

Cladosporium Species: The Predominant Species Present on Raspberries from the U.K. and Spain and Their Ability to Cause Skin and Stigmata Infections
 , , ,
, , ,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Determining Cladosporium Species Present on Raspberry Fruit
2.1.1. Isolate Collection
2.1.2. DNA Extraction, Amplification and Sequencing
2.1.3. Phylogenetic Analysis
2.2. Susceptibility of Fruit to Cladosporium Skin Lesions
2.2.1. Experimental Design and Treatments
2.2.2. Fruit Inoculation
2.2.3. Infection Assessment
2.2.4. Statistical Analyses
2.3. Susceptibility of Stigmata to Cladosporium Infection
2.3.1. Experimental Design
2.3.2. Fruit Inoculation
2.3.3. Assessment
2.3.4. Statistical Analyses
3. Results
3.1. Determination of Predominant Cladosporium Species on Raspberry Fruit
3.2. Fruit Susceptibility to Cladosporium Skin Lesions
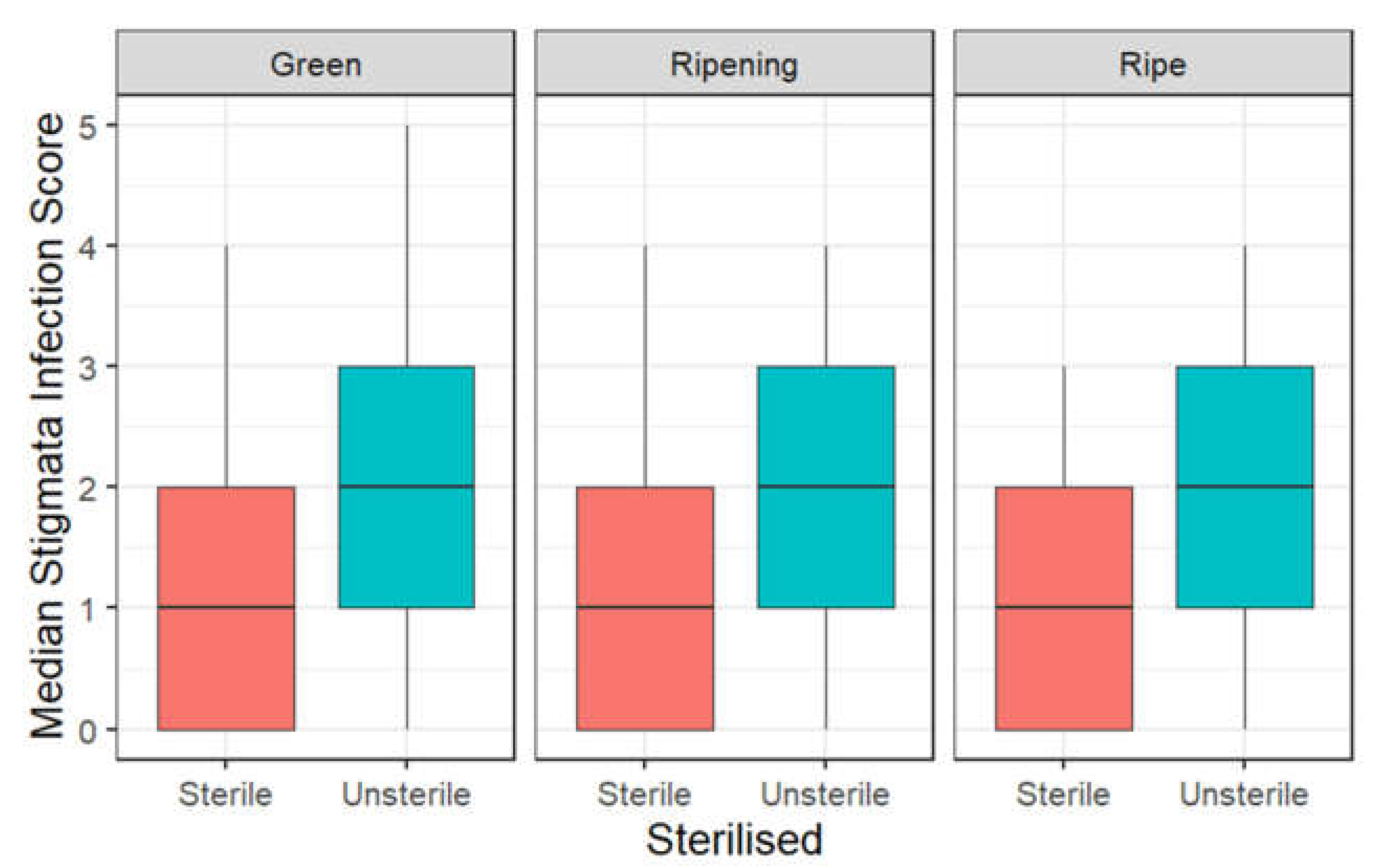
3.3. Infection of Stigmata by Cladosporium
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Department for Environment Food & Rural Affairs. Horticulture Statistics. 2020. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/1003935/hort-report-20jul21.pdf (accessed on 26 November 2021).
- Xu, X.; Wedgwood, E.; Berrie, A.M.; Allen, J.; O’Neill, T.M. Management of raspberry and strawberry grey mould in open field and under protection. A review. Agron. Sustain. Dev. 2011, 32, 531–543. [Google Scholar] [CrossRef]
- O’Neill, T.; Wedgwood, E.; Berrie, A.M.; Allen, J.; Xu, X. Managing grey mould on raspberry grown under protection without use of fungicides during flowering and fruiting. Agron. Sustain. Dev. 2012, 32, 673–682. [Google Scholar] [CrossRef]
- Swett, C.L.; Hamby, K.A.; Hellman, E.M.; Carignan, C.; Bourret, T.B.; Koivunen, E.E. Characterizing members of the Cladosporium cladosporioides species complex as fruit rot pathogens of red raspberries in the mid-Atlantic and co-occurrence with Drosophila Suzukii (Spotted wing drosophila). Phytoparasitica 2019, 47, 415–428. [Google Scholar] [CrossRef]
- Dennis, C. Effect of pre-harvest fungicides on the spoilage of soft fruit after harvest. Ann. Appl. Biol. 1975, 81, 227–234. [Google Scholar] [CrossRef]
- Jersch, S.; Scherer, C.; Huth, G.; Schlösser, E. Proanthocyanidins as basis for quiescence of Botrytis cinerea in immature strawberry fruits/Proanthocyanidine als ursache der quieszenz von Botrytis Cinerea in unreifen erdbeerfrüchten in unreifen erdbeerfrüchten. zeitschrift für pflanzenkrankheiten und pflanzenschutz. J. Plant Dis. Prot. 1989, 96, 365–378. [Google Scholar]
- Carisse, O.; Bouchard, J. Age-related susceptibility of strawberry leaves and berries to infection by Podosphaera aphanis. Crop Prot. 2010, 29, 969–978. [Google Scholar] [CrossRef]
- Nam, M.H.; Park, M.S.; Kim, H.S.; Kim, T.I.; Kim, H.G. Cladosporium cladosporioides and C. tenuissimum cause blossom blight in strawberry in Korea. Mycobiology 2015, 43, 354–359. [Google Scholar] [CrossRef]
- Gubler, W.D.; Feliciano, A.J.; Bordas, A.C.; Civerolo, E.C.; Melvin, J.A.; Welch, N.C. First report of blossom blight of strawberry caused by Xanthomonas fragariae and Cladosporium cladosporioides in California. Plant Dis. 1999, 83, 400. [Google Scholar] [CrossRef]
- Extract-N-Amp™ Plant PCR Kits Protocol. Available online: https://www.sigmaaldrich.com/GB/en/technical-documents/protocol/genomics/pcr/extract-n-amp-plant-pcr-kits-protocol (accessed on 18 March 2022).
- Bensch, K.; Braun, U.; Groenewald, J.Z.; Crous, P.W. The genus Cladosporium. Stud. Mycol. 2012, 72, 1–401. [Google Scholar]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. GlmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- R Core Team. R Foundation for Statistical Computing. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 26 November 2021).
- Christensen, R.H.B. Ordinal—Regression Models for Ordinal Data. 2019. Available online: http://www.cran.r-project.org/package=ordinal/ (accessed on 29 November 2019).
- Ayoubi, N.; Soleimani, M.J.; Zare, R. Cladosporium species, a new challenge in strawberry production in Iran. Phytopathol. Mediterr. 2017, 56, 486–493. [Google Scholar]
- Temperini, C.V.; Pardo, A.G.; Pose, G.N. Diversity of airborne Cladosporium species isolated from agricultural environments of northern Argentinean Patagonia: Molecular characterization and plant pathogenicity. Aerobiologia 2018, 34, 227–239. [Google Scholar] [CrossRef]
- Ellis, M.B. Dematiaceous Hyphomycetes, 1st ed.; Commonwealth Mycological Institute: Kew, UK, 1971; pp. 308–319. [Google Scholar]
- Harvey, R. Air-spora studies at Cardiff: I. Cladosporium. Trans. Br. Mycol. Soc. 1967, 50, 479–495. [Google Scholar] [CrossRef]
- Pasanen, A.L.; Niininen, M.; Kalliokoski, P.; Nevalainen, A.; Jantunen, M.J. Airborne Cladosporium and other fungi in damp versus reference residences. Atmos. Environ. 1992, 26, 121–124. [Google Scholar] [CrossRef]
- Anees-Hill, S.; Douglas, P.; Pashley, C.H.; Hansell, A.; Marczylo, E.L. A systematic review of outdoor airborne fungal spore seasonality across Europe and the implications for health. Sci. Total Environ. 2022, 818, 151716. [Google Scholar] [CrossRef]
- Briceño, E.X.; Latorre, B.A. Characterization of Cladosporium rot in grapevines, a problem of growing importance in Chile. Plant Dis. 2008, 92, 1635–1642. [Google Scholar] [CrossRef]
- Kozhar, O.; Peever, T.L. How does Botrytis cinerea infect red raspberry? Phytopathology 2018, 108, 1287–1298. [Google Scholar] [CrossRef]
- Barbosa, M.A.G.; Rehn, K.G.; Menezes, M.; Mariano, R.L.R. Antagonism of Trichoderma species on Cladosporium herbarum and their enzimatic characterization. Braz. J. Microiol. 2001, 32, 98–104. [Google Scholar] [CrossRef]
- El-Dawy, E.G.A.E.M.; Gherbawy, Y.A.; Hussein, M.A. Morphological, molecular characterization, plant pathogenicity and biocontrol of Cladosporium complex groups associated with faba beans. Sci. Rep. 2021, 11, 14183. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cladosporium Species | U.K. Fruit | Spanish Fruit |
|---|---|---|
| C. cladosporioides | 41.5 | 84.6 |
| C. fusiforme | 12.2 | 0.0 |
| C. sphaerospermum | 14.6 | 15.4 |
| C. europaeum | 14.6 | 0.0 |
| C. limoniforme | 9.8 | 0.0 |
| C. ramotellum | 7.3 | 0.0 |
| Sterilisation Treatment | Green | Ripening | Ripe |
|---|---|---|---|
| Sterilised | 66.4 (s.e. ± 6.9) | 74.5 (s.e. ± 12.3) | 69.9 (s.e. ± 16.5) |
| Unsterilised | 88.3 (s.e. ± 6.8) | 87.0 (s.e. ± 11.1) | 90.6 (s.e. ± 8.6) |
| Inoculum | Green | Ripening | Ripe |
|---|---|---|---|
| Control | 65.7 (s.e. ± 10.5) | 68.7 (s.e. ± 14.3) | 70.1 (s.e. ± 14.7) |
| Isolate 1 | 79.1 (s.e. ± 9.1) | 81.2 (s.e. ± 10.7) | 82.2 (s.e. ± 10.8) |
| Isolate 2 | 81.0 (s.e. ± 8.7) | 82.9 (s.e. ± 10.1) | 83.9 (s.e. ± 10.7) |
| Isolate 3 | 87.9 (s.e. ± 6.4) | 89.3 (s.e. ± 7.3) | 89.9 (s.e. ± 7.2) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farwell, L.H.; Deakin, G.; Harris, A.L.; Fagg, G.; Passey, T.; Verheecke-Vaessen, C.; Magan, N.; Xu, X. Cladosporium Species: The Predominant Species Present on Raspberries from the U.K. and Spain and Their Ability to Cause Skin and Stigmata Infections. Horticulturae 2023, 9, 128. https://doi.org/10.3390/horticulturae9020128
Farwell LH, Deakin G, Harris AL, Fagg G, Passey T, Verheecke-Vaessen C, Magan N, Xu X. Cladosporium Species: The Predominant Species Present on Raspberries from the U.K. and Spain and Their Ability to Cause Skin and Stigmata Infections. Horticulturae. 2023; 9(2):128. https://doi.org/10.3390/horticulturae9020128
Chicago/Turabian StyleFarwell, Lauren Helen, Greg Deakin, Adrian Lee Harris, Georgina Fagg, Thomas Passey, Carol Verheecke-Vaessen, Naresh Magan, and Xiangming Xu. 2023. "Cladosporium Species: The Predominant Species Present on Raspberries from the U.K. and Spain and Their Ability to Cause Skin and Stigmata Infections" Horticulturae 9, no. 2: 128. https://doi.org/10.3390/horticulturae9020128
APA StyleFarwell, L. H., Deakin, G., Harris, A. L., Fagg, G., Passey, T., Verheecke-Vaessen, C., Magan, N., & Xu, X. (2023). Cladosporium Species: The Predominant Species Present on Raspberries from the U.K. and Spain and Their Ability to Cause Skin and Stigmata Infections. Horticulturae, 9(2), 128. https://doi.org/10.3390/horticulturae9020128