
Tomato Accumulates Cadmium to a Concentration Independent of Plant Growth



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials and Design
2.2. Determination of Growth Parameters
2.3. Determination of Cd Concentration
2.4. Statistical Analyses
3. Results
3.1. Accumulation of Cd in Tomato
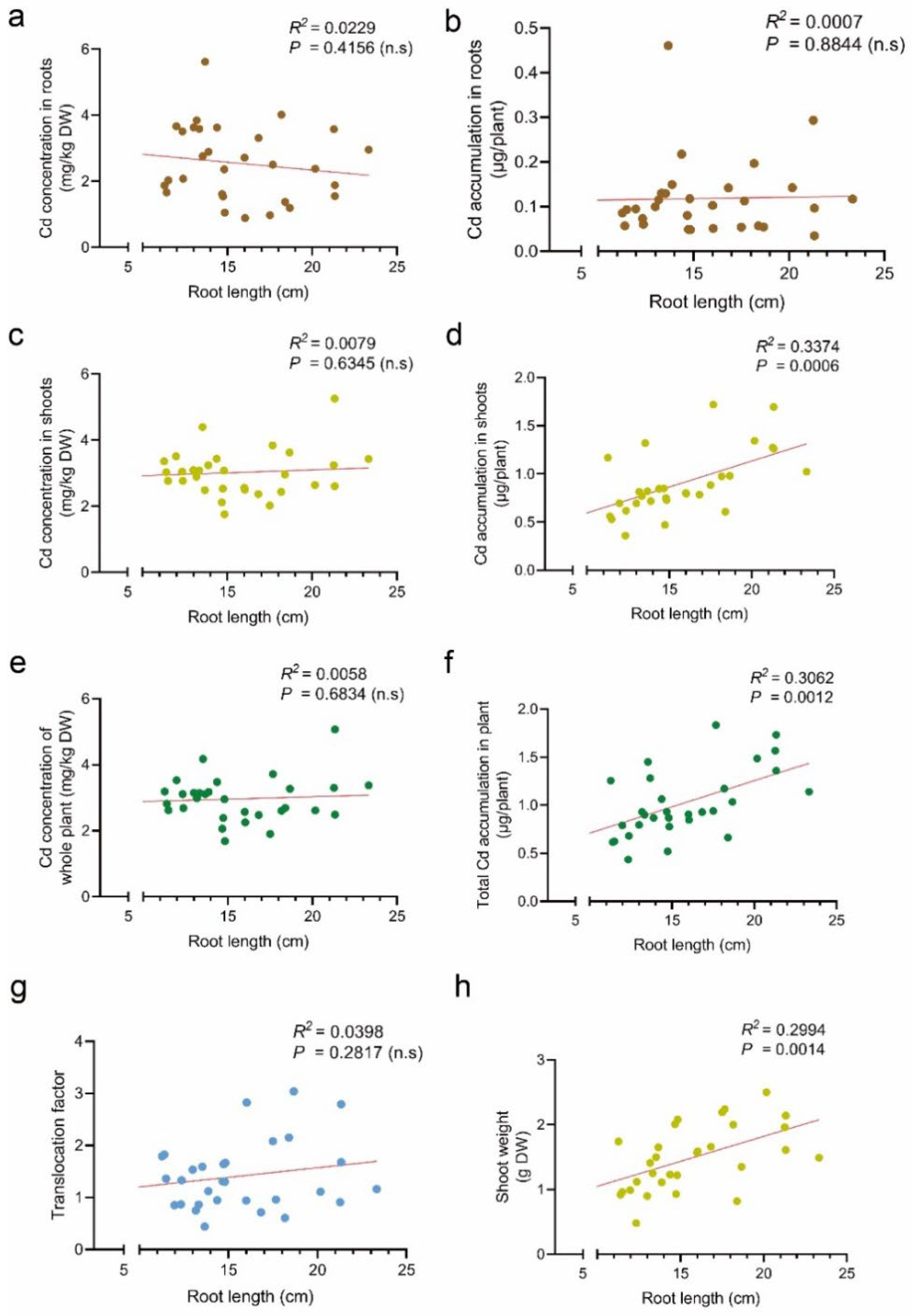
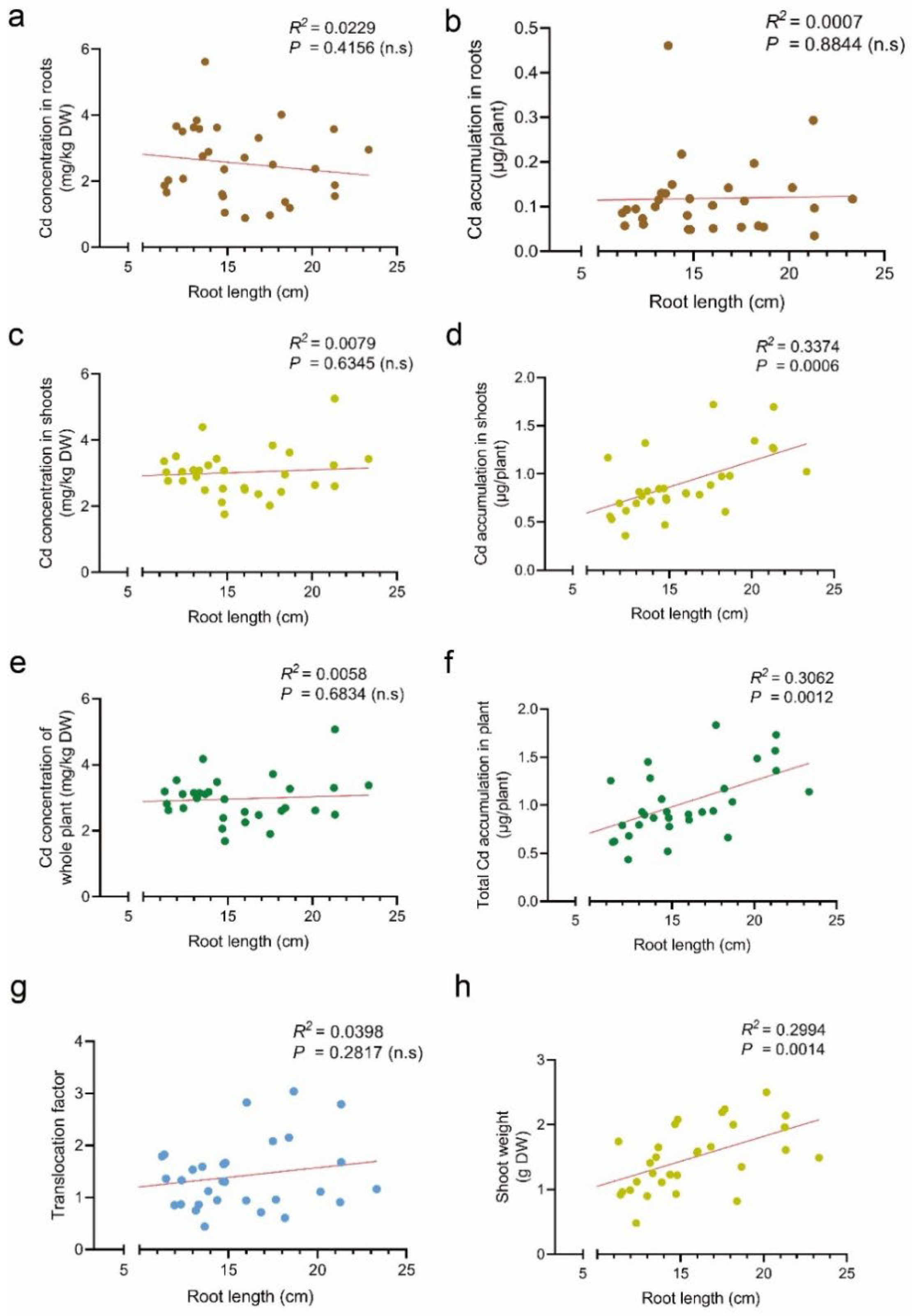
3.2. Effect of Root Length on Cd Accumulation in Tomato
3.3. Effect of Plant Height on Cd Accumulation in Tomato
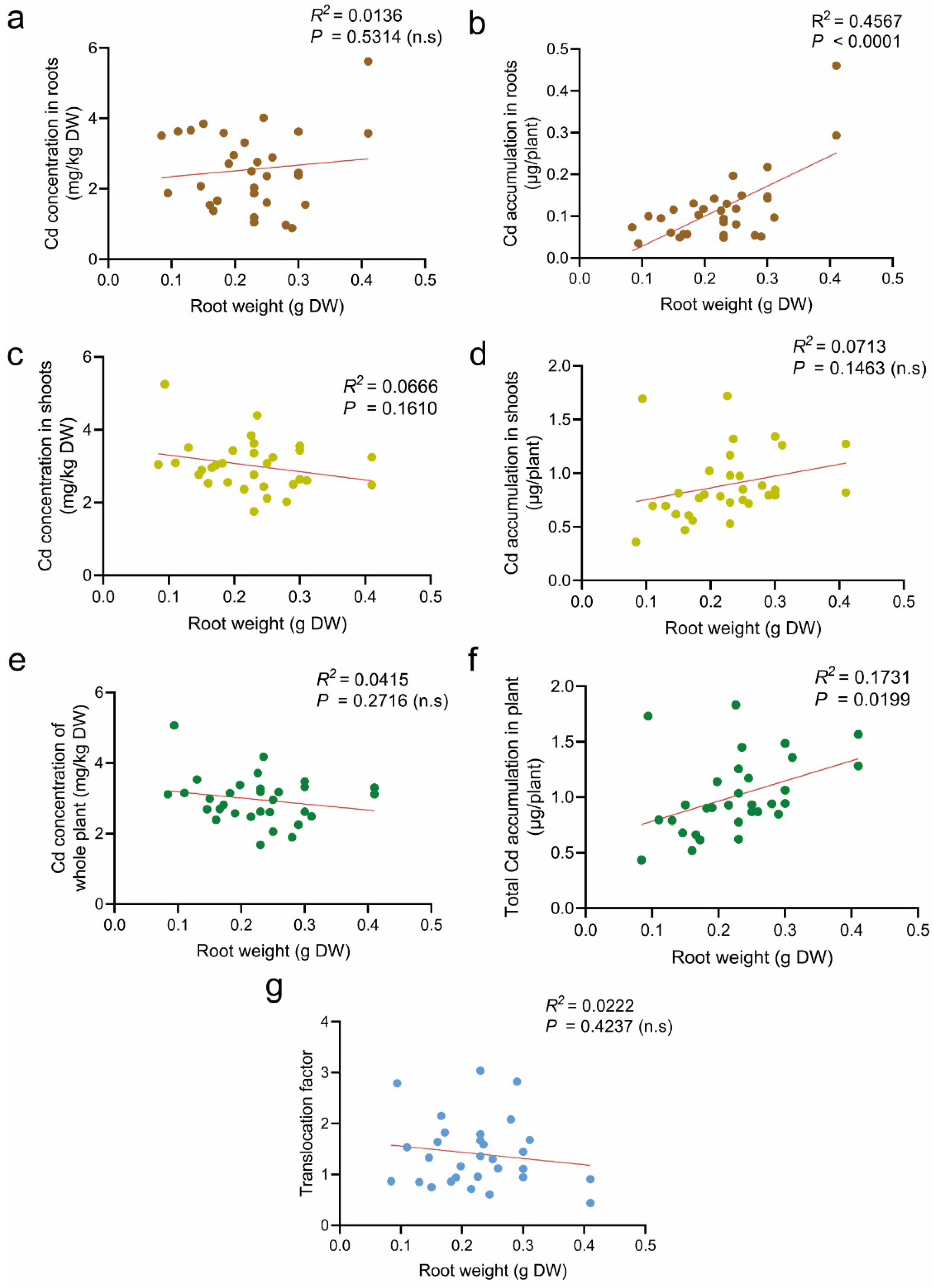
3.4. Effect of Root Weight on Cd Accumulation in Tomato
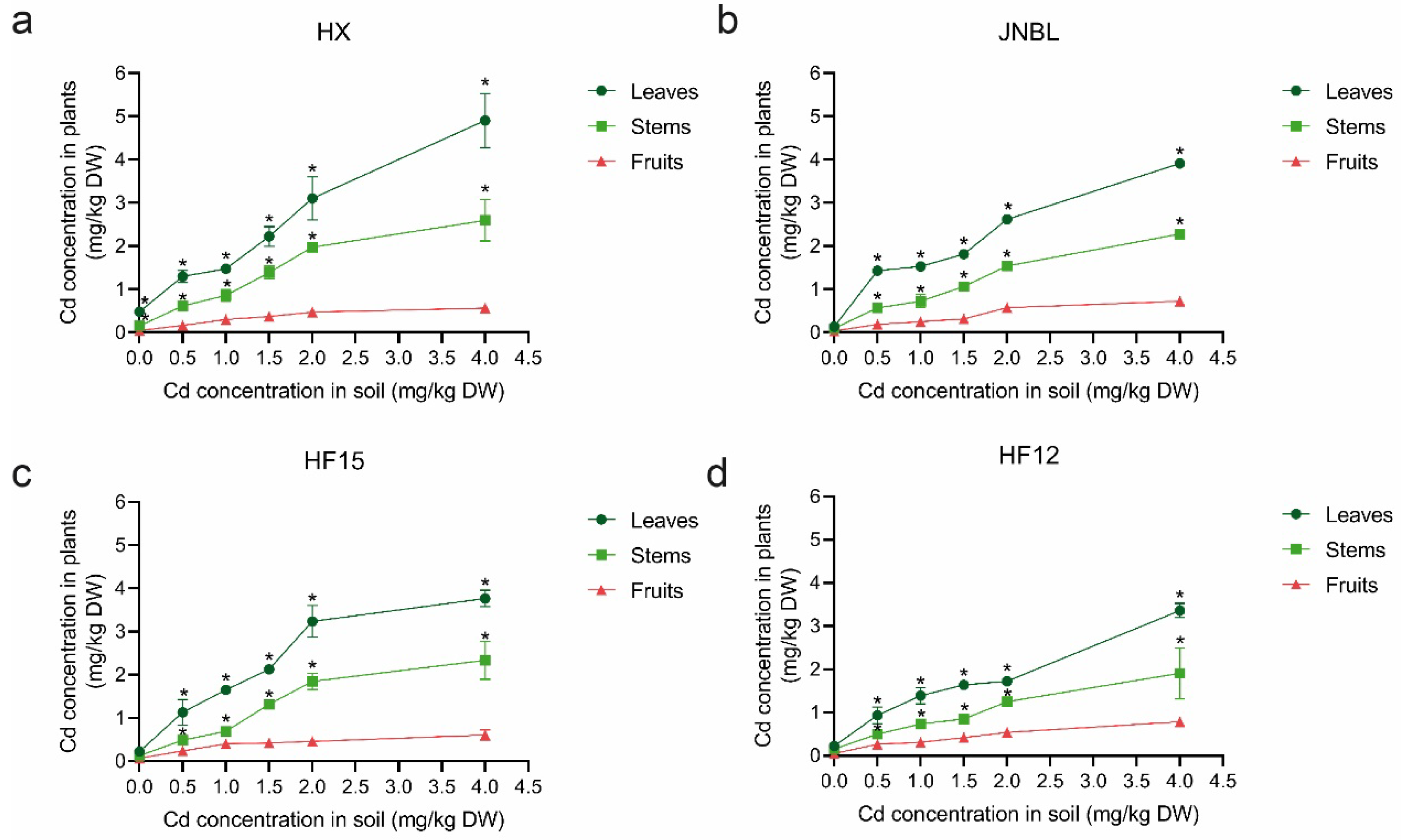
3.5. Differences of Cd Accumulation in Tomato Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Briffa, J.; Sinagra, E.; Blundell, R. Heavy metal pollution in the environment and their toxicological effects on humans. Heliyon 2020, 6, 26. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.M.; Wang, X.Y.; Liu, X.P. Detection of heavy metal ions by ratiometric photoelectric sensor. J. Agric. Food Chem. 2022, 70, 11468–11480. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Castro-Guerrero, N.; Mendoza-Cozatl, D.G. Moving toward a precise nutrition: Preferential loading of seeds with essential nutrients over non-essential toxic elements. Front. Plant Sci. 2014, 5, 7. [Google Scholar] [CrossRef]
- Koleli, N.; Eker, S.; Cakmak, I. Effect of zinc fertilization on cadmium toxicity in durum and bread wheat grown in zinc-deficient soil. Environ. Pollut. 2004, 131, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, J.C.; Yan, C.L.; Du, D.L.; Lu, H.L. The alleviation effect of iron on cadmium phytotoxicity in mangrove A. marina. Alleviation effect of iron on cadmium phytotoxicity in for mangrove Avicennia marina (Forsk.) Vierh. Chemosphere 2019, 226, 413–420. [Google Scholar]
- Topperwien, S.; Behra, R.; Sigg, L. Competition among zinc, manganese, and cadmium uptake in the freshwater alga Scenedesmus vacuolatus. Environ. Toxicol. Chem. 2007, 26, 483–490. [Google Scholar] [CrossRef]
- Pozgajova, M.; Navratilova, A.; Kovar, M. Curative potential of substances with bioactive properties to alleviate cd toxicity: A review. Int. J. Environ. Res. Public Health. 2022, 19, 42. [Google Scholar] [CrossRef]
- Liu, J.; Su, J.Y.; Wang, J.; Song, X.; Wang, H.W. A case study: Arsenic, cadmium and copper distribution in the soil-rice system in two main rice-producing provinces in China. Sustainability 2022, 14, 14355. [Google Scholar] [CrossRef]
- The Ministry of Environmental Protection. The Ministry of Land and Resources Report on the National Soil Contamination Survey; The Ministry of Environmental Protection: Beijing, China, 2014.
- Liu, W.T.; Zhou, Q.X.; Sun, Y.B.; Liu, R. Identification of Chinese cabbage genotypes with low cadmium accumulation for food safety. Environ. Pollut. 2009, 157, 1961–1967. [Google Scholar] [CrossRef]
- Selvam, A.; Wong, J.W.C. Cadmium uptake potential of Brassica napus cocropped with Brassica parachinensis and Zea mays. J. Hazard. Mater. 2009, 167, 170–178. [Google Scholar] [CrossRef]
- Delperee, C.; Lutts, S. Growth inhibition occurs independently of cell mortality in tomato (Solanum lycopersicum) exposed to high cadmium concentrations. J. Integr. Plant Biol. 2008, 50, 300–310. [Google Scholar] [CrossRef]
- Chen, X.M.; Shi, X.Y.; Ai, Q.; Han, J.Y.; Wang, H.S.; Fu, Q.S. Transcriptomic and metabolomic analyses reveal that exogenous strigolactones alleviate the response of melon root to cadmium stress. Hortic. Plant J. 2022, 8, 637–649. [Google Scholar] [CrossRef]
- Fusconi, A.; Repetto, O.; Bona, E.; Massa, N.; Gallo, C.; Dumas-Gaudot, E.; Berta, G. Effects of cadmium on meristem activity and nucleus ploidy in roots of Pisum sativum L. cv. Frisson seedlings. Environ. Exp. Bot. 2006, 58, 253–260. [Google Scholar] [CrossRef]
- Kubo, K.; Watanabe, Y.; Matsunaka, H.; Seki, M.; Fujita, M.; Kawada, N.; Hatta, K.; Nakajima, T. Differences in cadmium accumulation and root morphology in seedlings of japanese wheat varieties with distinctive grain cadmium concentration. Plant Prod. Sci. 2011, 14, 148–155. [Google Scholar] [CrossRef]
- Liu, Z.L.; Chen, M.D.; Lin, M.S.; Chen, Q.L.; Lu, Q.X.; Yao, J.; He, X.Y. Cadmium uptake and growth responses of seven urban flowering plants: Hyperaccumulator or bioindicator? Sustainability 2022, 14, 12. [Google Scholar] [CrossRef]
- Wu, M.X.; Luo, Q.; Liu, S.L.; Zhao, Y.; Long, Y.; Pan, Y.Z. Screening ornamental plants to identify potential Cd hyperaccumulators for bioremediation. Ecotoxicol. Environ. Saf. 2018, 162, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Bartoli, F.; Coinchelin, D.; Robin, C.; Echevarria, G. Impact of active transport and transpiration on nickel and cadmium accumulation in the leaves of the Ni-hyperaccumulator Leptoplax emarginata: A biophysical approach. Plant Soil 2012, 350, 99–115. [Google Scholar] [CrossRef]
- Van der Vliet, L.; Peterson, C.; Hale, B. Cd accumulation in roots and shoots of durum wheat: The roles of transpiration rate and apoplastic bypass. J. Exp. Bot. 2007, 58, 2939–2947. [Google Scholar] [CrossRef]
- Quinet, M.; Angosto, T.; Yuste-Lisbona, F.J.; Blanchard-Gros, R.; Bigot, S.; Martinez, J.P.; Lutts, S. Tomato fruit development and metabolism. Front. Plant Sci. 2019, 10, 23. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, X.C.; Jiang, J.B.; Zhao, T.T.; Xu, X.Y.; Yang, H.H.; Li, J.F. Virus-induced gene silencing of SlPYL4 decreases the drought tolerance of tomato. Hortic. Plant J. 2022, 8, 361–368. [Google Scholar] [CrossRef]
- Tomato Genome Consortium. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar] [CrossRef]
- Zhou, Z.; Yuan, Y.Q.; Wang, K.T.; Wang, H.J.; Huang, J.Q.; Yu, H.; Cui, X. Rootstock-scion interactions affect fruit flavor in grafted tomato. Hortic. Plant J. 2022, 8, 499–510. [Google Scholar] [CrossRef]
- Zhao, T.T.; Pei, T.; Jiang, J.B.; Yang, H.H.; Zhang, H.; Li, J.F.; Xu, X.Y. Understanding the mechanisms of resistance to tomato leaf mold: A review. Hortic. Plant J. 2022, 8, 667–675. [Google Scholar] [CrossRef]
- An, L.Y.; Pan, Y.H.; Wang, Z.B.; Zhu, C. Heavy metal absorption status of five plant species in monoculture and intercropping. Plant Soil 2011, 345, 237–245. [Google Scholar] [CrossRef]
- Hu, X.T.; Li, T.; Xu, W.H.; Chai, Y.R. Distribution of cadmium in subcellular fraction and expression difference of its transport genes among three cultivars of pepper. Ecotoxicol. Environ. Saf. 2021, 216, 10. [Google Scholar] [CrossRef]
- Wang, Q.; Huang, D.; Niu, D.S.; Deng, J.; Ma, F.W.; Liu, C.H. Overexpression of auxin response gene MdIAA24 enhanced cadmium tolerance in apple (Malus domestica). Ecotoxicol. Environ. Saf. 2021, 225, 8. [Google Scholar] [CrossRef]
- Ministry of Ecology and Environment of the People’s Republic of China. Soil Environmental Quality—Risk Control Standard for Soil Contamination of Agricultural Land; Ministry of Ecology and Environment of the People’s Republic of China: Beijing, China, 2018.
- Zhang, X.F.; Xia, H.P.; Li, Z.A.; Zhuang, P.; Gao, B. Potential of four forage grasses in remediation of Cd and Zn contaminated soils. Bioresour. Technol. 2010, 101, 2063–2066. [Google Scholar] [CrossRef]
- Rastmanesh, F.; Moore, F.; Keshavarzi, B. Speciation and phytoavailability of heavy metals in contaminated soils in sarcheshmeh area, Kerman Province, Iran. Bull. Environ. Contam. Toxicol. 2010, 85, 515–519. [Google Scholar] [CrossRef]
- Kubo, K.; Kobayashi, H.; Fujita, M.; Ota, T.; Minamiyama, Y.; Watanabe, Y.; Nakajima, T.; Shinano, T. Varietal differences in the absorption and partitioning of cadmium in common wheat (Triticum aestivum L.). Environ. Exp. Bot. 2016, 124, 79–88. [Google Scholar] [CrossRef]
- Szwalec, A.; Mundala, P.; Kedzior, R. Suitability of selected plant species for phytoremediation: A case study of a coal combustion ash landfill. Sustainability 2022, 14, 15. [Google Scholar] [CrossRef]
- Waheed, S.; Ahmad, R.; Irshad, M.; Khan, S.A.; Mahmood, Q.; Shahzad, M. Ca2SiO4 chemigation reduces cadmium localization in the subcellular leaf fractions of spinach (Spinacia oleracea L.) under cadmium stress. Ecotoxicol. Environ. Saf. 2021, 207, 10. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.T.; Wu, M.Y.; Yu, F.; Song, Q.; Zhao, Z.H.; Liao, L.; Tong, J.L. Enhanced cadmium phytoremediation capacity of poplar is associated with increased biomass and Cd accumulation under nitrogen deposition conditions. Ecotoxicol. Environ. Saf. 2022, 246, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.N.; Sa, G.; Zhang, Y.; Hou, S.Y.; Wu, X.; Zhao, N.; Zhang, Y.H.; Deng, S.R.; Deng, C.; Deng, J.Y.; et al. Populus euphratica annexin1 facilitates cadmium enrichment in transgenic Arabidopsis. J. Hazard. Mater. 2021, 405, 12. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.G.; Xia, S.L.; Liu, C.F.; Zhang, Z.; Shi, G.R. Variations in root morphology among 18 herbaceous species and their relationship with cadmium accumulation. Environ. Sci. Pollut. Res. 2017, 24, 4731–4740. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Z.; Zhou, H.; Shao, L.L.; Wang, H.R.; Zhang, Y.B.; Zhu, T.; Ma, L.T.; Ding, Q.; Ma, L.J. Root characteristics critical for cadmium tolerance and reduced accumulation in wheat (Triticum aestivum L.). J. Environ. Manag. 2022, 305, 11. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.Y.; Hu, C.M.; Jia, X.W.; Ren, Y.F.; Su, D.M.; He, J.Y. Physiological and biochemical bases of spermidine-induced alleviation of cadmium and lead combined stress in rice. Plant Physiol. Biochem. 2022, 189, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.D.; Yang, H.L.; Guo, W.L.; Li, X.Z.; Chen, B.H. Defense response of pumpkin rootstock to cadmium. Sci. Hortic. 2023, 308, 11. [Google Scholar] [CrossRef]
- Wang, F.J.; Tan, H.F.; Huang, L.H.; Cai, C.; Ding, Y.F.; Bao, H.; Chen, Z.X.; Zhu, C. Application of exogenous salicylic acid reduces Cd toxicity and Cd accumulation in rice. Ecotoxicol. Environ. Saf. 2021, 207, 9. [Google Scholar] [CrossRef]
- Wu, F.B.; Dong, J.; Cai, Y.; Chen, F.; Zhang, G.P. Differences in Mn uptake and subcellular distribution in different barley genotypes as a response to Cd toxicity. Sci. Total Environ. 2007, 385, 228–234. [Google Scholar] [CrossRef]
- Zhang, D.W.; Dong, F.; Zhang, Y.; Huang, Y.L.; Zhang, C.F. Mechanisms of low cadmium accumulation in storage root of sweetpotato (Ipomoea batatas L.). J. Plant Physiol. 2020, 254, 8. [Google Scholar] [CrossRef]
- Zhang, T.Y.; Wang, Y.; Munir, S.; Wang, T.T.; Ye, Z.B.; Zhang, J.H.; Zhang, Y.Y. Cyclin gene SlCycB1;2 alters plant architecture in association with histone H3.2 in tomato. Hortic. Plant J. 2022, 8, 341–350. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Zhang, C.; Zhang, Y. Tomato Accumulates Cadmium to a Concentration Independent of Plant Growth. Horticulturae 2023, 9, 1343. https://doi.org/10.3390/horticulturae9121343
Zhang X, Zhang C, Zhang Y. Tomato Accumulates Cadmium to a Concentration Independent of Plant Growth. Horticulturae. 2023; 9(12):1343. https://doi.org/10.3390/horticulturae9121343
Chicago/Turabian StyleZhang, Xingyu, Cong Zhang, and Yuyang Zhang. 2023. "Tomato Accumulates Cadmium to a Concentration Independent of Plant Growth" Horticulturae 9, no. 12: 1343. https://doi.org/10.3390/horticulturae9121343
APA StyleZhang, X., Zhang, C., & Zhang, Y. (2023). Tomato Accumulates Cadmium to a Concentration Independent of Plant Growth. Horticulturae, 9(12), 1343. https://doi.org/10.3390/horticulturae9121343