
Chinese Cabbage BrCAP Has Potential Resistance against Plasmodiophora brassicae

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Preparation of P. brassicae Suspension
2.3. Plant Materials Cultivation Method
2.4. Cloning of BrCAP and Bioinformatics Analysis
2.5. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
2.6. In Situ Hybridization
2.7. Subcellular Localization
2.8. Identification of Disease Resistance of A. thaliana Mutant cap to P. brassicae
3. Results
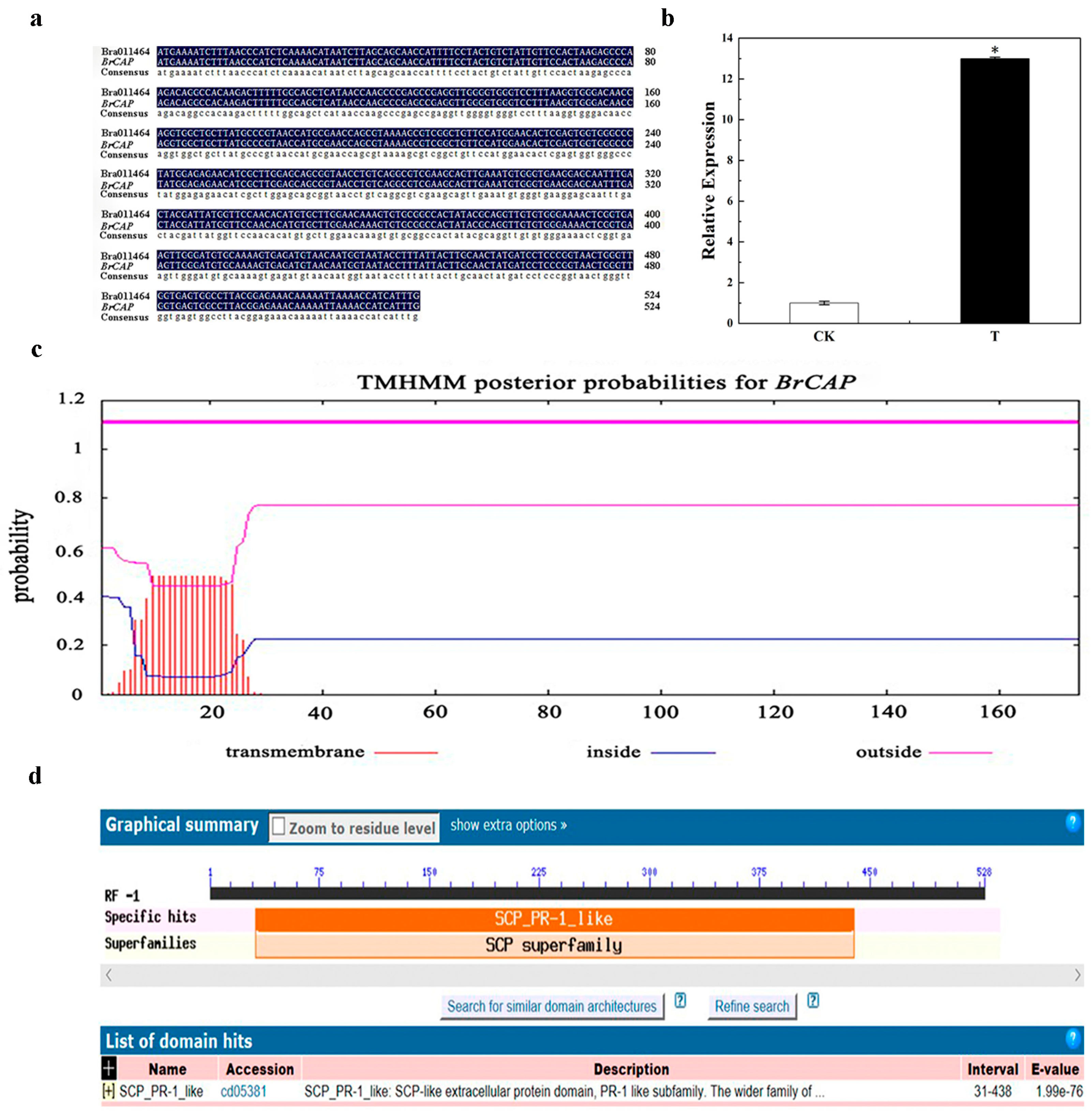
3.1. Obtainment and Prediction of BrCAP Gene
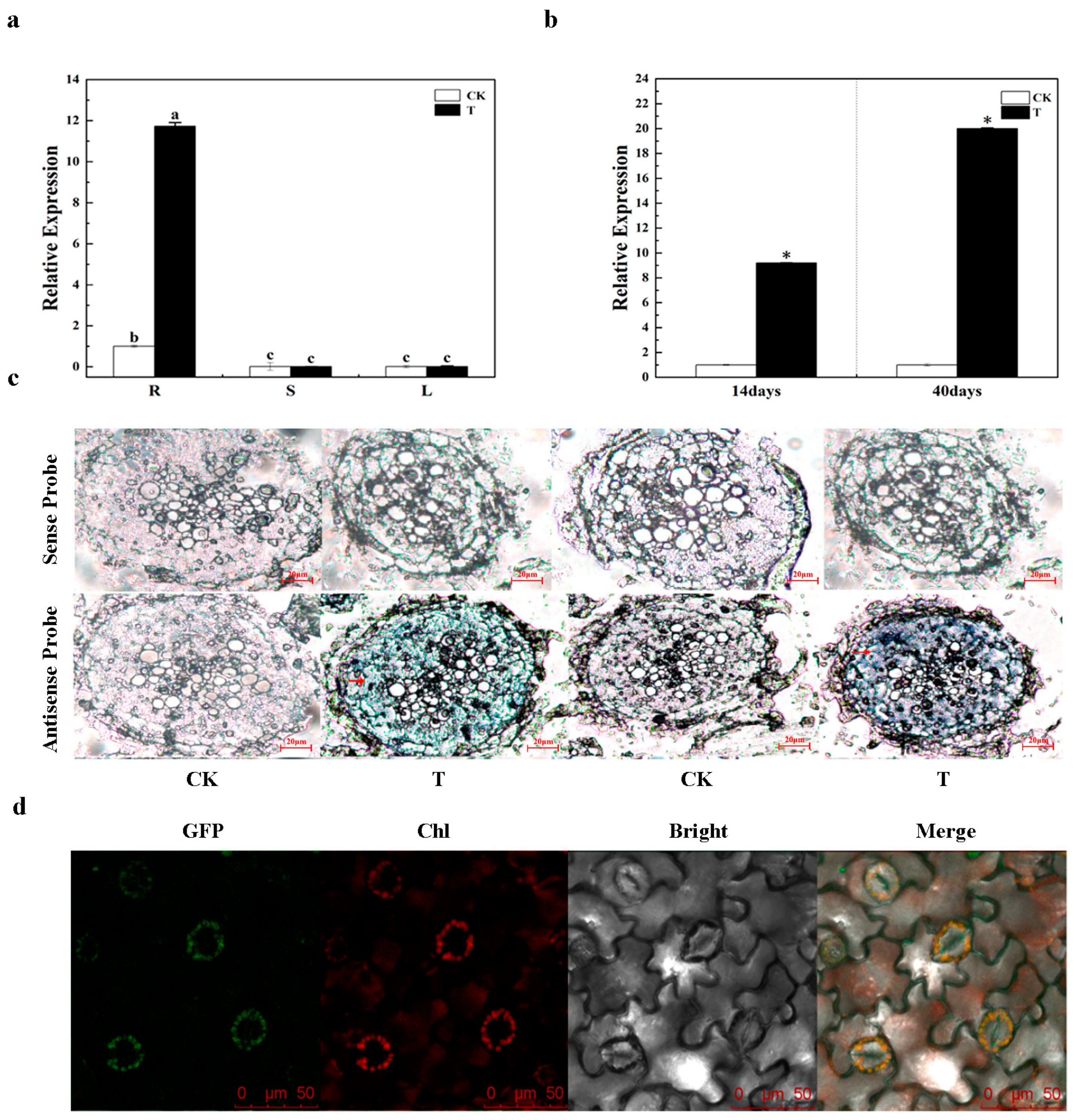
3.2. Expression Mode Analysis of BrCAP
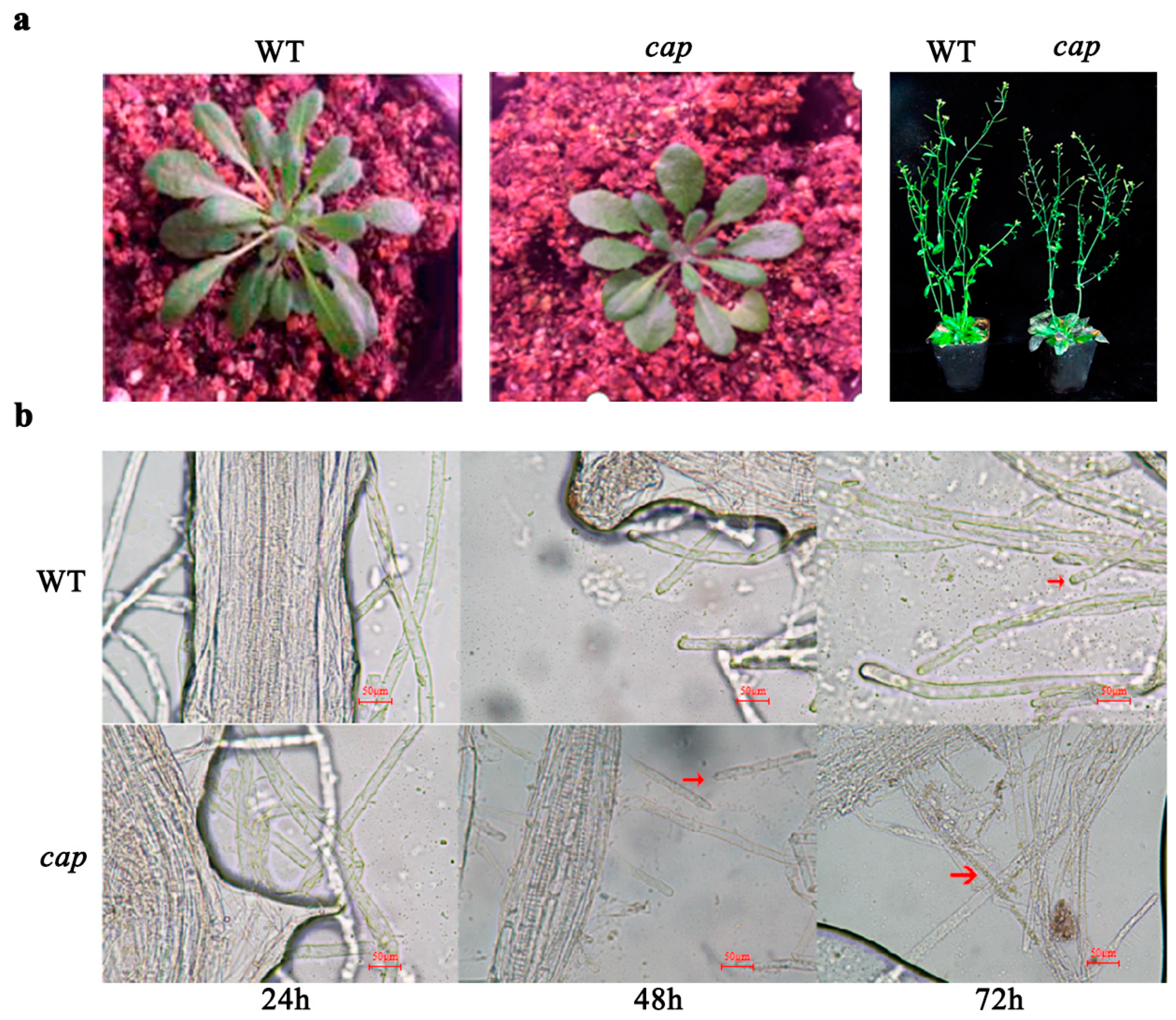
3.3. Resistance Identification of Arabidopsis Thaliana Mutant CAP
4. Discussion
4.1. The Expression of BrCAP in Chinese Cabbage Is Related to the Infection of P. brassicae
4.2. BrCAP was Localized on Chloroplasts
4.3. BrCAP May Play a Key Role in the Resistance of Chinese Cabbage against Clubroot Disease
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| P. brassicae | Plasmodiophora brassicae Woronin |
| RT-qPCR | Real-time quantitative polymerase chain reaction |
| 2-DE | two-dimensional electrophoresis |
| CAP | Cysteine-rich secreting protein, Antigen 5, and disease-related Protein 1 |
| A. thaliana | Arabidopsis thaliana |
| WT | wild type |
References
- Eom, S.H.; Baek, S.A.; Kim, J.K.; Hyun, T.K. Transcriptome analysis in Chinese cabbage (Brassica rapa ssp. pekinensis) provides the role of glucosinolate metabolism in response to drought stress. Molecules 2018, 23, 1186. [Google Scholar] [PubMed]
- Luo, H.; Chen, G.; Liu, C. An improved culture solution technique for Plasmodiophora brassicae infection and the dynamic infection in the root hair. Australas. Plant Pathol. 2014, 43, 53–60. [Google Scholar] [CrossRef]
- Li, J.; Philp, J.; Li, J.; Wei, Y.; Li, H.; Yang, K.; Ryder, M.; Toh, R.; Zhou, Y.; Denton, M.D.; et al. Trichoderma harzianum inoculation reduces the incidence of clubroot disease in Chinese cabbage by regulating the rhizosphere microbial community. Microorganisms 2020, 8, 1325. [Google Scholar] [CrossRef] [PubMed]
- Johannes, S.; Maria-Cruz, G.; Sebastian, W. Extracellular invertase is involved in the regulation of clubroot disease in Arabidopsis thaliana. Mol. Plant Pathol. 2011, 12, 247–262. [Google Scholar]
- Ahmed, H.U.; Hwang, S.F.; Strelkov, S.E.; Gossen, S.E.; Peng, G. Assessment of bait crops to reduce inoculum of clubroot (Plasmodiophora brassicae) of canola. Can. J. Plant Sci. 2011, 91, 545–551. [Google Scholar] [CrossRef]
- Hatakeyama, K.; Suwabe, K.; Tomita, R.N. Identification and characterization of Crr1a, a gene for resistance to clubroot disease (Plasmodiophora brassicae Woronin) in Brassica rapa L. PLoS ONE 2013, 8, e54745. [Google Scholar] [CrossRef]
- Luo, Y.; Dong, D.; Su, Y.; Wang, X.; Peng, Y.; Peng, J.; Zhou, C. Transcriptome analysis of Brassica juncea var. tumida tsen responses to Plasmodiophora brassicae primed by the biocontrol strain zhihengliuella aestuarii. Funct. Integr. Genom. 2018, 18, 301–314. [Google Scholar] [CrossRef]
- Daval, S.; Gazengel, K.; Belcour, A.; Linglin, J.; Guillerm-Erckelboudt, A.Y.; Sarniguet, A.; Manzanares-Dauleux, M.J.; Lebreton, L.; Mougel, C. Soil microbiota influences clubroot disease by modulating Plasmodiophora brassicae and Brassica napus transcriptomes. Microb. Biotechnol. 2020, 13, 1648–1672. [Google Scholar] [CrossRef]
- Feng, Y.; Cui, R.; Wang, S.; He, M.; Hua, Y.; Shi, L.; Ye, X.; Xu, F. Transcription factor BnaA9.WRKY47 contributes to the adaptation of Brassica napus to low boron stress by up-regulating the boric acid channel gene BnaA3.NIP5;1. Plant Biotechnol. J. 2020, 18, 1241–1254. [Google Scholar] [CrossRef]
- Chen, W.; Li, Y.; Yan, R.; Ren, L.; Liu, F.; Zeng, L.; Sun, S.; Yang, H.; Chen, K.; Xu, L.; et al. SnRK1.1-mediated resistance of Arabidopsis thaliana to clubroot disease is inhibited by the novel Plasmodiophora brassicae effector PBZF1. Mol. Plant Pathol. 2021, 22, 1057–1069. [Google Scholar] [CrossRef]
- Karim, M.M.; Dakouri, A.; Zhang, Y.; Chen, Q.; Peng, G.; Strelkov, S.E.; Gossen, B.D.; Yu, F. Two clubroot-resistance genes, Rcr3 and Rcr9wa, mapped in Brassica rapa using bulk segregant RNA sequencing. Int. J. Mol. Sci. 2020, 21, 5033. [Google Scholar] [CrossRef] [PubMed]
- Ce, F.; Mei, J.; He, H.; Zhao, Y.; Hu, W.; Yu, F.; Li, Q.; Ren, X.; Si, J.; Song, H.; et al. Identification of candidate genes for clubroot-resistance in Brassica oleracea using quantitative trait loci-sequencing. Front. Plant Sci. 2021, 12, 703520. [Google Scholar] [CrossRef] [PubMed]
- Wielkopolan, B.; Obrepalska-Steplowska, A. Three-way interaction among plants, bacteria, and coleopteran insects. Planta 2016, 244, 313–332. [Google Scholar] [CrossRef] [PubMed]
- Ryals, J.A.; Neuenschwander, U.H.; Willits, M.G.; Molina, A.; Steiner, H.Y.; Hunt, M.D. Systemic acquired resistance. Plant Cell 1996, 8, 1809–1819. [Google Scholar] [CrossRef] [PubMed]
- Nie, P.; Li, X.; Wang, S.; Guo, J.; Zhao, H.; Niu, D. Induced systemic resistance against botrytis cinerea by bacillus cereus AR156 through a JA/ET- and NPR1-dependent signaling pathway and activates PAMP-triggered immunity in Arabidopsis. Front. Plant Sci. 2017, 8, 238. [Google Scholar] [CrossRef]
- Li, T.; Huang, Y.; Xu, Z.S.; Wang, F.; Xiong, A.S. Salicylic acidinduced diferential resistance to the tomato yellow leaf curl virus among resistant and susceptible tomato cultivars. BMC Plant Biol. 2019, 19, 173. [Google Scholar] [CrossRef]
- Koornneef, A.; Pieterse, C.M. Cross talk in defense signaling. Plant Physiol. 2008, 146, 839–844. [Google Scholar] [CrossRef]
- Fang, K.S.; Vitale, M.; Fehlner, P.; King, T.P. cDNA cloning and primary structure of a white-face hornet venom allergen, antigen 5. PNAS 1988, 85, 895–899. [Google Scholar] [CrossRef]
- Chen, Y.L.; Lee, C.Y.; Cheng, K.T.; Chang, W.H.; Huang, R.N.; Nam, H.G.; Chen, Y.R. Quantitative peptidomics study reveals that a wound-induced peptide from PR-1 regulates immune signaling in tomato. Plant Cell 2014, 26, 4135–4148. [Google Scholar] [CrossRef]
- Fizames, C.; Muños, S.; Cazettes, C.; Nacry, P.; Boucherez, J.; Gaymard, F.; Piquemal, D.; Delorme, V.; Commes, T.; Doumas, P.; et al. The Arabidopsis root transcriptome by serial analysis of gene expression. Plant Pathol. 2004, 134, 67–80. [Google Scholar]
- Ambrosone, A.; Batelli, G.; Nurcato, R.; Aurilia, V.; Punzo, P.; Bangarusamy, D.K. The Arabidopsis RNA-binding protein AtRGGA regulates tolerance to salt and drought stress. Plant Physiol. 2015, 168, 292–306. [Google Scholar] [CrossRef] [PubMed]
- Harshavardhan, V.T.; Van, S.L.; Seiler, C.; Junker, A.; Weigelt-Fischer, K.; Klukas, C.; Altmann, T.; Sreenivasulu, N.; Bäumlein, H.; Kuhlmann, M. AtRD22 and AtUSPL1, members of the plant-specific BURP domain family involved in Arabidopsis thaliana drought tolerance. PLoS ONE 2014, 9, e110065. [Google Scholar] [CrossRef] [PubMed]
- Chien, P.S.; Nam, H.G.; Chen, Y.R. A salt-regulated peptide derived from the CAP superfamily protein negatively regulates salt-stress tolerance in Arabidopsis. J. Exp. Bot. 2015, 66, 5301–5313. [Google Scholar] [CrossRef] [PubMed]
- Paulose, B.; Kandasamy, S.; Dhankher, O.P. Expression profiling of Crambe abyssinica under arsenate stress identifies genes and gene networks involved in arsenic metabolism and detoxification. BMC Plant Biol. 2010, 10, 108. [Google Scholar] [CrossRef] [PubMed]
- Lan, P.; Li, W.; Wen, T.N.; Shiau, J.Y.; Wu, Y.C.; Lin, W.D.; Schmidt, W.S. iTRAQ protein profile analysis of Arabidopsis roots reveals new aspects critical for iron homeostasis. Plant Physiol. 2011, 155, 821–834. [Google Scholar] [CrossRef]
- Van, L.C.; Rep, M.; Pieterse, C.M. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar]
- Pecenkova, T.; Pleskot, R.; Zarsky, V. Subcellular localization of Arabidopsis pathogenesis-related 1 (PR1) protein. Int. J. Mol. Sci. 2017, 18, 825. [Google Scholar] [CrossRef]
- Breen, S.; Williams, S.J.; Outram, M.; Kobe, B.; Solomon, P.S. Emerging insights into the functions of pathogenesis-related protein 1. Trends Plant Sci. 2017, 22, 871–879. [Google Scholar] [CrossRef]
- Choudhary, V.; Schneiter, R. Pathogen-Related Yeast (PRY) proteins and members of the CAP superfamily are secreted sterol-binding proteins. Proc. Natl. Acad. Sci. USA 2012, 109, 16882–16887. [Google Scholar] [CrossRef]
- Ji, R.; Gao, S.; Bi, Q.; Wang, Y.; Lv, M.; Ge, W.; Feng, H. The Salicylic acid signaling pathway plays an important role in the resistant process of Brassica rapa L. ssp. pekinensis to Plasmodiophora brassicae Woronin. J. Plant Growth Regul. 2020, 40, 405–422. [Google Scholar] [CrossRef]
- Zhang, J.; Ge, W.; Chang, H.; Xin, X.; Ji, R. Discovery of BrATG6 and its potential role in Brassica rapa L. resistance to infection by Plasmodiophora brassicae. Gene 2021, 791, 145711. [Google Scholar] [CrossRef]
- Wangorsch, A.; Scheurer, S.; Blanca, M.; Blanca-Lopez, N.; Somoza, M.L.; Martín-Pedraza, L. Allergenic Properties and Molecular Characteristics of PR-1 Proteins. Front. Allergy 2022, 3, 824717. [Google Scholar] [CrossRef]
- Almeida-Silva, F.; Venancio, T.M. Pathogenesis-related protein 1 (PR-1) genes in soybean: Genome-wide identification, structural analysis and expression profiling under multiple biotic and abiotic stresses. Gene 2022, 809, 146013. [Google Scholar] [CrossRef]
- Akbudak, M.A.; Yildiz, S.; Filiz, E. Pathogenesis related protein-1 (PR-1) genes in tomato (Solanum lycopersicum L.): Bioinformatics analyses and expression profiles in response to drought stress. Genomics 2020, 112, 4089–4099. [Google Scholar] [CrossRef]
- Zhang, Q.; Guo, N.; Zhang, Y.; Yu, Y.; Liu, S. Genome-Wide characterization and expression analysis of pathogenesis-related 1 (PR-1) gene family in tea plant (Camellia sinensis (L.) O. Kuntze) in response to blister-blight disease stress. Int. J. Mol. Sci. 2022, 23, 1292. [Google Scholar] [CrossRef]
- Ma, L.; Meng, Q.; Shi, F.; Liu, J.; Li, Y.; Liu, C.; Liu, X.; Su, B.; He, C.; Ji, C. Genome-wide analysis of maize PR-1 gene family and expression profiles induced by plant hormones and fungal phytopathogens. Am. J. Transl. 2022, 14, 8315–8331. [Google Scholar]
- Liu, J.; Wang, H.; Huang, C.; Qian, H. Subcellular localization of MTA proteins in normal and cancer cells. Cancer Metastasis Rev. 2014, 33, 843–856. [Google Scholar] [CrossRef]
- Yang, Y.; LI, R.; QI, M. In vivo analysis of plant promoters and transcription factors by agroinfiltration of tobacco leaves. Plant J. 2000, 22, 543–551. [Google Scholar] [CrossRef]
- Christou, P. Strategies for variety-independent genetic transformation of important cereals, legumes and woody species utilizing particle bombardment. Euphytica 1995, 85, 13–27. [Google Scholar] [CrossRef]
- Sheen, J. Signal transduction in maize and Arabidopsis mesophyll protoplasts. Plant Physiol. 2001, 127, 1466–1475. [Google Scholar] [CrossRef]
- Gu, Y.; Innes, R.W. The keep on going protein of Arabidopsis regulates intracellular protein trafficking and is degraded during fungal infection. Plant Cell 2012, 24, 4717–4730. [Google Scholar] [CrossRef]
- Watanabe, S.; Shimada, T.L.; Hiruma, K.; Takano, Y. Pathogen infection trial increases the secretion of proteins localized in the endoplasmic reticulum body of Arabidopsis. Plant Physiol. 2013, 163, 659–664. [Google Scholar] [CrossRef]
- Zhang, J.R.; Sun, X.; Liang, F.; Wang, F.; Zhang, Y.J.; Wang, H.Y.; Liu, D.Q. Identification of the signal peptide and subcellular localization of pathogenesis-related protein1 gene from wheat. J Hebei Agric Univ. 2017, 40, 1–7. [Google Scholar]
- Wu, Y.; Zhang, D.; Chu, J.Y.; Boyle, P.; Wang, Y.; Brindle, I.D.; De Luca, V.; Despres, C. The Arabidopsis NPR1 protein is a receptor for the plant defense hormone salicylic acid. Cell Rep. 2012, 1, 639–647. [Google Scholar] [CrossRef]
- Matthews, B.F.; Beard, H.; Brewer, E.; Kabir, S.; MacDonald, M.H.; Youssef, R.M. Arabidopsis genes, AtNPR1, AtTGA2 and AtPR-5, confer partial resistance to soybean cystnematode (Heterodera glycines) when over expressed in transgenic soybean roots. BMC Plant Biol. 2014, 14, 96. [Google Scholar] [CrossRef]
- Gamir, J.; Darwiche, R.; Van’t Hof, P.; Choudhary, V.; Stumpe, M.; Schneiter, R.; Mauch, F. The sterol-binding activity of pathogenesis-related protein 1 reveals the mode of action of an antimicrobial protein. Plant J. 2017, 89, 502–509. [Google Scholar] [CrossRef]
- Bai, L.; Ma, X.; Zhang, G.; Song, S.; Zhou, Y.; Gao, L.; Miao, Y.; Song, C.P. A receptor-like kinase mediates ammonium homeostasis and is important for the polar growth of root hairs in Arabidopsis. Plant Cell 2014, 26, 1497–1511. [Google Scholar] [CrossRef]
- You, Q.; Dong, N.; Yang, H.; Feng, F.; Xu, Y.; Wang, C.; Yang, Y.; Ma, X.; Bai, L. The Arabidopsis receptor-like kinase CAP1 promotes shoot growth under ammonium stress. Biology 2022, 11, 1452. [Google Scholar] [CrossRef]
- Yu, J.; Ma, X.; Wang, L.; Dong, N.; Wang, K.; You, Q.; Xu, Y.; Wang, C.; Dong, Z.; Shi, Z.; et al. Arabidopsis CAP1 mediates ammonium-regulated root hair growth by influencing vesicle trafficking and the cytoskeletal arrangement in root hair cells. J. Genet. Genom. 2022, 49, 986–989. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Comparison Result | Percent Identity |
|---|---|
| Brassica rapa (XM_009121709.2) | 100% |
| Brassica napus (XM_013810782.3) | 99% |
| Brassica oleracea var (XM_013730593.1) | 98% |
| Arabidopsis thaliana CAP (NM_119530.3) | 94% |
| Expression Level of BrCAP | p | |
|---|---|---|
| CK | 1 ± 0.13 | 0.000 |
| T | 11.73 ± 0.18 * |
| Treatment | Expression Level of BrCAP | Statistical Data | |||
|---|---|---|---|---|---|
| Root | Stem | Leaf | F | p | |
| CK | 1 ± 0.13 b | 0.01 ± 0.0006 c | 0.01 ± 0.0013 c | 262.917 | 0.000 |
| T | 11.73 ± 0.18 a | 0.01 ± 0.0003 c | 0.01 ± 0 c | ||
| Period after Inoculation | Expression Level of BrCAP | Statistical Data | ||
|---|---|---|---|---|
| CK | T | F | p | |
| 14 days | 1 ± 0 | 9.2 ± 0.07 | 2.629 | 0.00 |
| 40 days | 1 ± 0.06 * | 20 ± 1.33 * | 0.130 | 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, J.; Gao, S.; Zhang, B.; Ge, W.; Zhang, J.; Ji, R. Chinese Cabbage BrCAP Has Potential Resistance against Plasmodiophora brassicae. Horticulturae 2023, 9, 517. https://doi.org/10.3390/horticulturae9040517
Zou J, Gao S, Zhang B, Ge W, Zhang J, Ji R. Chinese Cabbage BrCAP Has Potential Resistance against Plasmodiophora brassicae. Horticulturae. 2023; 9(4):517. https://doi.org/10.3390/horticulturae9040517
Chicago/Turabian StyleZou, Jiawei, Shiqi Gao, Bo Zhang, Wenjie Ge, Jing Zhang, and Ruiqin Ji. 2023. "Chinese Cabbage BrCAP Has Potential Resistance against Plasmodiophora brassicae" Horticulturae 9, no. 4: 517. https://doi.org/10.3390/horticulturae9040517
APA StyleZou, J., Gao, S., Zhang, B., Ge, W., Zhang, J., & Ji, R. (2023). Chinese Cabbage BrCAP Has Potential Resistance against Plasmodiophora brassicae. Horticulturae, 9(4), 517. https://doi.org/10.3390/horticulturae9040517