
Fruit Cuticle Composition in ‘Arbequina’ Olive: Time–Course Changes along On-Tree Ripening under Irrigated and Rain-Fed Conditions



Abstract
“Good morrow, fair ones; pray you, if you know,Where in the purlieus of this forest standsA sheep-cote fenc’d about with olive trees?”William ShakespeareAs You Like It (Act IV, Scene III)
1. Introduction
2. Materials and Methods
2.1. Plant Material and Toluidine Blue (TB) Test
2.2. Cuticle Isolation
2.3. Extraction and Analysis of Cuticular Wax
2.4. Extraction and Analysis of Cutin Monomers
2.5. Assessment of Cuticle Thickness
2.6. Statistical Analysis
3. Results and Discussion
3.1. The Impact of Irrigation on Physical Characteristics of Fruits
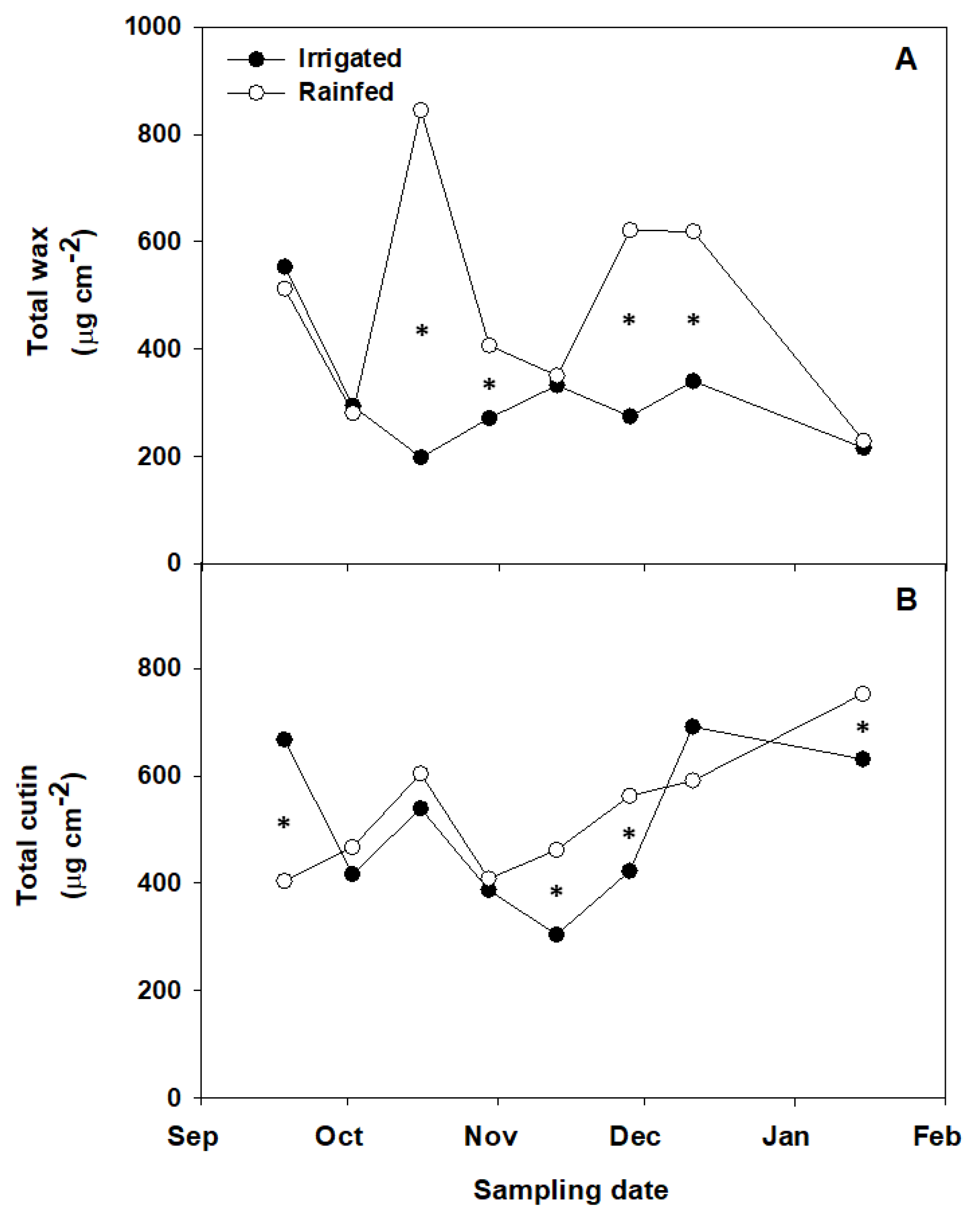
3.2. The Impact of Irrigation on Fruit Cuticle Characteristics
3.3. Wax Composition of ‘Arbequina’ Olive Fruit Cuticles
3.4. Cutin Monomer Composition of ‘Arbequina’ Olive Fruit Cuticles
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ninot, A.; Hermoso, J.F.; Martí, E.; Rovira, M.; Batlle, I.; Romero, A. Recuperació i Conservació de Varietat Autòctones d’Olivera. Dossier Tècnic; Departament d’Agricultura, Ramaderia i Pesca, Generalitat de Catalunya: Catalonia, Spain, 2015; Volume 80, pp. 6–16. ISSN 1699-5465. [Google Scholar]
- Tous, J. The influence of growing region and cultivar on olives and olive oil characteristics and on their functional constituents. In Olives and Olive Oil as Functional foods: Bioactivity, Chemistry and Processing; Kiritsakis, A., Shahidi, F., Eds.; John Wiley & Sons Ltd.: Oxford, UK, 2017; pp. 45–80. [Google Scholar] [CrossRef]
- Ninot, A.; Howad, W.; Romero, A. Les Varietats Catalanes d’Olivera; Quaderns Agraris (Institució Catalana d’Estudis Agraris): Barcelona, Spain, 2019; Volume 46, pp. 7–36. [Google Scholar] [CrossRef]
- Riederer, M.; Schreiber, L. Protecting against water loss: Analysis of the barrier properties of plant cuticles. J. Exp. Bot. 2001, 52, 2023–2032. [Google Scholar] [CrossRef] [PubMed]
- Buschhaus, C.; Jetter, R. Composition differences between epicuticular and intracuticular wax substructures: How do plants seal their epidermal surfaces? J. Exp. Bot. 2011, 62, 841–853. [Google Scholar] [CrossRef]
- Domínguez, E.; Heredia-Guerrero, J.A.; Heredia, A. The biophysical design of plant cuticles: An overview. New Phytol. 2011, 189, 938–9499. [Google Scholar] [CrossRef]
- Trivedi, P.; Nguyen, N.; Hykkerud, A.L.; Häggman, H.; Martinussen, I.; Jaakola, L.; Karppinen, K. Developmental and Environmental Regulation of Cuticular Wax Biosynthesis in Fleshy Fruits. Front. Plant Sci. 2019, 10, 431. [Google Scholar] [CrossRef] [PubMed]
- Lara, I.; Heredia, A.; Domínguez, E. Shelf Life Potential and the Fruit Cuticle: The Unexpected Player. Front. Plant Sci. 2019, 10, 770. [Google Scholar] [CrossRef]
- Lara, I.; Belge, B.; Goulao, L.F. The fruit cuticle as a modulator of postharvest quality. Postharvest Biol. Technol. 2014, 87, 103–112. [Google Scholar] [CrossRef]
- Riederer, M.; Arand, K.; Burghardt, M.; Huang, H.; Riedel, M.; Schuster, A.-C.; Smirnova, A.; Jiang, Y. Water loss from litchi (Litchi chinensis) and longan (Dimocarpus longan) fruits is biphasic and controlled by a complex pericarpal transpiration barrier. Planta 2015, 242, 1207–1219. [Google Scholar] [CrossRef]
- Bianchi, G. Lipids and phenols in table olives. Eur. J. Lipid Sci. Technol. 2003, 105, 229–242. [Google Scholar] [CrossRef]
- Lanza, B.; Di Serio, M.G. SEM characterization of olive (Olea europaea L.) fruit epicuticular waxes and epicarp. Sci. Hortic. 2015, 191, 49–56. [Google Scholar] [CrossRef]
- Vichi, S.; Cortés-Francisco, N.; Caixach, J.; Barrios, G.; Mateu, J.; Ninot, A.; Romero, A. Epicuticular Wax in Developing Olives (Olea europaea) Is Highly Dependent upon Cultivar and Fruit Ripeness. J. Agric. Food Chem. 2016, 64, 5985–5994. [Google Scholar] [CrossRef]
- Huang, H.; Burghardt, M.; Schuster, A.-C.; Leide, J.; Lara, I.; Riederer, M. Chemical Composition and Water Permeability of Fruit and Leaf Cuticles of Olea europaea L. J. Agric. Food Chem. 2017, 65, 8790–8797. [Google Scholar] [CrossRef]
- Diarte, C.; Lai, P.-H.; Huang, H.; Romero, A.; Casero, T.; Gatius, F.; Graell, J.; Medina, V.; East, A.; Riederer, M.; et al. Insights into Olive Fruit Surface Functions: A Comparison of Cuticular Composition, Water Permeability, and Surface Topography in Nine Cultivars During Maturation. Front. Plant Sci. 2019, 10, 1484. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Tanaka, H.; Machida, C.; Watanabe, M.; Machida, Y. A new method for rapid visualization of defects in leaf cuticle reveals five intrinsic patterns of surface defects in Arabidopsis. Plant J. 2004, 37, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Inglese, P.; Famiani, F.; Galvano, F.; Servili, M.; Esposto, S.; Urbani, S. Factors affecting extra-virgin olive oil composition. Hortic. Rev. 2011, 38, 83–147. [Google Scholar] [CrossRef]
- Diarte, C.; Iglesias, A.; Graell, J.; Lara, I. Firmness and cell wall changes during maturation of ‘Arbequina’ olive fruit: The impact of irrigation. Horticulturae 2022, 8, 872. [Google Scholar] [CrossRef]
- Berenguer, M.J.; Vossen, P.M.; Grattan, S.R.; Connell, J.H.; Polito, V.S. Tree Irrigation Levels for Optimum Chemical and Sensory Properties of Olive Oil. Hortscience 2006, 41, 427–432. [Google Scholar] [CrossRef]
- Gómez-Rico, A.; Salvador, M.D.; Moriana, A.; Pérez, D.; Olmedilla, N.; Ribas, F.; Fregapane, G. Influence of different irrigation strategies in a traditional Cornicabra cv. olive orchard on virgin olive oil composition and quality. Food Chem. 2007, 100, 568–578. [Google Scholar] [CrossRef]
- Morales-Sillero, A.; García, J.; Torres-Ruiz, J.; Montero, A.; Sánchez-Ortiz, A.; Fernández, J. Is the productive performance of olive trees under localized irrigation affected by leaving some roots in drying soil? Agric. Water Manag. 2013, 123, 79–92. [Google Scholar] [CrossRef]
- Mechri, B.; Tekaya, M.; Hammami, M.; Chehab, H. Effects of drought stress on phenolic accumulation in greenhouse-grown olive trees (Olea europaea). Biochem. Syst. Ecol. 2020, 92, 104112. [Google Scholar] [CrossRef]
- Fernández, V.; Eichert, T. Uptake of Hydrophilic Solutes Through Plant Leaves: Current State of Knowledge and Perspectives of Foliar Fertilization. Crit. Rev. Plant Sci. 2009, 28, 36–68. [Google Scholar] [CrossRef]
- Fernández, V.; Sotiropoulos, T.; Brown, P.H. Foliar Fertilization: Scientific Principles and Field Practices; International Fertilizer Industry Association: Paris, France, 2013; ISBN 979-10-92366-00-6. [Google Scholar]
- Patwari, P.; Salewski, V.; Gutbrod, K.; Kreszies, T.; Dresen-Scholz, B.; Peisker, H.; Steiner, U.; Meyer, A.J.; Schreiber, L.; Dörmann, P. Surface wax esters contribute to drought tolerance in Arabidopsis. Plant J. 2019, 98, 727–744. [Google Scholar] [CrossRef] [PubMed]
- Torres, R.; Ferrara, G.; Soto, F.; López, J.A.; Sánchez, F.; Mazzeo, A.; Pastor, A.P.; Domingo, R. Effects of soil and climate in a table grape vineyard with cover crops. Irrigation management using sensors networks. Cienc. Tec. Vitivinic. 2017, 32, 72–81. [Google Scholar] [CrossRef]
- Kosma, D.K.; Bourdenx, B.; Bernard, A.; Parsons, E.P.; Lu, S.; Joubes, J.; Jenks, M.A. The impacts of water deficiency on leaf cuticle lipids of Arabidopsis. Plant Physiol. 2003, 151, 1918–1929. [Google Scholar] [CrossRef] [PubMed]
- Zarrouk, O.; Pinheiro, C.; Misra, C.S.; Fernández, V.; Chaves, M.M. Fleshy fruit epidermis is a protective barrier under water stress. In Water Scarcity and Sustainable Agriculture in Semiarid Environment. Tools, Strategies, and Challenges for Woody Crops; García-Tejero, I.F., Zuazo, V.H.D., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 507–533. [Google Scholar] [CrossRef]
- Bianchi, G.; Murelli, C.; Vlahov, G. Surface waxes from olive fruits. Phytochemistry 1992, 31, 3503–3506. [Google Scholar] [CrossRef]
- Guinda, A.; Rada, M.; Delgado, T.; Gutiérrez-Adánez, P.; Castellano, J.M. Pentacyclic Triterpenoids from Olive Fruit and Leaf. J. Agric. Food Chem. 2010, 58, 9685–9691. [Google Scholar] [CrossRef]
- Jiménez-Herrera, R.; Pacheco-López, B.; Peragón, J. Water Stress, Irrigation and Concentrations of Pentacyclic Triterpenes and Phenols in Olea europaea L. cv. Picual Olive Trees. Antioxidants 2019, 8, 294. [Google Scholar] [CrossRef]
- Grncarevic, M.; Radler, F. The effect of wax components on cuticular transpiration-model experiments. Planta 1967, 75, 23–27. [Google Scholar] [CrossRef]
- Leide, J.; Hildebrandt, U.; Reussing, K.; Riederer, M.; Vogg, G. The Developmental Pattern of Tomato Fruit Wax Accumulation and Its Impact on Cuticular Transpiration Barrier Properties: Effects of a Deficiency in a β-Ketoacyl-Coenzyme A Synthase (LeCER6). Plant Physiol. 2007, 144, 1667–1679. [Google Scholar] [CrossRef]
- Leide, J.; Hildebrandt, U.; Vogg, G.; Riederer, M. The positional sterile (ps) mutation affects cuticular transpiration and wax biosynthesis of tomato fruits. J. Plant Physiol. 2011, 168, 871–877. [Google Scholar] [CrossRef]
- Belge, B.; Llovera, M.; Comabella, E.; Gatius, F.; Guillén, P.; Graell, J.; Lara, I. Characterization of Cuticle Composition after Cold Storage of “Celeste” and “Somerset” Sweet Cherry Fruit. J. Agric. Food Chem. 2014, 62, 8722–8729. [Google Scholar] [CrossRef] [PubMed]
- Belge, B.; Goulao, L.F.; Comabella, E.; Graell, J.; Lara, I. Postharvest heat and CO2 shocks induce changes in cuticle composition and cuticle-related gene expression in ‘October Sun’ peach fruit. Postharvest Biol. Technol. 2019, 148, 200–207. [Google Scholar] [CrossRef]
- Kallio, H.; Nieminen, R.; Tuomasjukka, S.; Hakala, M. Cutin Composition of Five Finnish Berries. J. Agric. Food Chem. 2006, 54, 457–462. [Google Scholar] [CrossRef]
- Järvinen, R.; Kaimainen, M.; Kallio, H. Cutin composition of selected northern berries and seeds. Food Chem. 2010, 122, 137–144. [Google Scholar] [CrossRef]
- Peschel, S.; Franke, R.; Schreiber, L.; Knoche, M. Composition of the cuticle of developing sweet cherry fruit. Phytochemistry 2007, 68, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Bonaventure, G.; Beisson, F.; Ohlrogge, J.; Pollard, M. Analysis of the aliphatic monomer composition of polyesters associated with Arabidopsis epidermis: Occurrence of octadeca-cis-6, cis-9-diene-1,18-dioate as the major component. Plant J. 2004, 40, 920–930. [Google Scholar] [CrossRef] [PubMed]
- Bedbabis, S.; Ben Rouina, B.; Boukhris, M.; Ferrara, G. Effects of Irrigation with Treated Wastewater on Root and Fruit Mineral Elements of Chemlali Olive Cultivar. Sci. World J. 2014, 2014, 973638. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rainfall (mm) | 20.2 | 9.6 | 77.8 | 14.8 | 34.0 | 47.0 | 13.0 | 2.9 | 48.0 | 33.0 | 9.6 | 8.1 |
| Absolute Max T (°C) | 15.4 | 16.6 | 25.8 | 27.8 | 33.5 | 37.0 | 37.5 | 38.7 | 30.2 | 28.4 | 22.1 | 17.7 |
| Average Max T (°C) | 8.0 | 13.5 | 18.1 | 20.8 | 25.8 | 31.2 | 32.4 | 32.1 | 25.4 | 23.5 | 14.8 | 9.5 |
| Average T (°C) | 4.2 | 8.6 | 11.8 | 13.9 | 18.7 | 24.1 | 25.4 | 25.3 | 19.2 | 17.3 | 9.3 | 5.1 |
| Average Min T (°C) | 0.4 | 3.7 | 5.5 | 7.1 | 11.5 | 17.0 | 18.4 | 18.6 | 12.9 | 11.1 | 3.7 | 0.7 |
| Absolute Min T (°C) | −5.3 | 0.2 | 0.2 | 2.2 | 5.0 | 11.9 | 12.0 | 10.8 | 7.2 | 6.5 | −1.8 | −4.2 |
| Picking | Date | Irrigation Regime | Weight (g) | Length (mm) | Diameter (mm) | F:S Ratio | Water Content (%) | TB Test | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 18 Sep | Irrigated | 1.23 | e A | 14.18 | d A | 12.66 | f A | 3.80 | c A | 67.02 | b A | - |
| 2 | 3 Oct | 1.62 | cd A | 15.51 | bc A | 14.00 | cde A | 4.57 | b A | 69.15 | a A | + | |
| 3 | 16 Oct | 1.81 | bc A | 16.01 | a A | 14.77 | a A | 5.55 | a A | 66.67 | b A | + | |
| 4 | 30 Oct | 1.86 | ab A | 15.60 | abc A | 14.08 | cd A | 5.58 | a A | 66.44 | b A | + | |
| 5 | 13 Nov | 2.02 | a A | 15.90 | ab A | 14.52 | ab A | 4.69 | b A | 60.71 | c A | + | |
| 6 | 28 Nov | 1.84 | ab A | 15.48 | bc A | 14.21 | bc A | 4.46 | b A | 54.99 | d A | ne | |
| 7 | 11 Dec | 1.58 | d A | 15.20 | c A | 13.68 | de A | 4.55 | b A | 44.92 | e A | ne | |
| 8 | 15 Jan | 1.59 | d A | 15.42 | c A | 13.61 | e A | 3.61 | c A | 41.92 | f A | ne | |
| 1 | 18 Sep | Rain fed | 1.08 | d B | 12.90 | c B | 11.63 | e B | 3.19 | e B | 60.18 | ab A | - |
| 2 | 3 Oct | 1.38 | b A | 13.54 | b B | 12.12 | cd B | 3.65 | cd B | 61.85 | a B | + | |
| 3 | 16 Oct | 1.23 | c B | 13.32 | bc B | 11.86 | de B | 3.63 | cd B | 48.94 | c B | - | |
| 4 | 30 Oct | 1.42 | b B | 13.73 | b B | 12.53 | bc B | 4.48 | a B | 57.78 | b B | - | |
| 5 | 13 Nov | 1.52 | a B | 14.59 | a B | 12.96 | ab B | 3.86 | bc A | 47.33 | c B | - | |
| 6 | 28 Nov | 1.28 | c B | 14.49 | a B | 12.69 | ab B | 3.41 | de B | 43.92 | d B | ne | |
| 7 | 11 Dec | 1.57 | a A | 14.40 | a B | 12.59 | abc B | 4.01 | b B | 39.28 | e B | ne | |
| 8 | 15 Jan | 1.54 | a A | 14.32 | a B | 13.04 | a B | 3.45 | de A | 39.44 | e B | ne | |
| Picking | Date | Irrigation Regime | Cuticle Yield (mg cm−2) | Wax/Cutin Ratio | Thickness (μm) | Wax (%) | Cutin (%) | C16/C18 * | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 18 Sep | Irrigated | 2.8 | a A | 0.83 | b B | 36.0 | b B | 20.0 | a A | 24.1 | bc A | 0.81 |
| 2 | 3 Oct | 2.1 | cd B | 0.71 | bc A | 43.6 | a A | 13.9 | b A | 19.6 | de A | 0.55 | |
| 3 | 16 Oct | 2.2 | cd B | 0.37 | de B | 39.8 | ab A | 9.2 | c B | 25.0 | bc A | 0.49 | |
| 4 | 30 Oct | 2.1 | cd A | 0.70 | bc B | 40.2 | ab A | 13.2 | b B | 18.8 | de A | 0.45 | |
| 5 | 13 Nov | 1.7 | e A | 0.99 | a A | 38.6 | ab A | 20.0 | a A | 18.3 | e A | 0.49 | |
| 6 | 28 Nov | 2.0 | d A | 0.65 | c B | 41.3 | ab A | 16.1 | b B | 21.5 | cd B | 0.62 | |
| 7 | 11 Dec | 2.3 | bc A | 0.49 | d B | 41.4 | ab A | 14.9 | b B | 30.2 | a A | 0.51 | |
| 8 | 15 Jan | 2.5 | ab A | 0.34 | e A | 44.9 | a A | 8.5 | c A | 25.0 | b A | 0.43 | |
| 1 | 18 Sep | Rain fed | 2.4 | c A | 1.38 | a A | 53.7 | a A | 21.8 | b A | 17.2 | c A | 0.66 |
| 2 | 3 Oct | 2.7 | b A | 0.60 | d A | 46.5 | abc A | 10.4 | d B | 17.4 | c A | 0.55 | |
| 3 | 16 Oct | 3.8 | a A | 0.79 | cd A | 49.3 | ab A | 22.1 | b A | 15.8 | c B | 0.45 | |
| 4 | 30 Oct | 2.2 | c A | 1.00 | bc A | 42.8 | bcd A | 18.1 | bc A | 18.2 | c A | 0.48 | |
| 5 | 13 Nov | 2.2 | c A | 0.70 | cd A | 36.5 | d A | 16.1 | c B | 21.2 | c A | 0.54 | |
| 6 | 28 Nov | 1.9 | d A | 1.10 | ab A | 38.8 | cd A | 33.4 | a A | 30.2 | ab A | 0.56 | |
| 7 | 11 Dec | 1.7 | d B | 1.05 | b A | 40.6 | cd A | 37.1 | a A | 35.5 | a A | 0.70 | |
| 8 | 15 Jan | 2.6 | b A | 0.30 | e B | 38.5 | cd A | 8.9 | d A | 29.3 | b A | 0.50 | |
| Picking | Date | Irrigation Regime | ACL | Acyclic/Cyclic Ratio | Triterpenoids (%) | Fatty Acids (%) | Alcohols (%) | n-Alkanes (%) | Sterols (%) | Unidentified (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 18 Sep | Irrigated | 24.3 | abc A | 0.17 | c A | 71.7 | a A | 7.4 | c A | 3.70 | d A | 1.0 | cd A | 0.9 | ab A | 15.3 | abc A |
| 2 | 3 Oct | 24.0 | e B | 0.20 | bc A | 69.0 | ab A | 9.6 | abc A | 3.60 | d B | 1.0 | cd B | 0.8 | b A | 16.1 | ab A | |
| 3 | 16 Oct | 24.0 | de A | 0.24 | abc A | 65.2 | b A | 10.2 | ab A | 3.94 | d A | 1.8 | a A | 1.1 | a A | 17.7 | a A | |
| 4 | 30 Oct | 24.3 | ab A | 0.28 | a A | 65.8 | ab A | 11.5 | a A | 6.13 | ab A | 1.1 | c A | 0.8 | ab A | 14.6 | bcd A | |
| 5 | 13 Nov | 24.1 | cde A | 0.26 | ab A | 66.6 | ab A | 11.2 | a A | 5.41 | bc A | 1.0 | cd A | 0.8 | b A | 15.0 | abc A | |
| 6 | 28 Nov | 24.3 | ab A | 0.24 | abc A | 70.4 | ab A | 8.7 | bc A | 7.38 | a A | 1.1 | cd A | 0.6 | b A | 11.8 | d A | |
| 7 | 11 Dec | 24.4 | a A | 0.26 | ab A | 67.8 | ab A | 10.2 | ab A | 5.77 | bc A | 1.4 | b A | 0.6 | b A | 14.1 | bcd A | |
| 8 | 15 Jan | 24.2 | bcd B | 0.23 | abc A | 70.4 | ab A | 10.6 | ab A | 4.51 | cd B | 0.8 | d A | 0.8 | b A | 12.8 | cd A | |
| 1 | 18 Sep | Rain fed | 24.4 | b A | 0.17 | cd A | 69.8 | b A | 7.1 | c A | 3.81 | de A | 1.4 | a A | 0.9 | a A | 17.0 | ab A |
| 2 | 3 Oct | 24.8 | a A | 0.29 | a A | 62.4 | c A | 10.1 | a A | 6.61 | a A | 1.3 | a A | 0.7 | ab A | 18.8 | a A | |
| 3 | 16 Oct | 24.2 | bc A | 0.17 | cd A | 72.0 | ab A | 8.1 | bc A | 4.14 | cde A | 0.4 | b B | 0.6 | b A | 14.7 | bc A | |
| 4 | 30 Oct | 23.9 | d B | 0.21 | bc A | 70.0 | b A | 9.7 | ab A | 5.04 | bcd A | 0.4 | b B | 0.9 | a A | 13.9 | bc A | |
| 5 | 13 Nov | 23.3 | e B | 0.20 | bcd A | 70.6 | b A | 9.8 | a A | 3.66 | e B | 0.5 | b B | 0.8 | ab A | 14.7 | bc A | |
| 6 | 28 Nov | 24.1 | bcd A | 0.15 | d B | 76.9 | a A | 6.6 | c B | 5.08 | bc A | 0.3 | b B | 0.9 | ab A | 10.2 | d A | |
| 7 | 11 Dec | 24.0 | cd A | 0.18 | cd A | 72.6 | ab A | 7.9 | c B | 4.26 | cde A | 0.7 | b B | 0.8 | ab A | 13.7 | bc A | |
| 8 | 15 Jan | 24.3 | b A | 0.24 | ab A | 69.1 | b A | 10.0 | a A | 6.15 | ab A | 0.6 | b A | 0.6 | b A | 13.6 | c A | |
| Picking | Date | Irrigation Regime | FA (%) | α,ω-diFA (%) | α,ω-diFA, mcOH (%) | ω-OH FA (%) | ω-OH FA, mcOH (%) | α-OH FA (%) | Alcohols (%) | Unidentified (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 15 Sep | Irrigated | 21.35 | ab A | 4.39 | d A | 1.97 | a A | 23.48 | c A | 19.39 | a A | 1.25 | ab A | 1.82 | cd A | 26.34 | a A |
| 2 | 4 Oct | 21.88 | ab A | 9.34 | bc A | 1.14 | bc B | 27.76 | ab A | 12.48 | bc A | 0.63 | b A | 1.64 | d A | 25.14 | ab A | |
| 3 | 17 Oct | 27.32 | a A | 8.52 | c B | 1.17 | bc B | 26.47 | bc A | 12.13 | bc A | 0.82 | ab A | 1.70 | d A | 21.86 | d B | |
| 4 | 31 Oct | 23.47 | ab A | 10.73 | ab A | 0.99 | c B | 28.63 | ab A | 10.28 | c B | 0.80 | ab B | 1.91 | bcd B | 23.18 | d B | |
| 5 | 14 Nov | 17.58 | bc A | 10.97 | a A | 1.01 | c B | 29.52 | a A | 11.84 | c A | 0.88 | ab B | 2.05 | abc B | 26.17 | a A | |
| 6 | 29 Nov | 13.42 | c A | 11.15 | a A | 1.93 | a A | 28.00 | ab A | 18.18 | a A | 1.73 | a A | 2.23 | ab A | 23.36 | cd A | |
| 7 | 12 Dec | 13.45 | c A | 12.04 | a A | 1.78 | ab A | 29.37 | a A | 14.97 | b B | 1.23 | ab A | 2.18 | abc A | 24.98 | abc A | |
| 8 | 16 Jan | 19.57 | b A | 12.21 | a A | 1.20 | bc A | 28.35 | ab A | 11.60 | c A | 1.19 | ab A | 2.39 | a A | 23.48 | bcd B | |
| 1 | 15 Sep | Rain fed | 13.45 | bc A | 7.37 | d A | 2.00 | a A | 27.06 | a A | 16.97 | ab A | 1.48 | ab A | 2.08 | ab A | 29.58 | a A |
| 2 | 4 Oct | 16.42 | ab A | 10.44 | abc A | 1.68 | a A | 28.26 | a A | 14.21 | bc A | 0.75 | c A | 1.72 | c A | 26.52 | bc A | |
| 3 | 17 Oct | 19.95 | a B | 10.67 | abc A | 1.66 | a A | 27.50 | a A | 10.75 | c B | 0.88 | bc A | 1.83 | bc A | 26.76 | bc A | |
| 4 | 31 Oct | 16.26 | b B | 9.84 | bc A | 2.05 | aA | 26.65 | a A | 13.69 | bc A | 1.39 | ab A | 2.15 | a A | 27.97 | ab A | |
| 5 | 14 Nov | 16.62 | ab A | 9.46 | bc B | 1.51 | a A | 27.82 | a A | 15.33 | bc A | 1.66 | a A | 2.20 | a A | 25.41 | c A | |
| 6 | 29 Nov | 11.77 | c A | 11.67 | a A | 1.68 | a A | 29.74 | a A | 14.81 | bc A | 1.12 | abc A | 2.17 | a A | 27.03 | bc A | |
| 7 | 12 Dec | 13.30 | bc A | 9.12 | cd B | 1.90 | a A | 25.88 | a B | 20.30 | a A | 1.27 | abc A | 2.16 | a A | 26.08 | bc A | |
| 8 | 16 Jan | 15.68 | b A | 11.00 | ab A | 1.42 | a A | 26.84 | a A | 14.89 | bc A | 1.26 | abc A | 2.30 | a A | 26.62 | bc A | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diarte, C.; Iglesias, A.; Graell, J.; Lara, I. Fruit Cuticle Composition in ‘Arbequina’ Olive: Time–Course Changes along On-Tree Ripening under Irrigated and Rain-Fed Conditions. Horticulturae 2023, 9, 394. https://doi.org/10.3390/horticulturae9030394
Diarte C, Iglesias A, Graell J, Lara I. Fruit Cuticle Composition in ‘Arbequina’ Olive: Time–Course Changes along On-Tree Ripening under Irrigated and Rain-Fed Conditions. Horticulturae. 2023; 9(3):394. https://doi.org/10.3390/horticulturae9030394
Chicago/Turabian StyleDiarte, Clara, Anna Iglesias, Jordi Graell, and Isabel Lara. 2023. "Fruit Cuticle Composition in ‘Arbequina’ Olive: Time–Course Changes along On-Tree Ripening under Irrigated and Rain-Fed Conditions" Horticulturae 9, no. 3: 394. https://doi.org/10.3390/horticulturae9030394
APA StyleDiarte, C., Iglesias, A., Graell, J., & Lara, I. (2023). Fruit Cuticle Composition in ‘Arbequina’ Olive: Time–Course Changes along On-Tree Ripening under Irrigated and Rain-Fed Conditions. Horticulturae, 9(3), 394. https://doi.org/10.3390/horticulturae9030394