process of Toona [11].
Molecular mechanisms related to gibberellin delaying plant aging have also been reported. A study identified BrNAC087 (a homolog of ANAC087, senescence-associated NAC transcription factor) as an important regulator of postharvest Chinese flowering cabbage leaf senescence (Figure 1). It was demonstrated that BrNAC087 acted as a direct activator of two chlorophyll catabolic genes (BrPPH and BrRCCR) and one GAs inactivation gene BrGA2ox1, by specifically binding to the promoters of BrPPH, BrRCCR, and BrGA2ox1. Exogenous GA3 treatment could effectively inhibit the expression of BrNAC087 and reduce the expression levels of BrPPH, BrRCCR, and BrGA2ox1, thus reducing chlorophyll degradation and GAs catabolism and achieving the purpose of delaying leaf senescence [10].
3. Gibberellins for Exterior Quality Control of Horticultural Crops
Gibberellins could influence fruit appearance by regulating cell expansion and enlargement, enhancing fruit stalk length, controlling fruit shape, adjusting pigment content, and modulating fruit color (Table 2). It has been shown that the fruit shape index (the ratio of the longitudinal diameter to the transverse diameter of the fruit and a quality indicator for commercial fruits) was influenced by GAs [16]. Cruz-Castillo et al. showed that GA3 supplied to the pedicel of ‘Hayward’ kiwifruit increased the length of the terminal pedicel and affected the fruit shape index [17]. Liu et al. found that GA3 treatment could significantly increase apple fruit stalk length and improve fruit shape [18]. Another study on ‘Chandler’ strawberry showed similar experimental results, which found that GA3 and cytokinin (substances with cell division activity) could promote cell elongation and thus achieve an elongation of fruit stalk length to improve the fruit shape index [19]. It has been found that GA3 treatment in tomato fruit can promote cell expansion and enlargement, thus improving the shape index [9]. GA3 spraying on self-pollinated apple plants increased fruit weight, reduced the proportion of asymmetric fruits, and restored the shape and quality of external fruits to the level of cross-pollinated fruits [18]. It is worth noting that, at the molecular level, GA-related genes could regulate fruit exterior shape during fruit development. It was claimed that, in cucumber, the expression pattern of the GA receptor gene CsGID1a was closely related to the formation of fruit form. Silencing the CsGID1a gene resulted in fruits exhibiting an abnormal carpel and ovary phenotype, while overexpression of CsGID1a in the Arabidopsis double mutant (gid1a and gid1c) exhibited ‘cucumber ovary-like’ fruits [20].
Table 2.
Effect of GAs on regulating the exterior quality of horticultural crops.
Gibberellins could modulate fruit color changes by influencing the content of pigments such as chlorophylls, carotenoids, and anthocyanins. It was also found that GA3 treatment on broccoli could control chlorophyll metabolism and delay yellowing [21]. GA3 treatment on ‘Valencia’ orange induced chlorophyll accumulation and reduced carotenoid contents (β-astaxanthin, all-trans-violaxanthin, and 9-cis-violaxanthin) [27]. It has also been shown that GA3 treatment could inhibit the degradation of anthocyanins to suppress lychee browning [26]. The treatment of GA3 on grapes reduced the flavonol and anthocyanin content and also slowed down the color change of the skin [25]. In addition, Prohexadion-calcium (ProCa, an inhibitor of GA biosynthesis with growth retarding activity) was shown to reduce chlorophyll and increase carotenoid concentrations in the yellow matter of citrus fruits [22]. Similarly, GA biosynthesis inhibitor polycarbazole (PBZ) was found to exert a similar effect in citrus fruit color conversion [23].
It was further suggested that gibberellins successfully affect the accumulation of various pigments in the fruit by regulating the activities of various enzymes in the fruit. Exogenous GA3 delayed strawberry fruit color change by inhibiting the increase in phenylalanine ammonia-lyase (PAL) activity and delaying the decrease in chlorophyllase and peroxidase (enzymes that may be involved in chlorophyll metabolism) activity (Figure 1). Similar results have been found in studies on the mechanism of color break regulation in citrus fruits, where the use of exogenous GAs delayed the reduction of 1-deoxy-D-xylulose 5-phosphate synthase (DXS) activity, while the increasing gene expression of pheophorbidea oxygenase (PAO) and phytoene synthase (PSY) increased the content of their corresponding transcripts, and delayed the consumption of pericarp chlorophyll and the accumulation of total carotenoids and retarded fruit color change [24].
4. Gibberellins for Improving the Internal Quality of Horticultural Crops
Gibberellins could influence taste and texture quality, such as fruit hardness, water loss, soluble solids, acids, and nutrient contents (Table 3). It was demonstrated that the GA3 treatment of kiwifruit could effectively delay the decrease of fruit hardness, maintain the total acid content, and accumulation of soluble solids [28]. Moreover, several studies have also shown that GA3 treatment effectively delays postharvest fruit weight loss and decay of the ‘Angelino’ plum [29] and Japanese plum varieties of ‘Obilnaja’ and ‘BlackStar’ [30]. Meanwhile, Ozturk et al. also found that the application of GA3 with CaCl2 significantly delayed the weight loss of sweet cherry [31]. These findings further support the fact that GAs are effective in delaying the aging of fruits. GA3 treatment of plum fruit increased the soluble solids content and reduced the titratable acidity at harvest, which could effectively maintain the fruit quality during storage [32]. Zang et al. found that the soluble solids content increased in GA3-treated ‘rabbit eye’ blueberry, thus improving the fresh eating quality of ‘rabbit eye’ blueberry [33]. A similar phenomenon in sweet cherry cultivars was that GA3 treatment significantly increased acidity levels [34]. Meanwhile, it was also found that treating the Japanese plum with GA3 effectively controlled the degradation of soluble solids and titratable acids [35]. Similar results have been found in apples and mangoes [16]. In addition, the water content in fruits treated with GAs has been varied, and Forchlorfenuron (CPPU) and GA3 have been reported to significantly reduce water loss in banana and broccoli [36].
Gibberellins could also influence the nutrient contents of vitamins, phenols, and soluble proteins. ‘Barhee’ dates were sprayed with different rates of GAs which effectively controlled the degradation of the vitamin C (Vc) content in the fruit [37]. Li et al. dipped the fruits of plum varieties into GA3 solution after harvest and found that the application of GA3 to plums significantly delayed the decrease in ascorbic acid concentration and the increase in total phenolic content [38]. Meanwhile, studies on fruits such as ‘rabbiteye’ blueberry, Japanese plum, and ‘nanguo’ pear fruit have shown that GA3 treatment can delay the reduction of ascorbic acid concentration and total phenolic content in the fruits, maintaining the postharvest fruit quality [33,35,39]. Besides, the results of some studies showed that the spraying of different concentrations of GA3 significantly improved the quality of garlic and Welsh onion; their soluble protein content was significantly higher [7,40]. It was also found that the phenolic content of fruit was significantly higher after GA3 spraying on ‘Cabernet Sauvignon’ grapes [41].
Table 3.
Effect of GAs on regulating the internal quality of horticultural crops.
Table 3.
Effect of GAs on regulating the internal quality of horticultural crops.
| Horticultural Varieties | Main Points | Concentration of GAs | References |
|---|---|---|---|
| Apple (Malus pumila Mill.) | GA3 spraying retards the decrease in soluble solids content in fruits. | 100 mg L−1 GA3 | Liu et al., 2022 [18] |
| Banana (Musa nana Lour.) | GA3 and CPPU treatments reduce water loss in fruits. | 50 mg L−1 GA3 + 10 mg L−1 CPPU | Huang et al., 2012 [36] |
| ‘rabbit eye’ blueberry (Vaccinium virgatum Ait.) | GA3 spraying retards the decrease in soluble solids content of fruits. | 500 mg L−1 GA3 | Zang et al., 2016 [33] |
| ‘rabbit eye’ blueberry (Vaccinium virgatum Ait.) | GA3 spraying retards the decline of ascorbic acid concentration and the increase of total phenolic content of fruits. | 500 mg L−1 GA3 | Zang et al., 2016 [33] |
| Broccoli (Brassica capitata var. italica) | GA3 and CPPU treatments reduce water loss. | 50 mg L−1 GA3 + 10 mg L−1 CPPU | Huang et al., 2012 [36] |
| Sweet cherry (Cerasus pseudocerasus (Lindl.) G. Don) | GA3 spraying retards fruit weight loss. | 30 mg L−1 GA3 | Burhan et al., 2022 [31] |
| Sweet cherry (Cerasus pseudocerasus (Lindl.) G. Don) | GA3 spraying raises fruit acidity levels. | 60 mg L−1 GA3 | Ozkan et al., 2016 [34] |
| ‘Barhee’ date (Ziziphus jujuba Mill.) | GA3 spraying retards the decline of Vc content in fruits. | 50 ppm GA3 | Awad et al., 2012 [37] |
| Garlic (Allium sativum L.) | GA3 injection increases soluble protein content of fruits. | 1 mM GA3 | Liu et al., 2019 [7] |
| ‘Cabernet Sauvignon’ grape (Vitis vinifera L.) | GA3 spraying increases phenol content of fruits. | 15 mg L−1 GA3 | Song et al., 2023 [41] |
| Kiwifruit (Actinidia chinensis Planch.) | GA3 spraying retards the decline of fruit hardness and soluble solids content. | 0.3 g L−1 GA3 | Yang et al., 2023 [28] |
| Welsh onion (Allium cepa L.) | GA3 spraying increases soluble protein content of fruits. | 40 ppm GA3 | Yamazaki et al., 2015 [40] |
| ‘Nanguo’ pear (Pyrus spp) | GA3 soaking retards the decline of ascorbic acid concentration and the increase of total phenolic content of fruits. | 100 mg L−1 GA3 | Martínez-Romero et al., 2000 [39] |
| Plum (Prunus salicina Lindl.) | GA3 spraying retards the decrease in soluble solids content of fruits and reduces titratable acidity at harvest. | 200 mg L−1 GA3 | Barac et al., 2022 [32] |
| Plum (Prunus salicina Lindl.) | GA3 spraying retards the reduction of soluble solids and titratable acid content in fruits. | 50 mg L−1 GA3 | Erogul et al., 2015 [35] |
| Plum (Prunus salicina Lindl.) | GA3 soaking retards the decline of ascorbic acid concentration and the increase of total phenolic content of fruits. | 200 μL L−1 GA3 | Li et al., 2006 [38] |
| Plum (Prunus salicina Lindl.) | GA3 spraying retards the decline of ascorbic acid concentration and the increase of total phenolic content of fruits. | 50 mg L−1 GA3 | Erogul et al., 2015 [35] |
| ‘Angelino’ plum (Prunus salicina Lindl.) | GA3 spraying retards fruit weight loss and decay. | 50 mg L−1 GA3 | Erogul et al., 2016 [29] |
| ‘BlackStar’ plum (Prunus salicina Lindl.) | GA3 spraying retards fruit weight loss and decay. | 50 ppm GA3 | Harman et al., 2016 [30] |
| ‘Obilnaja’ plum (Prunus salicina Lindl.) | GA3 spraying retards fruit weight loss and decay. | 50 ppm GA3 | Harman et al., 2016 [30] |
5. Gibberellins for Improving Cold Resistance in Horticultural Crops
Gibberellins participate in plant responses to biotic and abiotic stresses. When fruits are subjected to postharvest stress, the exogenous application of GAs can effectively regulate biotic and abiotic stress resistance. Cold damage is one of the more important abiotic stresses in the postharvest aspect of fruits, and it is a major constraint to the low-temperature storage of cold-sensitive fruits, resulting in sunken skins, watery spots, dry scars, flesh flocculation or lignification, reduced flavor, failure to properly ripen, and accelerated decay. It was found that elevated GA3 levels were accompanied by the down-regulation of DELLA genes (members of the GRAS protein family that repress GA responses, degraded by the 26S-proteasome upon interaction with the GID1-GA complex) and the expression of key GA biosynthetic genes, GA20ox1 and GA3ox1, was down-regulated in the application of exogenous GA3 (Figure 1). Zhu et al. found that GA3 treatment reduced the cold damage index, maintained the integrity of cell membranes during low-temperature storage, and activated the feedback mechanism of GA anabolism and the expression of C-repeat binding transcription factor 1 (CBF1), an important regulator of cold resistance, thus effectively improving the cold resistance of tomato fruits [42]. Ding et al. showed that, during low-temperature storage of the GA-deficient mutant gib-3 tomato fruit, the cold damage index was significantly higher than that of normal fruit, and the breakage of microstructures such as cell membranes and cell walls was accelerated and more severe. GA deficiency also led to an up-regulation of GA3ox1 and a significant down-regulation of CFB1, indicating that GA deficiency reduced the cold resistance of the fruit [43]. Ding et al. treated cherry tomato with GA3 and stored them at low temperatures; the fruit cold damage index decreased significantly, effectively maintaining the stability of the cell membrane during storage, reducing the production and accumulation of O2− and improving the antioxidant capacity of the fruit. The results indicated that GA treatment effectively controlled postharvest cold damage in the fruit [44] (Table 4).
Table 4.
Effect of GAs on regulating cold resistance of horticultural crops.
6. Gibberellins Regulate Disease Resistance in Horticultural Crops
Research has indicated that gibberellin treatment effectively controlled the incidence of disease during storage. Disease is a series of morphological, physiological, and biochemical pathological changes in fruits under the influence of biotic or abiotic factors. In the horticultural postharvest sector, this mainly manifests as the phenomenon of fruit that are healthy at harvest but infected with disease afterward, and as fruits that are apparently intact but diseased at harvest and only show traces of disease after harvest. Trichothecium roseum [45] and Fusarium [46] are susceptible to postharvest fruit infestation, causing pink mold and white mold, which cause extensive fruit rot. Ma et al. found that GA3 did not inhibit the growth of Trichothecium roseum and Fusarium, but when mixed with azoxystrobin (AZX), the spot area of both diseases was reduced to different degrees compared to the two alone, thus inferring that GA3 improved fruit resistance by regulating the physiological activity of postharvest fruits [47]. Black skin disease is a major physiological disease that occurs on pear fruit after harvesting, producing irregular black-brown spots of varying sizes on the surface of the fruit skin, which seriously reduces the quality of the appearance of the fruit [48]. Ma et al. treated ‘apple pear’ fruit with GA3 and found that POD, CAT, APX, and SOD activities increased, while PPO activity decreased and the thickness of surface protective tissue increased; the results indicated that GA treatment effectively controlled ‘apple pear’ black skin disease [49]. In addition, in studies on persimmon’s black spots, GAs were found to reduce rot by inhibiting the extracellular enzyme activity of Alternaria alternata (A. alternate) to maintain fruit cell wall integrity and limit pathogen expansion and access to host nutrients [50]. Similarly, one study found that GAs use reduced fruit cuticle dehiscence and susceptibility to persimmon fruit black spot disease during late fruit growth and storage. The present results suggest that GAs cause delayed fruit ripening and are a major factor in enhancing resistance to persimmon fruit black spots during harvest and storage [51] (Table 5).
Table 5.
Effect of GAs on regulating disease resistance of horticultural crops.
The molecular mechanisms of gibberellin to regulate disease resistance in fruits were studied. DELLA regulates the balance of salicylic acid (SA)/jasmonate (JA) signaling during plant immunity [53]. SA and JA play key roles in the induction of the systemic acquired resistance (SAR) process for defense against pathogenic bacteria infestation, while isochorismate synthase 1 (ICS1) transcription activates SA biosynthesis; nonexpressor of pathogenesis-related genes 1 (NPR1) is an important regulator of the SA-regulated SAR process [54] (Figure 1). Early blight caused by Streptomyces aegypti infestation is one of the most common postharvest fungal diseases [55]. A study indicated that ICS1 and NPR1 were up-regulated, PAL, PPO, CHI, and GLU activities were significantly enhanced, brassinolide (BR) content was increased, fruit incidence was significantly reduced, and spot area was significantly decreased in the GA-deficient mutant strain gib-3 tomato fruit, indicating that GAs negatively regulated postharvest tomato fruit resistance to early blight [52].
7. Conclusions
In conclusion, as important plant hormones, gibberellins play a significant role in the postharvest physiological regulation of fruits. During the storage of fruits, GAs can effectively delay the respiratory leap in some fruits and can delay the weight loss and decay of fruits by inhibiting ethylene production, reducing respiration intensity, scavenging free radicals in the body to maintain cell membrane integrity, and delaying the senescence of fruit stalks to prevent them from falling off. They can also improve the intrinsic and extrinsic quality of fruit storage by improving fruit shape, regulating color, and delaying the reduction of soluble solids. GAs also play an important role in enhancing postharvest stress resistance in fruits. This review systematically described the role of GAs in the physiological regulation of postharvest fruits and their existing applications, providing a more comprehensive perspective to further enhance the scope of their application in postharvest fruits and to clarify their usage in practice. At the same time, this paper lays a certain foundation for accelerating and improving the research on the physiological aspects of the regulation of GAs in postharvest fruits, and also points to a certain direction for in-depth molecular-level mechanism research.
Author Contributions
J.Z., Y.C., X.R. and Y.D. conceived and wrote the manuscript. Y.D. and X.R. provided financial support; J.T., X.H., M.L. and C.L. collected information and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Key S&T Special Projects of Shaanxi Province, China (2020zdzx03-05-01); Natural Science Basic Research Program of Shaanxi (2022JQ-178); Primary Research and Development Plan of Ningxia Hui Autonomous Region (2021BBF0214); and Modern Agricultural Industry Technology System of China (Z225020701).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare that there is no conflict of interest regarding the publication of this article.
References
- Binenbaum, J.; Weinstain, R.; Shani, E. Gibberellin Localization and Transport in Plants. Trends Plant Sci. 2018, 23, 410–421. [Google Scholar] [CrossRef] [PubMed]
- MacMillan, J.; Suter, P.J. The occurrence of gibberellin A1 in higher plants: Isolation from the seed of runner bean (Phaseolus multiflorus). Naturwissenschaften 1958, 45, 46. [Google Scholar] [CrossRef]
- MacMillan, J. Occurrence of Gibberellins in Vascular Plants, Fungi, and Bacteria. J. Plant Growth Regul. 2001, 20, 387–442. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S. Gibberellin metabolism and its regulation. Annu. Rev. Plant Biol. 2008, 59, 225–251. [Google Scholar] [CrossRef]
- Zhu, Z.; Liang, H.; Chen, G.; Li, F.; Wang, Y.; Liao, C.; Hu, Z. The bHLH transcription factor SlPRE2 regulates tomato fruit development and modulates plant response to gibberellin. Plant Cell Rep. 2019, 38, 1053–1064. [Google Scholar] [CrossRef]
- Li, H.; Wu, H.; Qi, Q.; Li, H.; Li, Z.; Chen, S.; Ding, Q.; Wang, Q.; Yan, Z.; Gai, Y.; et al. Gibberellins Play a Role in Regulating Tomato Fruit Ripening. Plant Cell Physiol. 2019, 60, 1619–1629. [Google Scholar] [CrossRef]
- Liu, H.; Deng, R.; Huang, C.; Cheng, Z.; Meng, H. Exogenous gibberellins alter morphology and nutritional traits of garlic (Allium sativum L.) bulb. Sci. Hortic. 2019, 246, 298–306. [Google Scholar] [CrossRef]
- Xie, Y.; Onik, J.C.; Hu, X.; Duan, Y.; Lin, Q. Effects of (S)-Carvone and Gibberellin on Sugar Accumulation in Potatoes during Low Temperature Storage. Molecules 2018, 23, 3118. [Google Scholar] [CrossRef]
- Chen, S.; Wang, X.J.; Tan, G.F.; Zhou, W.Q.; Wang, G.L. Gibberellin and the plant growth retardant Paclobutrazol altered fruit shape and ripening in tomato. Protoplasma 2020, 257, 853–861. [Google Scholar] [CrossRef]
- Fan, Z.Q.; Wei, W.; Tan, X.L.; Shan, W.; Kuang, J.F.; Lu, W.J.; Su, X.G.; Lakshmanan, P.; Lin, H.T.; Chen, J.Y. A NAC transcription factor BrNAC087 is involved in gibberellin-delayed leaf senescence in Chinese flowering cabbage. Postharvest Biol. Technol. 2021, 181, 111673. [Google Scholar] [CrossRef]
- Zhao, H.; Lv, W.J.; Fan, Y.L.; Li, H.Q. Gibberellic acid enhances postharvest toon sprout tolerance to chilling stress by increasing the antioxidant capacity during the short-term cold storage. Sci. Hortic. 2018, 237, 184–191. [Google Scholar] [CrossRef]
- Zhou, G.; Liu, C.; Cheng, Y.; Ruan, M.; Ye, Q.; Wang, R.; Yao, Z.; Wan, H. Molecular Evolution and Functional Divergence of Stress-Responsive Cu/Zn Superoxide Dismutases in Plants. Int. J. Mol. Sci. 2022, 23, 7082. [Google Scholar] [CrossRef] [PubMed]
- Paul, G.K.; Mahmud, S.; Dutta, A.K.; Sarkar, S.; Laboni, A.A.; Hossain, M.S.; Nagata, A.; Karmaker, P.; Razu, M.H.; Kazi, T.; et al. Volatile compounds of Bacillus pseudomycoides induce growth and drought tolerance in wheat (Triticum aestivum L.). Sci. Rep. 2022, 12, 19137. [Google Scholar] [CrossRef]
- Silva, F.V.M.; Sulaiman, A. Control of Enzymatic Browning in Strawberry, Apple, and Pear by Physical Food Preservation Methods: Comparing Ultrasound and High-Pressure Inactivation of Polyphenoloxidase. Foods 2022, 11, 1942. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.K.; Wang, C.Y.; Gross, K.C.; Smith, D.L. Reduction of chilling injury and transcript accumulation of heat shock proteins in tomato fruit by methyl jasmonate and methyl salicylate. Plant Sci. 2001, 161, 1153–1159. [Google Scholar] [CrossRef]
- Li, J.; Cao, J.; Wang, C.; Hao, N.; Zhang, X.; Liu, M.; Wu, T. Research Progress on the Leaf Morphology, Fruit Development and Plant Architecture of the Cucumber. Plants 2022, 11, 2128. [Google Scholar] [CrossRef]
- Cruz-Castillo, J.G.; Woolley, D.J. Pedicel weight and length do not affect kiwifruit size. Eur. J. Hortic. Sci. 2006, 71, 272–276. [Google Scholar]
- Liu, C.; Xiao, P.; Jiang, F.; Wang, S.; Liu, Z.; Song, G.; Li, W.; Lv, T.; Li, J.; Wang, D.; et al. Exogenous gibberellin treatment improves fruit quality in self-pollinated apple. Plant Physiol. Biochem. 2022, 174, 11–21. [Google Scholar] [CrossRef]
- Sharma, R.R.; Singh, R. Gibberellic acid influences the production of malformed and button berries, and fruit yield and quality in strawberry (Fragaria×ananassa Duch.). Sci. Hortic. 2009, 119, 430–433. [Google Scholar] [CrossRef]
- Liu, B.; Liu, X.; Yang, S.; Chen, C.; Xue, S.; Cai, Y.; Wang, D.; Yin, S.; Gai, X.; Ren, H. Silencing of the gibberellin receptor homolog, CsGID1a, affects locule formation in cucumber (Cucumis sativus) fruit. New Phytol. 2016, 210, 551–563. [Google Scholar] [CrossRef]
- Wang, H.T.; Ou, L.Y.; Chen, T.A.; Kuan, Y.C. Refrigeration, forchlorfenuron, and gibberellic acid treatments differentially regulate chlorophyll catabolic pathway to delay yellowing of broccoli. Postharvest Biol. Technol. 2023, 197, 112221. [Google Scholar] [CrossRef]
- Barry, G.H.; Le Roux, S. Preharvest Foliar Sprays of Prohexadione-calcium, a Gibberellin-biosynthesis Inhibitor, Induce Chlorophyll Degradation and Carotenoid Synthesis in Citrus Rinds. Hortscience 2010, 45, 242–247. [Google Scholar] [CrossRef]
- Rehman, M.; Singh, Z.; Khurshid, T. Pre-harvest spray application of prohexadione-calcium and paclobutrazol improves rind colour and regulates fruit quality in M7 Navel oranges. Sci. Hortic. 2018, 234, 87–94. [Google Scholar] [CrossRef]
- Alos, E.; Cercos, M.; Rodrigo, M.J.; Zacarias, L.; Talon, M. Regulation of color break in citrus fruits. Changes in pigment profiling and gene expression induced by gibberellins and nitrate, two ripening retardants. J. Agric. Food Chem. 2006, 54, 4888–4895. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, K.; Maoz, I.; Lapidot, O.; Kochanek, B.; Butnaro, Y.; Shlisel, M.; Lerno, L.; Ebeler, S.E.; Lichter, A. Effects of gibberellin and cytokinin on phenolic and volatile composition of Sangiovese grapes. Sci. Hortic. 2022, 295, 111672. [Google Scholar] [CrossRef]
- Qu, S.S.; Li, M.M.; Wang, G.; Zhu, S.J. Application of ABA and GA3 alleviated browning of litchi (Litchi chinensis Sonn.) via different strategies. Postharvest Biol. Technol. 2021, 181, 111672. [Google Scholar] [CrossRef]
- Keawmanee, N.; Ma, G.; Zhang, L.C.; Yahata, M.; Murakami, K.; Yamamoto, M.; Kojima, N.; Kato, M. Exogenous gibberellin induced regreening through the regulation of chlorophyll and carotenoid metabolism in Valencia oranges. Plant Physiol. Biochem. 2022, 173, 14–24. [Google Scholar] [CrossRef]
- Yang, H.; Li, J.; Li, X.; Wu, R.; Zhang, X.; Fan, X.; Li, G.; Gong, H.; Yin, X.; Zhang, A. The mechanism of gibberellins treatment suppressing kiwifruit postharvest ripening processes by transcriptome analysis. Postharvest Biol. Technol. 2023, 198, 112223. [Google Scholar] [CrossRef]
- Erogul, D.; Sen, F. The effect of preharvest gibberellic acid applications on fruit quality of ʽAngelinoʼ plums during storage. Sci. Hortic. 2016, 202, 111–116. [Google Scholar] [CrossRef]
- Harman, Y.; Sen, F. The effect of different concentrations of pre-harvest gibberellic acid on the quality and durability of ‘Obilnaja’ and ‘Black Star’ plum varieties. Food Sci. Technol. 2016, 36, 362–368. [Google Scholar] [CrossRef]
- Burhan, O.; Erdal, A.; Saraçoğlu, O.; Karakaya, O.; Gun, S. Effects of GA 3, CACl 2 and Modified Atmosphere Packaging (MAP) Applications on Fruit Quality of Sweet Cherry at Cold Storage. Int. J. Fruit Sci. 2022, 22, 696–710. [Google Scholar] [CrossRef]
- Barac, G.; Mastilovic, J.; Kevresan, Z.; Milic, B.; Kovac, R.; Milovic, M.; Kalajdzic, J.; Bajic, A.; Magazin, N.; Keserovic, Z. Effects of Plant Growth Regulators on Plum (Prunus domestica L.) Grown on Two Rootstocks at Harvest and at the Postharvest Period. Horticultuae 2022, 8, 621. [Google Scholar] [CrossRef]
- Zang, Y.X.; Chun, I.J.; Zhang, L.L.; Hong, S.B.; Zheng, W.W.; Xu, K. Effect of gibberellic acid application on plant growth attributes, return bloom, and fruit quality of rabbiteye blueberry. Sci. Hortic. 2016, 200, 13–18. [Google Scholar] [CrossRef]
- Ozkan, Y.; Ucar, M.; Yildiz, K.; Ozturk, B. Pre-harvest gibberellic acid (GA3) treatments play an important role on bioactive compounds and fruit quality of sweet cherry cultivars. Sci. Hortic. 2016, 211, 358–362. [Google Scholar] [CrossRef]
- Erogul, D.; Sen, F. Effects of gibberellic acid treatments on fruit thinning and fruit quality in Japanese plum (Prunus salicina Lindl.). Sci. Hortic. 2015, 186, 137–142. [Google Scholar] [CrossRef]
- Huang, H.; Jiang, Y. Effect of plant growth regulators on banana fruit and broccoli during storage. Sci. Hortic. 2012, 145, 62–67. [Google Scholar] [CrossRef]
- Awad, M.A.; Al-Qurashi, A.D. Gibberellic acid spray and bunch bagging increase bunch weight and improve fruit quality of ‘Barhee’ date palm cultivar under hot arid conditions. Sci. Hortic. 2012, 138, 96–100. [Google Scholar] [CrossRef]
- Li, H.; Jiang, Y.; Li, J. Use of GA3 to inhibit flesh browning development of plum fruit during storage at low temperature. Eur. J. Hortic. Sci. 2006, 71, 231–235. [Google Scholar]
- Martínez-Romero, D.; Valero, D.; Serrano, M.; Burló, F.; Carbonell-Barrachina, A.; Burgos, L.; Riquelme, F. Exogenous Polyamines and Gibberellic Acid Effects on Peach (Prunus persica L.) Storability Improvement. J. Food Sci. 2000, 65, 288–294. [Google Scholar] [CrossRef]
- Yamazaki, H.; Shiraiwa, N.; Itai, A.; Honda, I. Involvement of Gibberellins in the Regulation of Tillering in Welsh Onion (Allium fistulosum L.). Hortic. J. 2015, 84, 334–341. [Google Scholar] [CrossRef]
- Song, J.Q.; Zhang, A.; Gao, F.; Liang, H.Z.; Li, M.Q.; Zhang, J.; Wang, G.J.; Qu, H.G.; Cheng, S.W.; Ruan, S.L.; et al. Modification of wine phenolic profiles by gibberellic acid application in ‘Cabernet Gernischt’ grapevines before anthesis. J. Sci. Food Agric. 2023, 103, 1216–1225. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Ding, Y.; Zhao, J.; Nie, Y.; Zhang, Y.; Sheng, J.; Tang, X. Effects of Postharvest Gibberellic Acid Treatment on Chilling Tolerance in Cold-Stored Tomato (Solanum lycopersicum L.) Fruit. Food Bioprocess Technol. 2016, 9, 1202–1209. [Google Scholar] [CrossRef]
- Ding, Y.; Zhao, J.; Nie, Y.; Fan, B.; Wu, S.; Zhang, Y.; Sheng, J.; Shen, L.; Zhao, R.; Tang, X. Salicylic-Acid-Induced Chilling- and Oxidative-Stress Tolerance in Relation to Gibberellin Homeostasis, C-Repeat/Dehydration-Responsive Element Binding Factor Pathway, and Antioxidant Enzyme Systems in Cold-Stored Tomato Fruit. J. Agric. Food Chem. 2016, 64, 8200–8206. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Sheng, J.; Li, S.; Nie, Y.; Zhao, J.; Zhu, Z.; Wang, Z.; Tang, X. The role of gibberellins in the mitigation of chilling injury in cherry tomato (Solanum lycopersicum L.) fruit. Postharvest Biol. Technol. 2015, 101, 88–95. [Google Scholar] [CrossRef]
- Gong, D.; Bi, Y.; Li, S.; Li, Y.; Wang, Y. Trichothecium roseum infection promotes ripening of harvested muskmelon fruits and induces the release of specific volatile compound. J. Plant Pathol. 2019, 101, 529–538. [Google Scholar] [CrossRef]
- Xue, H.; Bi, Y.; Sun, Y.; Hussain, R.; Wang, H.; Zhang, S.; Zhang, R.; Long, H.; Nan, M.; Cheng, X.; et al. Acetylsalicylic acid treatment reduce Fusarium rot development and neosolaniol accumulation in muskmelon fruit. Food Chem. 2019, 289, 278–284. [Google Scholar] [CrossRef]
- Ma, L.Y.; Zhao, L.; Bi, Y.; An, L. Efficacy of erythromycin and daidzein with pyrimethanil for postharvest disease control in melon. Henan Agric. Sci. 2005, 78–81. [Google Scholar] [CrossRef]
- Jia, Z.S.; Wang, Z.H.; Wang, W.H.; Jiang, Y.B. Relationship between the occurrence of black skin disease and related physiological indicators during storage of Dangshan pear fruit. North China J. Agric. 2021, 36, 203–209. [Google Scholar]
- Ma, L.; Li, H.X.; Wang, Y.; Li, Y.C.; Bi, Y. The effect of pre-harvest gibberellins on ‘black peel’ pears. Effect of pre-harvest gibberellin spray on the prevention and control of black skin disease in ‘apple pear’ during storage at room temperature. China Fruit Tree 2018, 193, 1–4+10. [Google Scholar] [CrossRef]
- Eshel, D.; Ben-Arie, R.; Dinoor, A.; Prusky, D. Resistance of Gibberellin-Treated Persimmon Fruit to Alternaria alternata Arises from the Reduced Ability of the Fungus to Produce Endo-1,4-beta-Glucanase. Phytopathology 2000, 90, 1256–1262. [Google Scholar] [CrossRef]
- Biton, E.; Kobiler, I.; Feygenberg, O.; Yaari, M.; Friedman, H.; Prusky, D. Control of alternaria black spot in persimmon fruit by a mixture of gibberellin and benzyl adenine, and its mode of action. Postharvest Biol. Technol. 2014, 94, 82–88. [Google Scholar] [CrossRef]
- Wu, S.J. Mechanism of Action of Gibberellin and Brassinolide in the Regulation of Early Blight Resistance in Postharvest Tomato Fruits. Master’s thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2018. [Google Scholar]
- De Bruyne, L.; Hofte, M.; De Vleesschauwer, D. Connecting growth and defense: The emerging roles of brassinosteroids and gibberellins in plant innate immunity. Mol. Plant 2014, 7, 943–959. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D. Salicylic acid signaling in disease resistance. Plant Sci. 2014, 228, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, P.; Oh, Y.; Panthee, D.R. Current Status of Early Blight Resistance in Tomato: An Update. Int. J. Mol. Sci. 2017, 18, 2019. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).