
Effect of Low R:FR Ratio on Nitrogen Assimilation and NRT Gene Expression in Pakchoi under Excessive Nitrate Stress

 ,
,
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Effects of R:FR Ratio on Growth of Pakchoi with Excessive Nitrate Stress
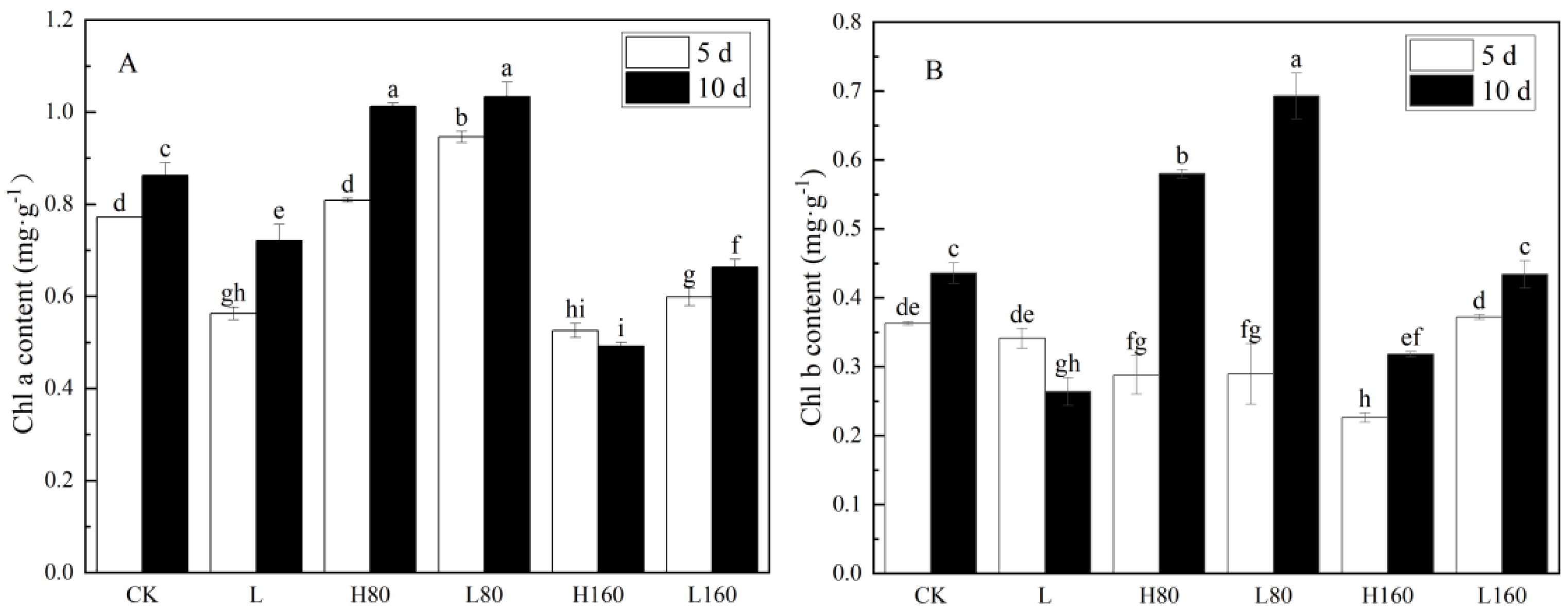
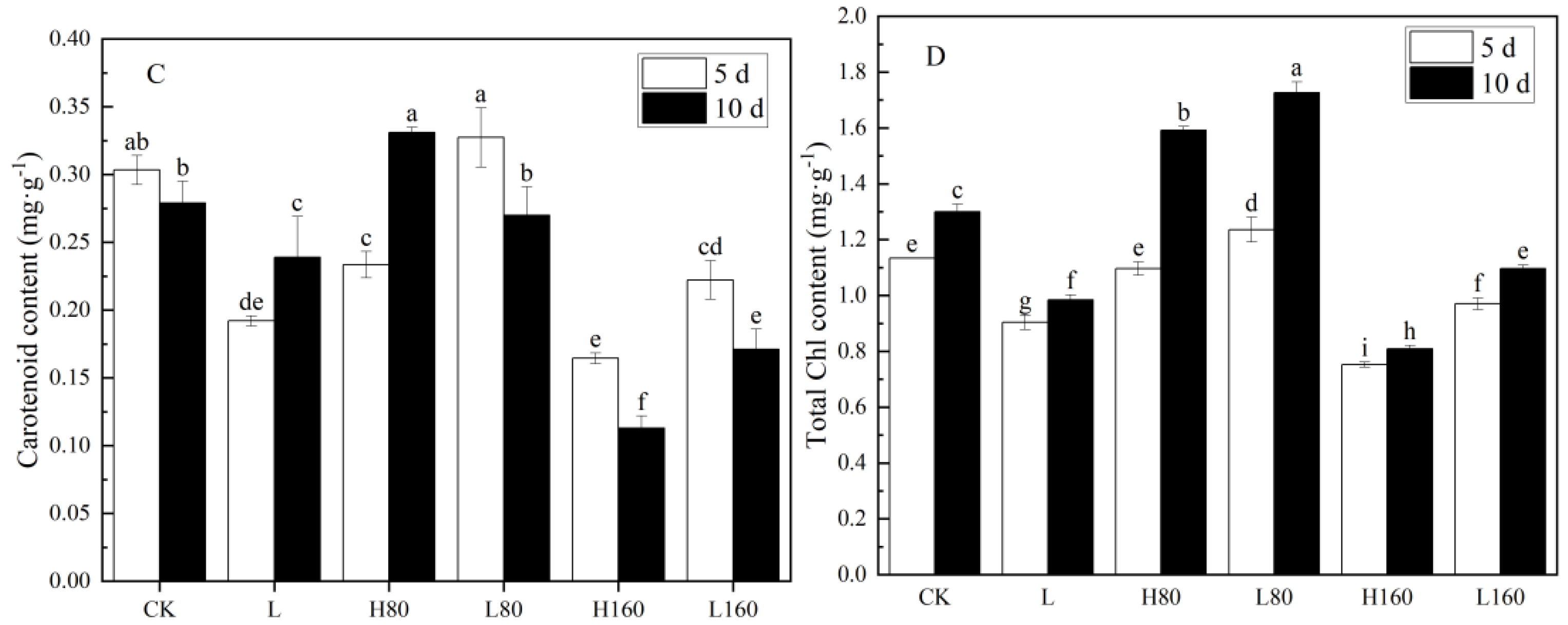
3.2. Effects of Low R:FR on Chlorophyll Content of Pakchoi with Excessive Nitrate Stress
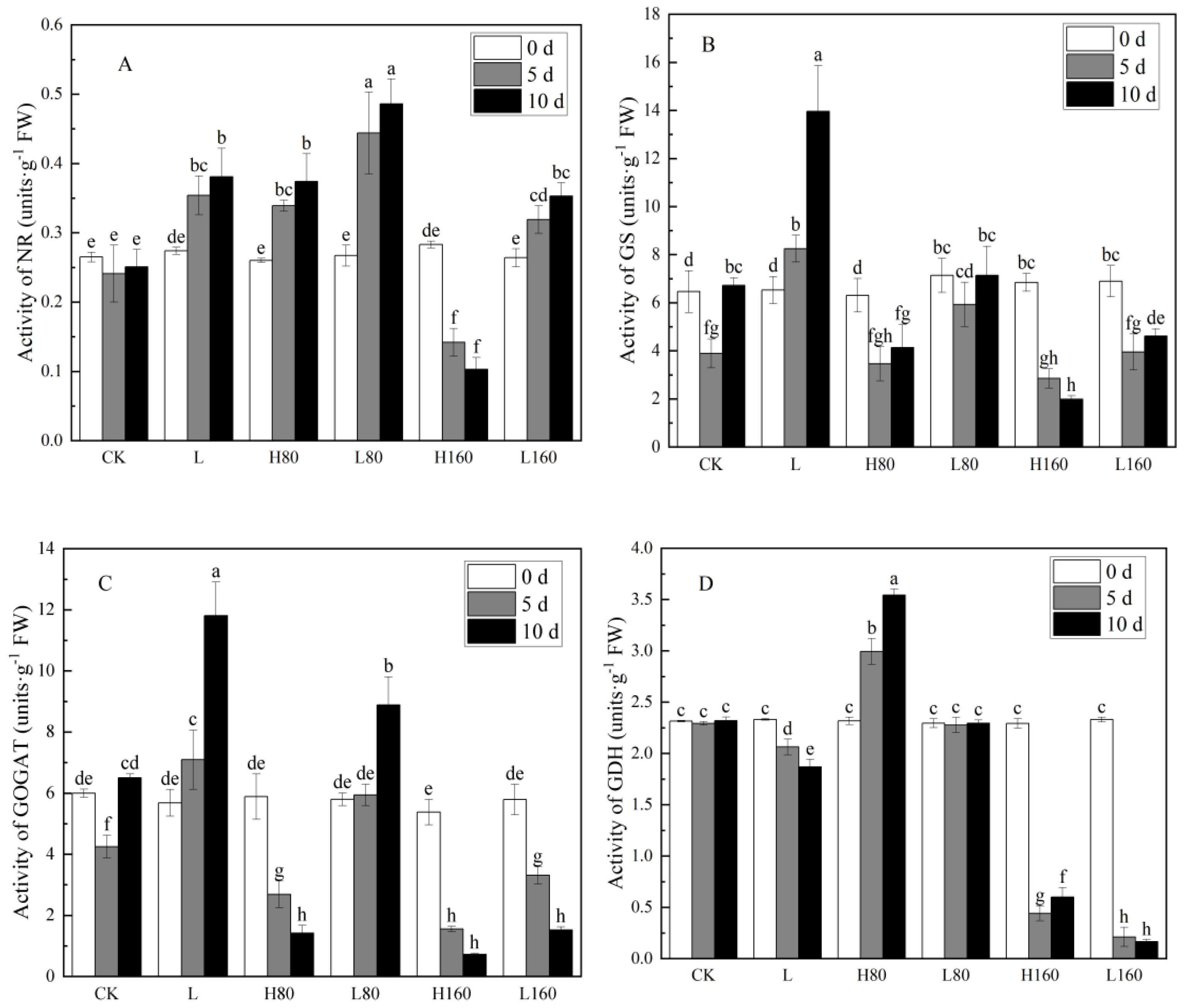
3.3. Effects of Low R:FR on Activities of Key Enzymes of Nitrogen Metabolism in Pakchoi under Excessive Nitrate Stress
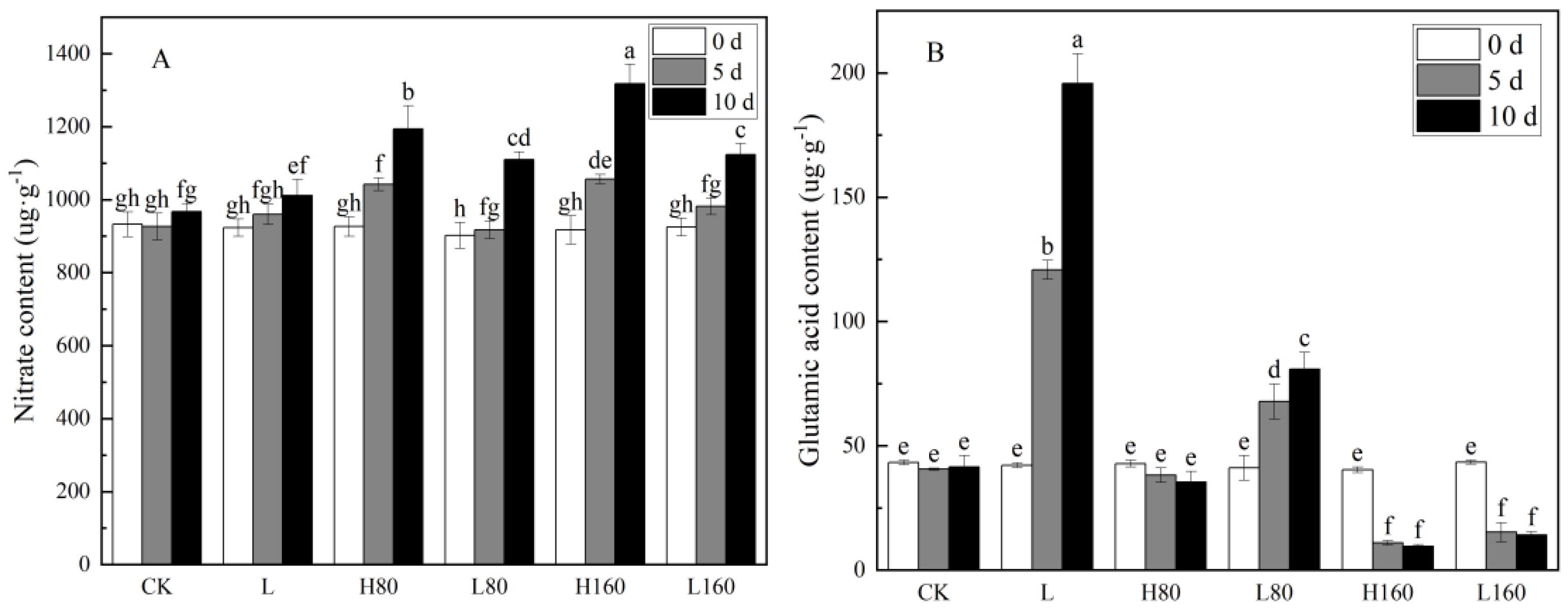
3.4. Effects of Low R:FR on Nitrate Content and Glutamic Acid Content in Pakchoi under Excessive Nitrate Stress
3.5. Correlation Analysis of Nitrogen Metabolism-Related Enzymes and Nitrogen Form
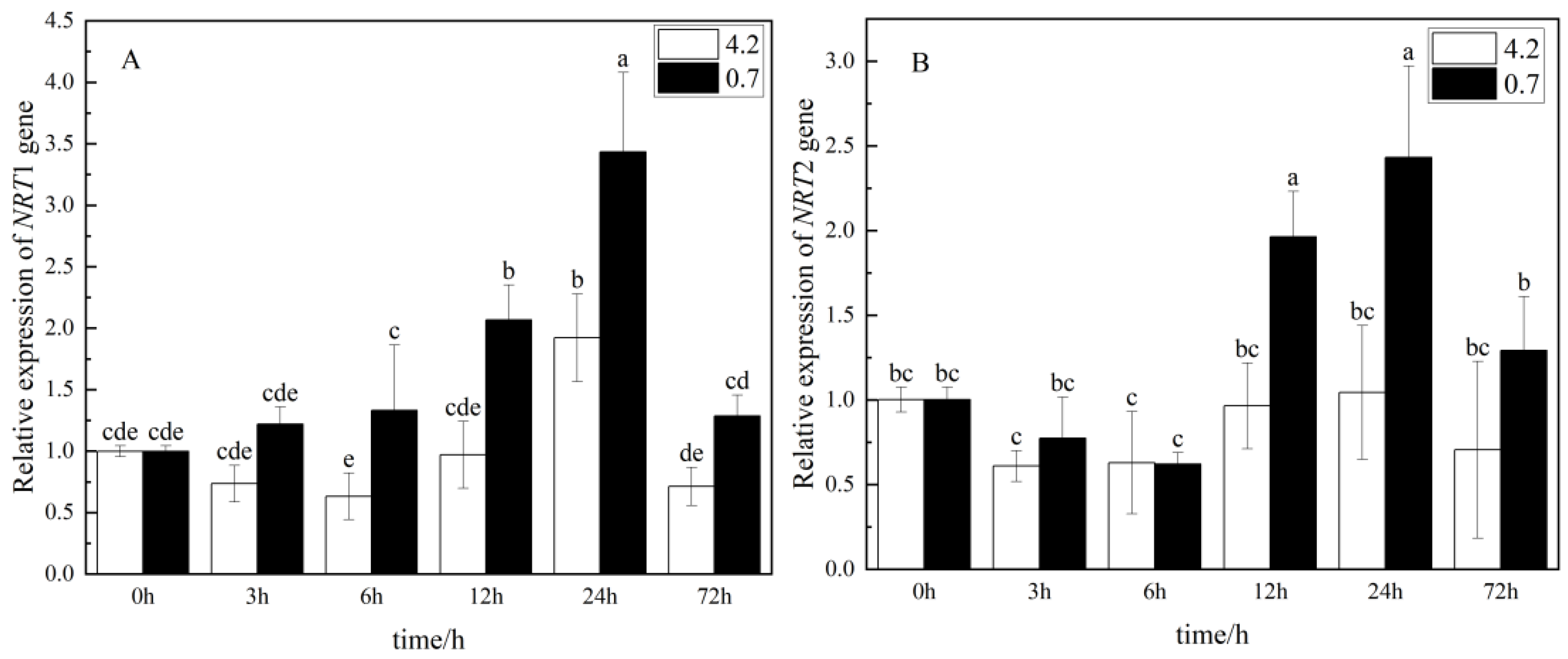
3.6. Effect of Light and Quality Ratio of LED on NRT Gene Expression in Pakchoi under Excessive Nitrate Stress
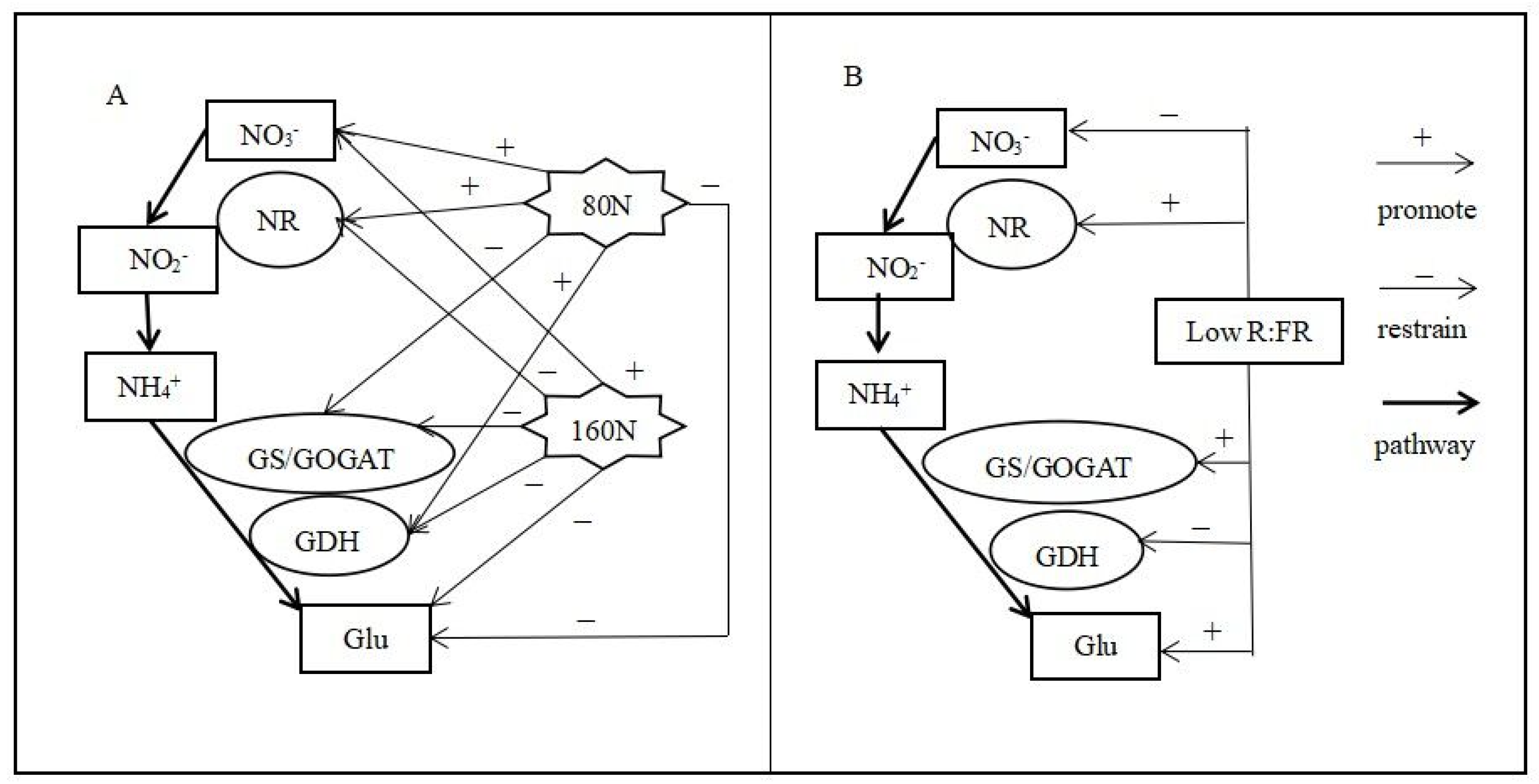
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yang, P.; Zhao, Z.H. Effects of nitrogen fertilizer on growth and nitrate accumulation of pakchoi. China Fruit Veget. 2017, 37, 25–28. [Google Scholar]
- Yang, X.Y.; Yang, J.S. The impact of nitrogen supply level on the growth and nitrate accumulation of pakchoi. Plant Nutr. Fert. Sci. 2007, 1, 160–163. [Google Scholar]
- Chen, S.Q.; Jiang, Y.T. Cause, harm and treatment measures of secondary salinization of facility soil. Liaoning Huagong. 2018, 47, 1146–1148. [Google Scholar]
- Sun, Z.D.; Han, X.R.; Peng, J.; Fan, F.; Zhang, Q.G. Effect of exogenous nitric oxide on PSII function and distribution and utilization of luminous energy in tomato seedlings under stress of Ca(NO3)2. J. Nucl. Agric. Sci. 2016, 30, 2451–2459. [Google Scholar]
- Qi, X.L.; Zou, Z.R.; Yang, R. Alleviative effect of exogenous ALA on lettuce plants under NaCl stress. Acta Agric. Boreali-Occident. Sin. 2008, 17, 202–206. [Google Scholar]
- Zhou, C.; Zheng, C. Effects of different nitrogen levels on yield and quality of pakchoi. Inform.Agric.Sci.Tech. 2006, 2, 36–37. [Google Scholar]
- Lu, Y.; Shu, S.; Zhu, W.M.; Guo, S.R. Effects of exogenous spermidine on growth, photosynthesis and quality of Chinese cabbage under calcium nitrate stress. Xibei Zhiwu Xuebao 2015, 35, 0787–0792. [Google Scholar]
- Liu, S.W.; Wang, N.; Irfan, M.; Xu, J.Y.; Zhang, Y.J.; Cai, G.X. Effect of fermentation broth of endoph ytic fungi on physiological and biochemical characteristics of tomato seedling under calcium nitrate stress. Iran. J. Sci. Technol. Trans. Sci. Trans. A Sci. 2019, 43, 1427–1432. [Google Scholar]
- Pang, Q.Q.; Chen, R.Y.; Liu, H.C.; Song, S.W.; Su, W.; Wen, G.W. Effects of fulvic acid on the growth of pakchoi and the activities of enzymes related to nitrogen metabolism under nitrate stress. J. Zhejiang Agric. Univ. 2015, 27, 2136–2140. [Google Scholar]
- Cao, K.; Yu, J.; Xu, D.W.; Ai, K.Q.; Bao, E.C.; Zou, Z.R. Exposure to lower red to far-red light ratios improve tomato tolerance to salt stress. BMC Plant Biol. 2018, 18, 92. [Google Scholar] [CrossRef]
- Park, Y.; Runkle, E.S. Far-red radiation promotes growth of seedings by increasing leaf expansion and whole plant net assessment. Environ. Exp. Bot. 2017, 136, 41–49. [Google Scholar] [CrossRef]
- Duong, T.N.; Takamura, T.; Watanabe, H.; Okamoto, K.; Tanaka, M. Responses of strawberry plantlets cultured in vitro under superbright red and blue light-emitting diodes (LEDs). Appl. Fundam. Aspects Plant Cell Tissue Organ. Cult. 2003, 73, 43–52. [Google Scholar]
- Gao, D.G.; Yu, J.; Zou, Z.R.; Cao, K.; Bao, E.C.; Meng, L.L.; Li, S.Z.; Ye, L. Effects of different ratios of red light to far-red light on photosynthetic capacity of tomato under salt stress. J.Anhui Agric.Univ. 2021, 48, 578–583. [Google Scholar]
- Zhang, L.W. Effects of Light Quality on Physiological Characteristics and Quality of Three Kinds of Sprouting Vegetables. Master’s Thesis, Shandong Agricultural University, Jinan, China, 2010. [Google Scholar]
- Zhen, S.Y.; Iersel, M.V. Far-red light is needed for efficient photochemistry and photosynthesis. Plant Physiol. 2017, 209, 115–122. [Google Scholar] [CrossRef]
- Ning, Y.; Deng, H.H.; Li, Q.M.; Mi, Q.H.; Han, B.; Ai, X.Z. Effects of red and blue light quality on carbon and nitrogen metabolism and key enzyme activities of celery. Acta Phytopsoil. Sin. 2015, 1, 112–118. [Google Scholar]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef]
- Zhi, L.; Shi, H.Z.; Liu, S.G.; Wang, D.Z.; Zu, C.L.; Yang, Y.F. Changes of earbon nitrogen metabolism of flue-cured tobacco with sweet aroma in south anhui under different fertilization rates. J. Exp. Bot. 2015, 66, 2055–2065. [Google Scholar]
- Wang, Y.Y.; Hsu, P.K.; Tsay, Y.F. Uptake, allocation and signaling of nitrate trends. Plant Sci. 2012, 17, 458–467. [Google Scholar]
- Zhao, S.P.; Zhang, R.L.; Xu, M.F.; Zheng, J.C. Study on the mechanism of the effect of ammonium nitrate ratio on nitrate accumulation in different genotypes of pakchoi. Soils. 2011, 43, 32–38. [Google Scholar]
- Sandrine, F.R.; ‚Erwan, L.D.; ‚Philippe, L.; James, H.M.; Alain, O. Effect of nitrate pulses on BnNRT1 and BnNRT2 genes: mRNA levels and nitrate in flux rates in relation to the duration of N deprivation in Brassica napus L. J. Exp. Bot. 2002, 53, 1711–1721. [Google Scholar]
- Dong, S.J. Effect of LED Light Supplement on Pepper Seedling Growth and Colonization of Arbuscular Mycorrhizal Fungi. Master’s Thesis, Zhejiang University, Hangzhou, China, 2021. [Google Scholar]
- Chen, L.B.; Huang, J.; Liu, Q.L.; Li, Z.L.; Chen, X.; Han, J.X.; Gan, Y.R.; He, Y.X.; Jiang, C.X.; Tang, Y.X.; et al. Low R:FR ratio affects pakchoi’s growth and nitrate content under excess nitrate stress. Horticulturae 2022, 8, 186. [Google Scholar] [CrossRef]
- Yu, J. Effect of Different Ratio of Red Light to Far Red Light on Salt Tolerance of Tomato. Northwest. Master’s Thesis, Agriculture and Forestry University, Shanxi, China, 2018. [Google Scholar]
- Zhou, Q.F.; Wang, E.Z. High-efficient horticultural facilities and horticultural lighting source. J. Chang. Veg. 1999, 1, 1–4. [Google Scholar]
- Lichtenhaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Xiong, Q.E. A Experimental Course of Plant Physiology; Sichuan Science and Technology Press: Chengdu, China, 2003. [Google Scholar]
- Lin, C.C.; Kao, C.H. Disturbed ammonium assimilation is associated with growth inhibition of roots in rice seedlings caused by NaCl. Plant Growth Regul. 1996, 18, 233–238. [Google Scholar] [CrossRef]
- Jin, O.; Chen, B. Comparison of determination methods of glutamate content. Chin. J. Biochem. Pharm. 1993, 59–60. [Google Scholar]
- Zhao, S.P.; Zhang, Y.Z.; Ye, X.Z.; Zheng, J.C. Research on the mechanism of different nitrate accumulationin Chinese cabbage with different genotypes. Plant Nutr. Fert. Sci. 2021, 16, 681–688. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR methods. Acta Agron. Sci. 2002, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.L.; Li, H.R.; Guo, S.Y.; Wang, S.M.; Shi, H.Z.; Han, Q.Q.; Bao, A.K.; Qing, M.A. Research advances in higher plant adaptation to salt stress. Acta Prataculturae Sin. 2015, 24, 220–236. [Google Scholar]
- Zhou, X.T.; Li, Z.L.; He, J.J.; Wang, X.Y.; Liu, Q.L.; Huang, J.; Xie, Y.D.; He, Z.Q. Effects of red to far-red light ratio on growth and photosynthetic characteristics of tomato seedlings under calcium nitrate stress. Photosynthetica 2021, 59, 625–632. [Google Scholar] [CrossRef]
- Li, Z.L.; Chen, G.Y.; Gao, F.Q.; Luo, J.P.; Li, C.W.; He, Z.Q.; Zhou, X.T. Effects of different light quality ratio on growth and photosynthesis capacity in pakchoi under excess nitrate stress. IOP Conf. Ser. Earth Environ. Sci. 2021, 621, 012105. [Google Scholar] [CrossRef]
- Gao, X.X.; Li, R.C.; Wen, X.Z.; Miao, Y.X. Effect of the ratio of red light and far-red light on the growth and photosynthetic characteristics of cucumber seedlings under salt stress. Shandong Agric. Sci. 2021, 53, 36–41. [Google Scholar]
- Xu, F.F.; Xu, P.; Hu, Z.T.; Zhao, J. Photosynthetic Physiological Response of pakchoi to Salt Stress. Mol. Plant Breed. 2018, 16, 3327–3332. [Google Scholar]
- Ai, K.Q.; Su, H.; Zh, H.J.; Zhou, B.; Cao, K.; Zou, Z.R. Effects of different red and far-red light on chlorophyll synthesis of tomato under salt stress. North. Hortic. 2019, 1, 14–22. [Google Scholar]
- Wang, Z.R.; Wang, H.Y.; Deng, H.F.; Xv, C.Q. Regulation of transcription factor HY5 on plant light morphogenesis and nitrogen metabolism. China Veget. 2018, 5, 20–27. [Google Scholar]
- Chi, S.L.; Yang, Y.; Xu, W.H.; Chen, X.G.; Chen, Y.Q.; Xie, W.W.; Xiong, S.X.; Wang, Z.Y.; Xie, D.T. Correlation between nitrate content in pakchoi and NO3-: NH4+and key enzymes of nitrogen metabolism. Food Sci. 2015, 36, 70–77. [Google Scholar]
- Zhang, Y.Y.; Song, X.F.; Xu, J.B.; Zhou, Q.; Liu, H.Q.; Wang, Y.; Zhang, Z.Y.; Song, H.L. Effects of different nitrogen concentrations on the growth and physiological indicators of water hyacinth, especially the activities of key enzymes of nitrogen metabolism. Plant Resour. Environ. 2021, 30, 39–46. [Google Scholar]
- Yu, Y.C.; Xu, T.; Li, X.; Tang, J.; Ma, D.F.; Li, Z.Y.; Sun, J. NaCl-induced changes of ion homeostasis and nitrogen metabolism in two sweet potato (Ipomoea batatas L.) cultivars exhibit different salt tolerance at adventitious root stage. Environ. Exp. Bot 2016, 129, 23–36. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | White Light | Far-Red Light | Additional 80 mmol·L−1 NO3− Was Added | Additional 160 mmol·L−1 NO3− Was Added |
|---|---|---|---|---|
| CK | + | |||
| L | + | + | ||
| H80 | + | + | ||
| L80 | + | + | + | |
| H160 | + | + | ||
| L160 | + | + | + |
| Treatments | Fresh Weight (g/plant) | Dry Weight (g/plant) | ||
|---|---|---|---|---|
| Aboveground | Belowground | Aboveground | Belowground | |
| CK | 15.422 ± 1.480 a | 0.945 ± 0.130 a | 0.675 ± 0.040 a | 0.072 ± 0.010 a |
| L | 15.606 ± 1.220 a | 1.055 ± 0.140 a | 0.691 ± 0.020 a | 0.081 ± 0.011 a |
| H80 | 8.67 ± 0.960 cd | 0.635 ± 0.040 c | 0.398 ± 0.010 c | 0.049 ± 0.003 c |
| L80 | 12.882 ± 1.810 b | 0.794 ± 0.020 b | 0.484 ± 0.020 b | 0.061 ± 0.002 b |
| H160 | 6.834 ± 0.660 d | 0.443 ± 0.010 d | 0.293 ± 0.020 e | 0.033 ± 0.004 d |
| L160 | 9.224 ± 0.530 c | 0.681 ± 0.010 bc | 0.337 ± 0.020 d | 0.054 ± 0.004 bc |
| Index | NO3− | Glu | NR | GDH | GS | GOGAT |
|---|---|---|---|---|---|---|
| NO3− | 1.000 | |||||
| Glu | −0.546 | 1.000 | ||||
| NR | −0.439 | 0.458 | 1.000 | |||
| GDH | −0.265 | 0.239 | 0.430 | 1.000 | ||
| GS | −0.736 | 0.965 ** | 0.490 | 0.208 | 1.000 | |
| GOGAT | −0.730 | 0.891 * | 0.516 | 0.271 | 0.921 ** | 1.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, X.; Gan, Y.; Jiang, C.; Chen, Q.; Chen, X.; Chen, L.; He, Z. Effect of Low R:FR Ratio on Nitrogen Assimilation and NRT Gene Expression in Pakchoi under Excessive Nitrate Stress. Horticulturae 2023, 9, 159. https://doi.org/10.3390/horticulturae9020159
Zhou X, Gan Y, Jiang C, Chen Q, Chen X, Chen L, He Z. Effect of Low R:FR Ratio on Nitrogen Assimilation and NRT Gene Expression in Pakchoi under Excessive Nitrate Stress. Horticulturae. 2023; 9(2):159. https://doi.org/10.3390/horticulturae9020159
Chicago/Turabian StyleZhou, Xiaoting, Yirong Gan, Chenxiang Jiang, Qiang Chen, Xu Chen, Libang Chen, and Zhongqun He. 2023. "Effect of Low R:FR Ratio on Nitrogen Assimilation and NRT Gene Expression in Pakchoi under Excessive Nitrate Stress" Horticulturae 9, no. 2: 159. https://doi.org/10.3390/horticulturae9020159
APA StyleZhou, X., Gan, Y., Jiang, C., Chen, Q., Chen, X., Chen, L., & He, Z. (2023). Effect of Low R:FR Ratio on Nitrogen Assimilation and NRT Gene Expression in Pakchoi under Excessive Nitrate Stress. Horticulturae, 9(2), 159. https://doi.org/10.3390/horticulturae9020159