

Studies on the Identification of Resistance to Fusarium oxysporum (Schlecht.) in Different Genetic Backgrounds of Asparagus officinalis (L.) and Its Defense Responses

Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Isolates and Cultures
2.2. Plant Material and Growing Conditions
2.3. Seedling Resistance Test
2.4. Staining Procedure and Microscopic Determination of Hypersensitive Response
2.5. SSR Markers and Distance Analysis
2.6. Statistical Analysis
3. Results
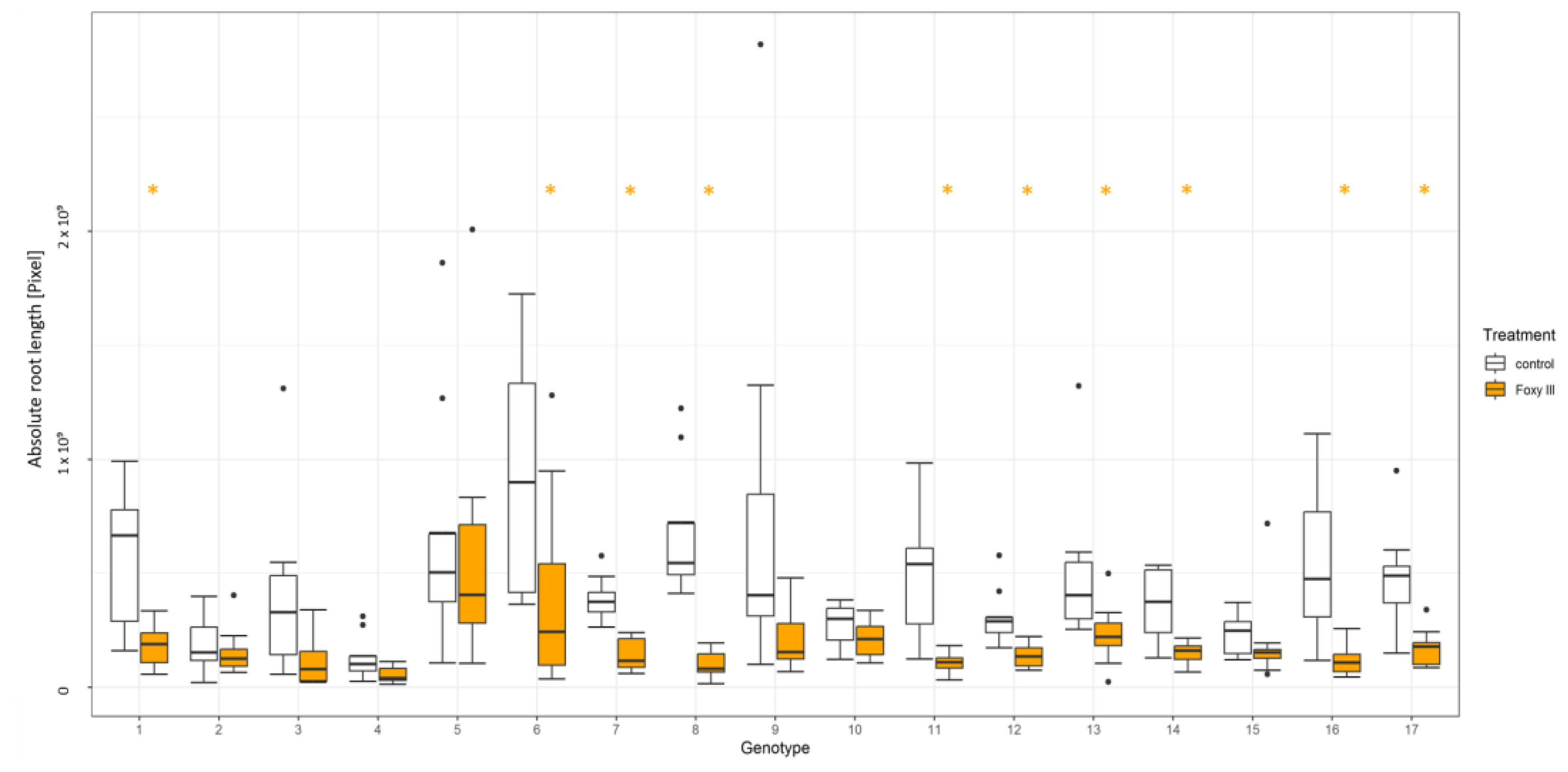
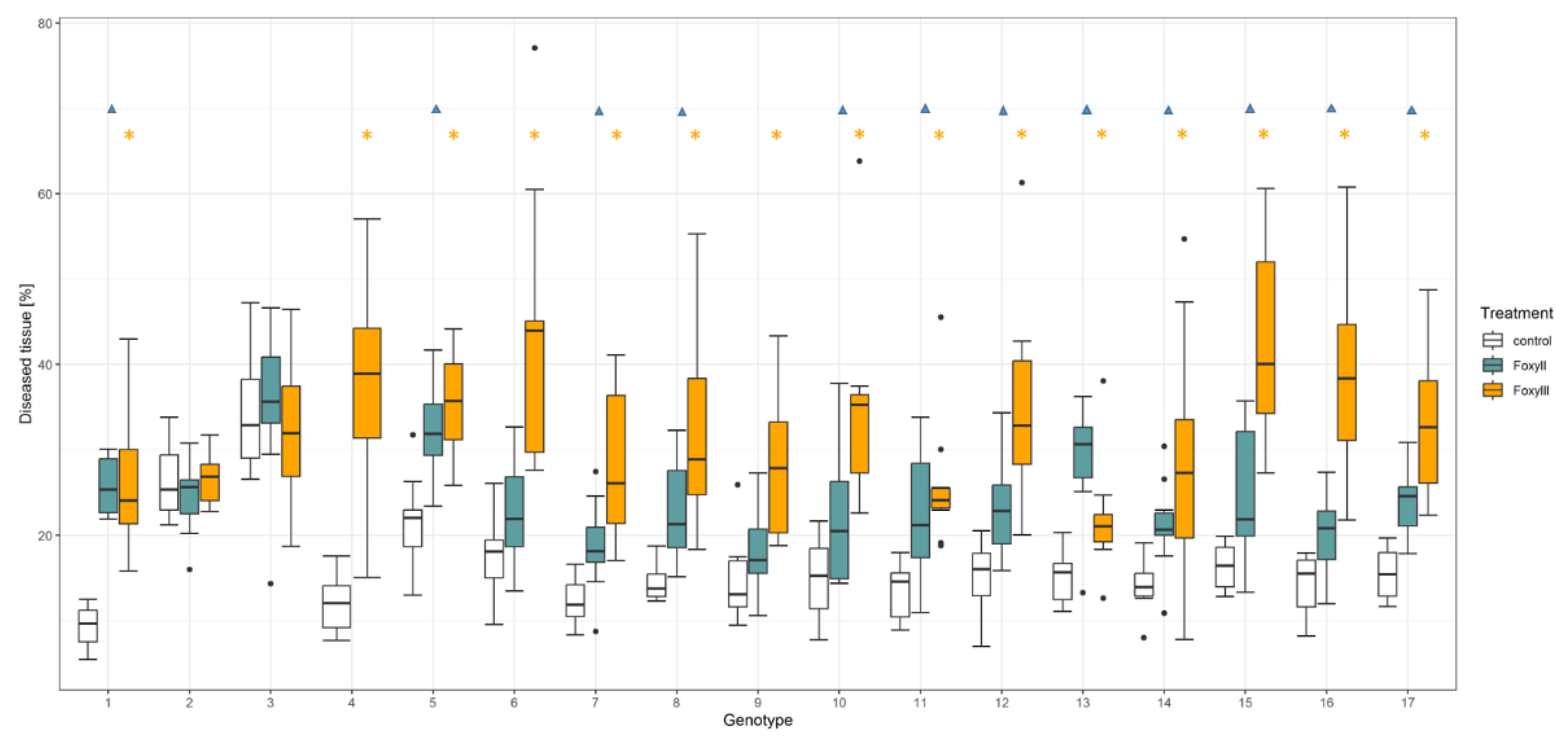
3.1. Resistance Test
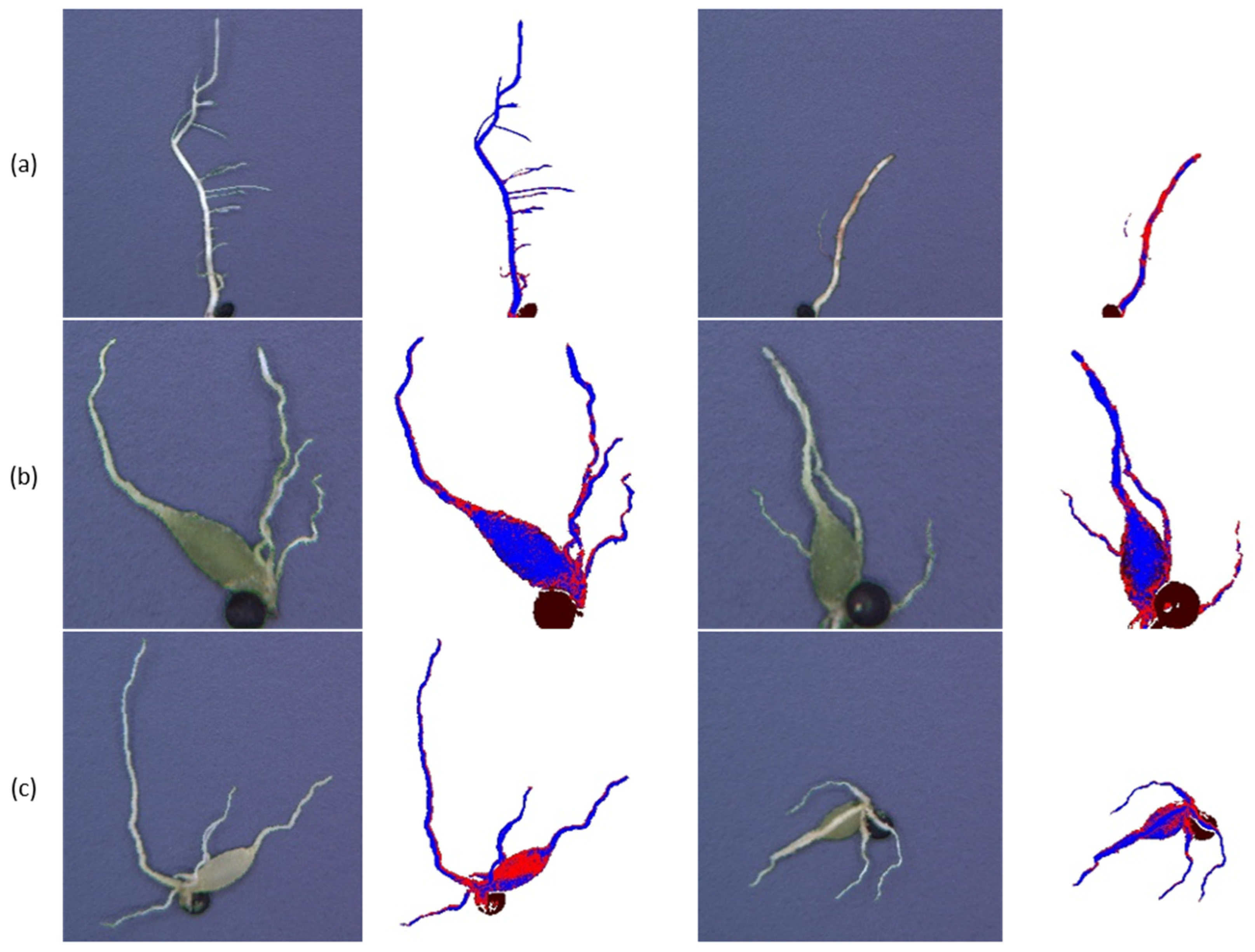
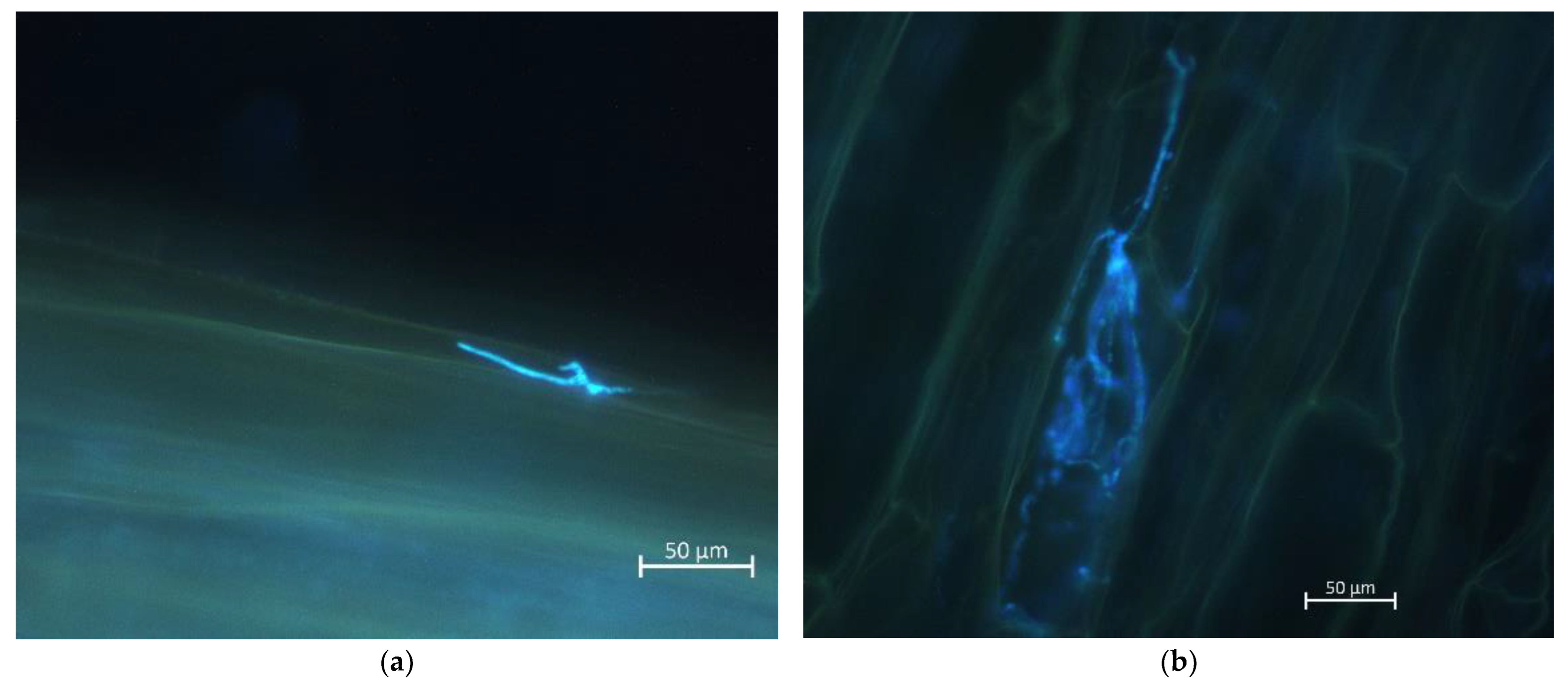
3.2. Microscopic Analyses
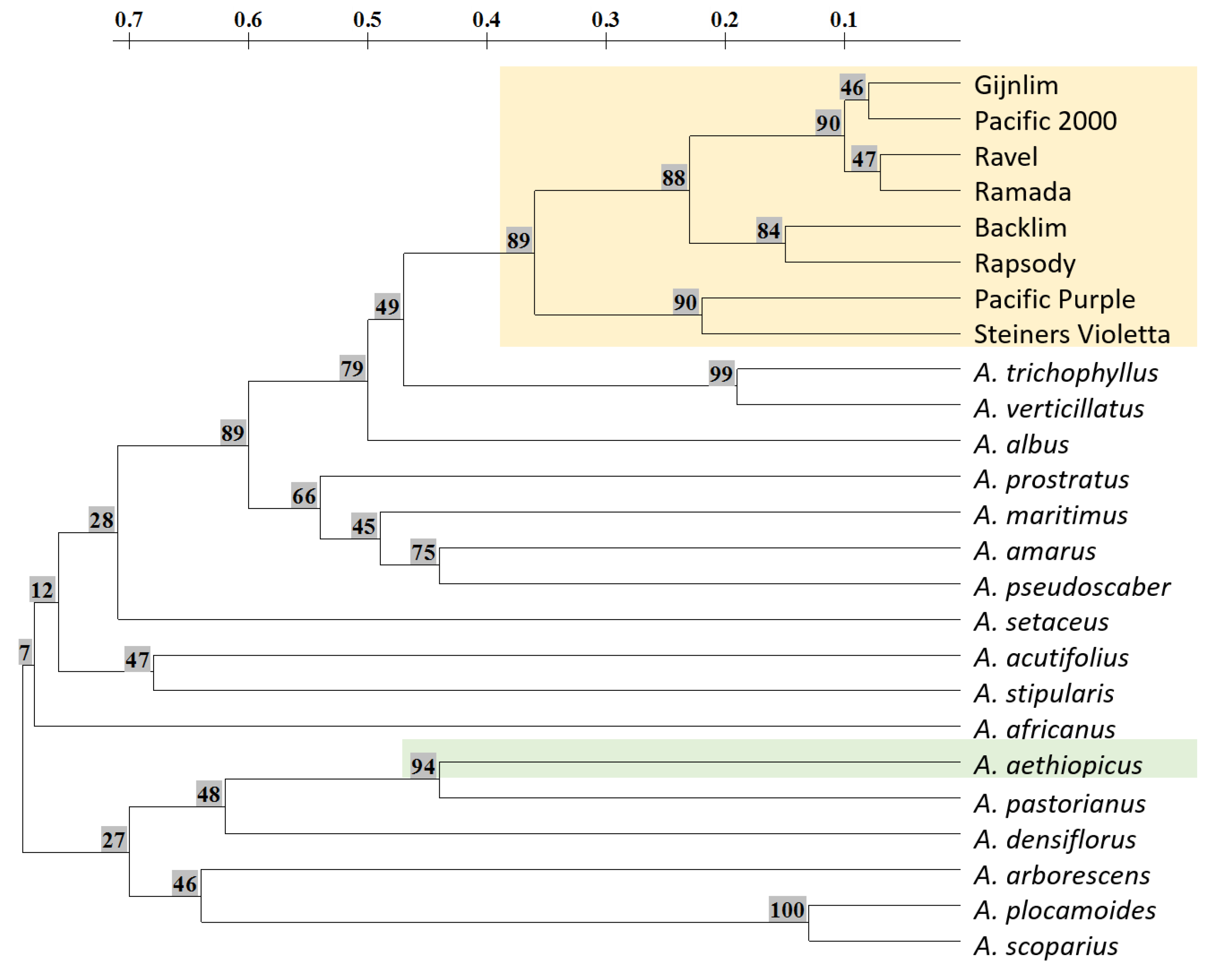
3.3. Phylogenetic Relationship
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. FAOSTAT. Available online: https://www.fao.org/faostat/en/#home (accessed on 16 September 2022).
- Bharati, J.; Kumar, S. Shatavari (Asparagus racemosus). In Phytobiotics and Animal Production; Singh, P.K., Ed.; International Books & Periodical Supply Service: Delhi, India, 2019; pp. 567–590. [Google Scholar]
- Benincasa, P.; Tei, F.; Rosati, A. Plant Density and Genotype Effects on Wild Asparagus (Asparagus acutifolius L.) Spear Yield and Quality. HortScience 2007, 42, 1163–1166. [Google Scholar] [CrossRef]
- Andrzejak, R.; Janowska, B. The occurrence of Fusarium species in white spears of Asparagus officinalis. Not. Bot. Horti Agrobot. 2021, 49, 12559. [Google Scholar] [CrossRef]
- Blok, W.J.; Bollen, G.J. Fungi on roots and stem bases of asparagus in The Netherlands: Species and pathogenicity. Eur. J. Plant Pathol. 1995, 101, 15–24. [Google Scholar] [CrossRef]
- Brizuela, A.M.; de La Lastra, E.; Marín-Guirao, J.I.; Gálvez, L.; de Cara-García, M.; Capote, N.; Palmero, D. Fusarium Consortium Populations Associated with Asparagus Crop in Spain and Their Role on Field Decline Syndrome. J. Fungi 2020, 6, 336. [Google Scholar] [CrossRef]
- Logrieco, A.; Doko, B.; Moretti, A.; Frisullo, S.; Visconti, A. Occurrence of Fumonisin B1 and B2 in Fusarium proliferatum Infected Asparagus Plants. J. Agric. Food Chem. 1998, 46, 5201–5204. [Google Scholar] [CrossRef]
- de La Lastra, E.; Marín-Guirao, J.I.; López-Moreno, F.J.; Soriano, T.; de Cara-García, M.; Capote, N. Potential inoculum sources of Fusarium species involved in asparagus decline syndrome and evaluation of soil disinfestation methods by qPCR protocols. Pest Manag. Sci. 2021, 77, 4749–4757. [Google Scholar] [CrossRef]
- Weber, Z.; Karolewski, Z.; Irzykowska, L.; Knaflewski, M.; Kosiada, T. Occurrence of Fusarium species in spears of Asparagus (Asparagus officinalis). Phytopathol. Pol. 2007, 45, 9–15. [Google Scholar]
- Apaza, W.; Talledo, A.; Casas, A. Reaction of 10 Asparagus cultivars to infested soil with Fusarium oxysporum f.sp. asparagi and Meloidogyne incognita. Acta Hortic. 2018, 1223, 233–238. [Google Scholar] [CrossRef]
- Fiume, F.; Fiume, G. Field response of some asparagus varieties to rust, Fusarium crown root rot, and violet root rot. Commun. Agric. Appl. Biol. Sci. 2003, 68, 659–671. [Google Scholar]
- Blok, W.J.; Bollen, G.J. Inoculum sources of Fusarium oxysporum f.sp. asparagi in asparagus production. Ann. Appl. Biol. 1996, 128, 219–231. [Google Scholar]
- Elmer, W.H. Management of Fusarium crown and root rot of asparagus. Crop Prot. 2015, 73, 2–6. [Google Scholar] [CrossRef]
- Reid, T.C.; Hausbeck, M.K.; Kizilkaya, K. Use of Fungicides and Biological Controls in the Suppression of Fusarium Crown and Root Rot of Asparagus Under Greenhouse and Growth Chamber Conditions. Plant Dis. 2002, 86, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Reid, T.C.; Hausbeck, M.K.; Kizilkaya, K. Effects of Sodium Chloride on Commercial Asparagus and of Alternative Forms of Chloride Salt on Fusarium Crown and Root Rot. Plant Dis. 2001, 85, 1271–1275. [Google Scholar] [CrossRef] [PubMed]
- Brizuela, A.M.; Lalak-Kańczugowska, J.; Koczyk, G.; Stępień, Ł.; Kawaliło, M.; Palmero, D. Geographical Origin Does Not Modulate Pathogenicity or Response to Climatic Variables of Fusarium oxysporum Associated with Vascular Wilt on Asparagus. J. Fungi 2021, 7, 1056. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Guerra, C.A.; Cano-Díaz, C.; Egidi, E.; Wang, J.-T.; Eisenhauer, N.; Singh, B.K.; Maestre, F.T. The proportion of soil-borne pathogens increases with warming at the global scale. Nat. Clim. Change 2020, 10, 550–554. [Google Scholar] [CrossRef]
- Kathe, L.; Krämer, R.; Budahn, H.; Pillen, K.; Rabenstein, F.; Nothnagel, T. Development of a bioassay to assess resistance to Fusarium oxysporum (Schlecht.) in asparagus (Asparagus officinalis L.). J. Phytopathol. 2019, 167, 558–566. [Google Scholar] [CrossRef]
- He, C.; Hsiang, T.; Wolyn, D.J. Activation of defense responses to Fusarium infection in Asparagus densiflorus. Eur. J. Plant Pathol. 2001, 107, 473–483. [Google Scholar] [CrossRef]
- Stephens, C.T.; De Vries, R.M.; Sink, K.C. Evaluation of Asparagus Species for resistance to Fusarium oxysporum f. sp. asparagi and F. moniliforme. HortScience 1989, 2, 365–368. [Google Scholar] [CrossRef]
- Stephens, C.T.; Wade, E.H. An In Vitro Assay to Evaluate Sources of Resistance in Asparagus spp. to Fusarium Crown and Root Rot. Plant Dis. 1988, 72, 334–337. [Google Scholar] [CrossRef]
- Lewis, G.D.; Shoemaker, P.B. Presence of Fusarium oxysporum f.asparagi on asparagus seed and Fusarium resistance in plant introduction lines of asparagus. Phytopathology 1964, 54, 128. [Google Scholar]
- Plath, S.; Krämer, R.; Lantos, E.; Nothnagel, T. Breeding programs to transmit Asparagus virus 1 resistance. Acta Hortic. 2018, 1223, 17–24. [Google Scholar] [CrossRef]
- Falavigna, A.; Alberti, P.; Casali, P.E.; Toppino, L.; Huaisong, W.; Mennella, G. Interspecific hybridization for asparagus breeding in Italy. Acta Hortic. 2008, 776, 291–298. [Google Scholar] [CrossRef]
- McCollum, G.D. Asparagus densiflorus cultivars Sprengeri and Myers cross-pollinations with A. officinalis and other species. Asparagus Res. Newsl. 1988, 6, 1–10. [Google Scholar]
- Bozzini, A. Interspecific hybridization and experimental mutagenesis in the genetic improvement of asparagus. Genet. Agrar. 1962, 16, 212–218. [Google Scholar]
- Ito, T.; Ochiai, T.; Fukuda, T.; Ashizawa, H.; Kanno, A.; Kameya, T.; Sonoda, T. Potential of interspecific hybrids in the genus Asparagus. Acta Hortic. 2008, 776, 279–284. [Google Scholar] [CrossRef]
- Marcellán, O.; Camadro, E. Formation and development of embryo and endosperm in intra- and inter-specific crosses of Asparagus officinalis and A. densiflorus cv. Sprengeri. Sci. Hortic. 1999, 81, 1621–1625. [Google Scholar] [CrossRef]
- Marcellán, O.N.; Camadro, E.L. Self- and cross-incompatibility in Asparagus officinalis and Asparagus densiflorus cv. Sprengeri. Can. J. Bot. 1996, 74, 1621–1625. [Google Scholar] [CrossRef]
- Mercati, F.; Riccardi, P.; Harkess, A.; Sala, T.; Abenavoli, M.R.; Leebens-Mack, J.; Falavigna, A.; Sunseri, F. Single nucleotide polymorphism-based parentage analysis and population structure in garden asparagus, a worldwide genetic stock classification. Mol. Breed. 2015, 35, 59. [Google Scholar] [CrossRef]
- Knaflewski, M. Genealogy asparagus cultivars. Acta Hortic. 1996, 415, 87–92. [Google Scholar] [CrossRef]
- Nothnagel, T.; Budahn, H.; Krämer, I.; Schliephake, E.; Lantos, E.; Plath, S.; Krämer, R. Evaluation of genetic resources in the genus Asparagus for resistance to Asparagus virus 1 (AV-1). Genet. Resour. Crop Evol. 2017, 64, 1873–1887. [Google Scholar] [CrossRef]
- Serfling, A.; Templer, S.E.; Winter, P.; Ordon, F. Microscopic and Molecular Characterization of the Prehaustorial Resistance against Wheat Leaf Rust (Puccinia triticina) in Einkorn (Triticum monococcum). Front. Plant Sci. 2016, 7, 1668. [Google Scholar] [CrossRef] [PubMed]
- Porebski, S.; Bailey, L.G.; Baum, B.R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Report. 1997, 15, 8–15. [Google Scholar] [CrossRef]
- Mercati, F.; Riccardi, P.; Leebens-Mack, J.; Abenavoli, M.R.; Falavigna, A.; Sunseri, F. Single nucleotide polymorphism isolated from a novel EST dataset in garden asparagus (Asparagus officinalis L.). Plant Sci. 2013, 203, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Oetting, W.S.; Lee, H.K.; Flanders, D.J.; Wiesner, G.L.; Sellers, T.A.; King, R.A. Linkage analysis with multiplexed short tandem repeat polymorphisms using infrared fluorescence and M13 tailed primers. Genomics 1995, 30, 450–458. [Google Scholar] [CrossRef]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef]
- Blok, W.J.; Zwankhuizen, M.J.; Bollen, G.J. Biological Control of Fusarium oxysporum f.sp. asparagi by Applying Non-pathogenic Isolates of F. oxysporum. Biocontrol Sci. Technol. 1997, 7, 527–542. [Google Scholar] [CrossRef]
- Pontier, D.; Balagué, C.; Roby, D. The hypersensitive response. A programmed cell death associated with plant resistance. C. R. L’académie Sci.-Ser. III-Sci. Vie 1998, 321, 721–734. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Asparagus Species | Cultivar/ Wild Relative 1 | Seed Origin 2 | Accession | Geographic Origin |
|---|---|---|---|---|---|
| 1 | A. amarus DC. | Wild relative (Pop.) | CRA | Italy | |
| 2 | A. aethiopicus L. | Wild relative (Pop.) | GG | 3333-3 | Malaga |
| 3 | A. densiflorus (Kunth) Jessop | Wild relative (Pop.) | ISR | Israel | |
| 4 | A. stipularis Forssk. | Wild relative (Pop.) | IBZ | Ibiza | |
| 5 | A. officinalis L. | Landrace (Pop.) | WUR | CGN25609 | Armenia |
| 6 | A. officinalis L. | Landrace (Pop.) | WUR | CGN25611 | Armenia |
| 7 | A. officinalis L. | cv. Argenteuil (OP) | RZ | French | |
| 8 | A. officinalis L. | cv. Grolim (F1) | LIM | The Netherlands | |
| 9 | A. officinalis L. | cv. Ivancicky (OP) | IPK | ASP 1 | unknown |
| 10 | A. officinalis L. | cv. Leistungsauslese (OP) | DS | Germany | |
| 11 | A. officinalis L. | cv. Ramada (F1) | SWS | Germany | |
| 12 | A. officinalis L. | cv. Ramires (F1) | SWS | Germany | |
| 13 | A. officinalis L. | cv. Ravel (F1) | SWS | Germany | |
| 14 | A. officinalis L. | cv. Ruhm von Braunschweig (OP) | IPK | ASP 31 | Germany |
| 15 | A. officinalis L. | cv. Schwetzinger Meisterschuss (OP) | SWS | Germany | |
| 16 | A. officinalis L. | cv. Start (OP) | IPK | ASP 3 | unknown |
| 17 | A. officinalis L. | cv. Thielim (F1) | LIM | The Netherlands |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jacobi, J.; Budahn, H.; Nothnagel, T.; König, J. Studies on the Identification of Resistance to Fusarium oxysporum (Schlecht.) in Different Genetic Backgrounds of Asparagus officinalis (L.) and Its Defense Responses. Horticulturae 2023, 9, 158. https://doi.org/10.3390/horticulturae9020158
Jacobi J, Budahn H, Nothnagel T, König J. Studies on the Identification of Resistance to Fusarium oxysporum (Schlecht.) in Different Genetic Backgrounds of Asparagus officinalis (L.) and Its Defense Responses. Horticulturae. 2023; 9(2):158. https://doi.org/10.3390/horticulturae9020158
Chicago/Turabian StyleJacobi, Julia, Holger Budahn, Thomas Nothnagel, and Janine König. 2023. "Studies on the Identification of Resistance to Fusarium oxysporum (Schlecht.) in Different Genetic Backgrounds of Asparagus officinalis (L.) and Its Defense Responses" Horticulturae 9, no. 2: 158. https://doi.org/10.3390/horticulturae9020158
APA StyleJacobi, J., Budahn, H., Nothnagel, T., & König, J. (2023). Studies on the Identification of Resistance to Fusarium oxysporum (Schlecht.) in Different Genetic Backgrounds of Asparagus officinalis (L.) and Its Defense Responses. Horticulturae, 9(2), 158. https://doi.org/10.3390/horticulturae9020158