


Preliminary Analysis, Combined with Omics of Chilling Injury Mechanism of Peach Fruits with Different Cold Sensitivities during Postharvest Cold Storage
 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Determination of Ethylene and ABA Content and Firmness
2.3. Lipidomics Analysis
2.4. Transcriptome Sequencing
2.5. Analysis of Differential Genes by WGCNA
2.6. Prediction of Transcription Factors and Enrichment Analysis of Cis Motifs
2.7. Data Analysis
3. Results
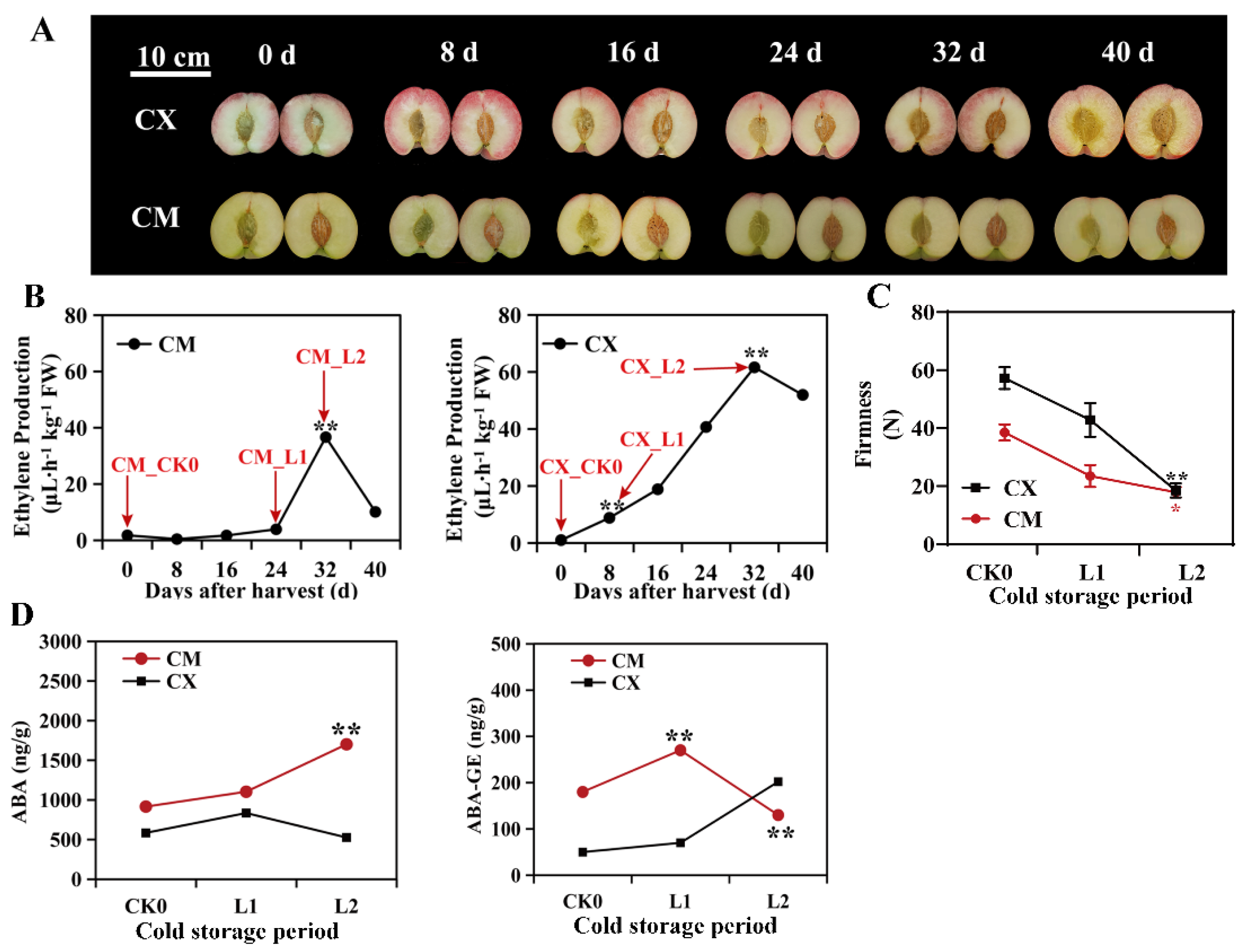
3.1. Changes in Chilling Injury Symptoms and the Ethylene Contents of Peaches with Different Cold Sensitivities during Low-Temperature Storage
3.2. Changes in ABA Contents and Fruit Firmness in Peaches with Different Cold Sensitivities during Low-Temperature Storage
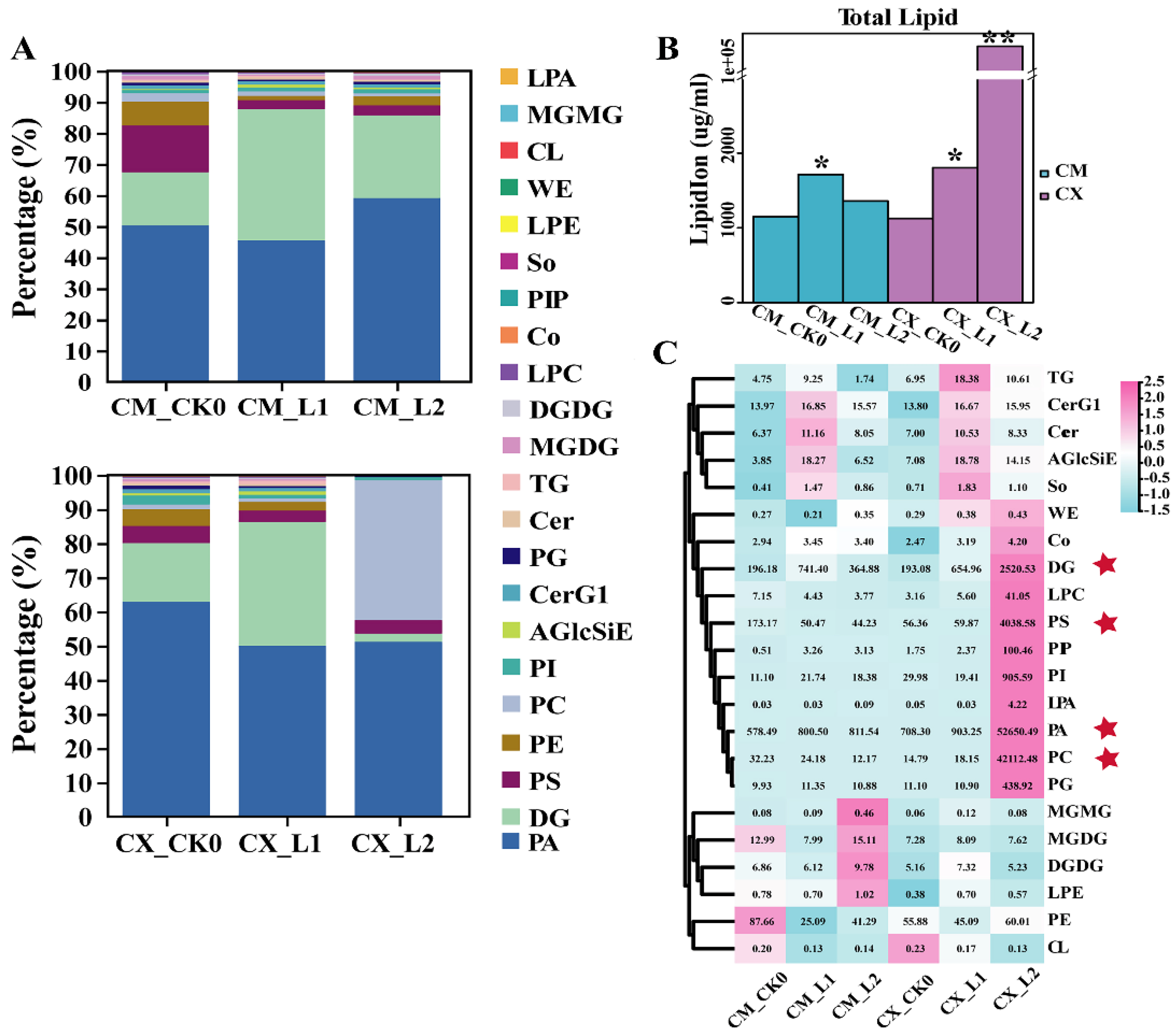
3.3. Changes in Lipid Compositions and Contents of Peaches with Different Cold Sensitivities during Low-Temperature Storage
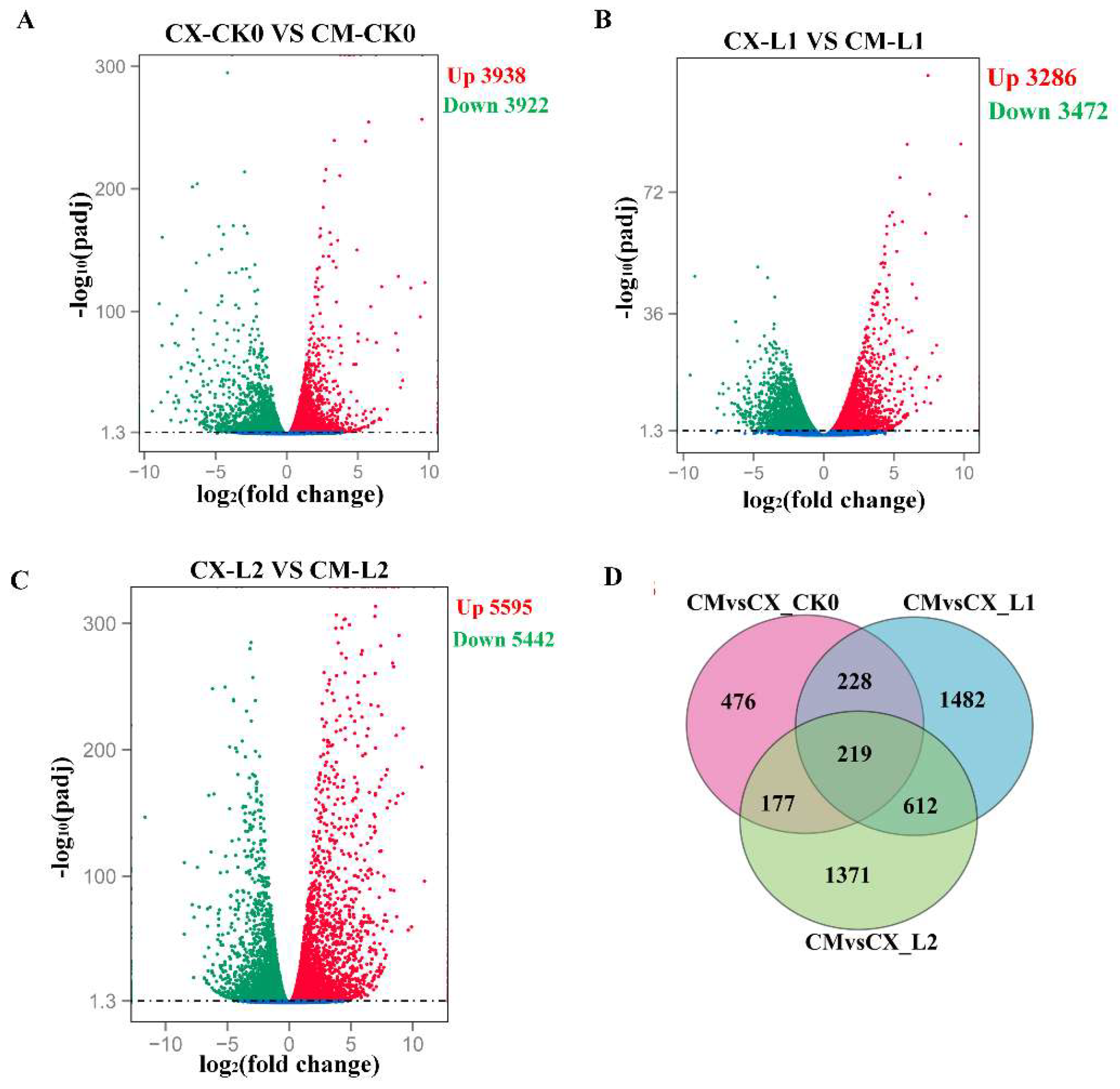
3.4. Transcriptional Analysis of Differentially Expressed Genes
3.5. Analysis of Differential Expression of Genes Related to Ethylene Biosynthesis and Signal Transduction in Peach Fruits during Low-Temperature Storage
3.6. Differential Expression Analysis of Genes Related to ABA Metabolism and Signal Transduction during Low-Temperature Storage
3.7. Differential Expression Analysis of Genes Related to Lipid Metabolism in Peach Fruits during Low-Temperature Storage
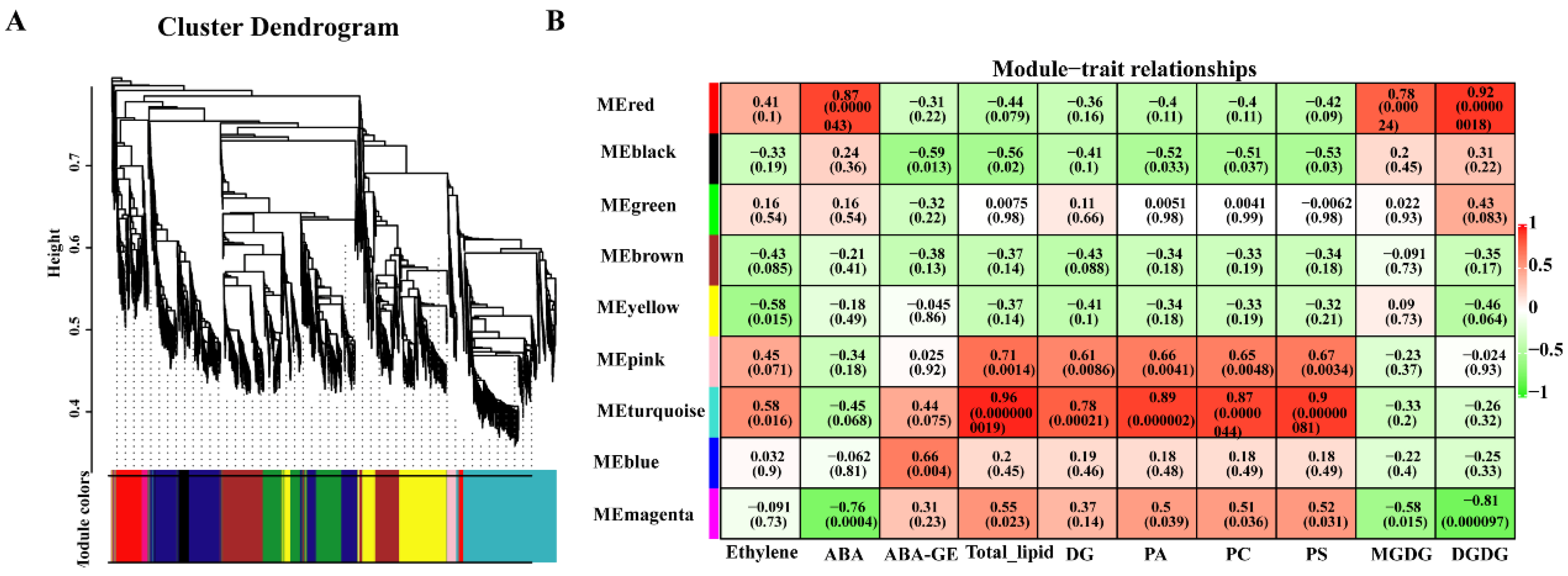
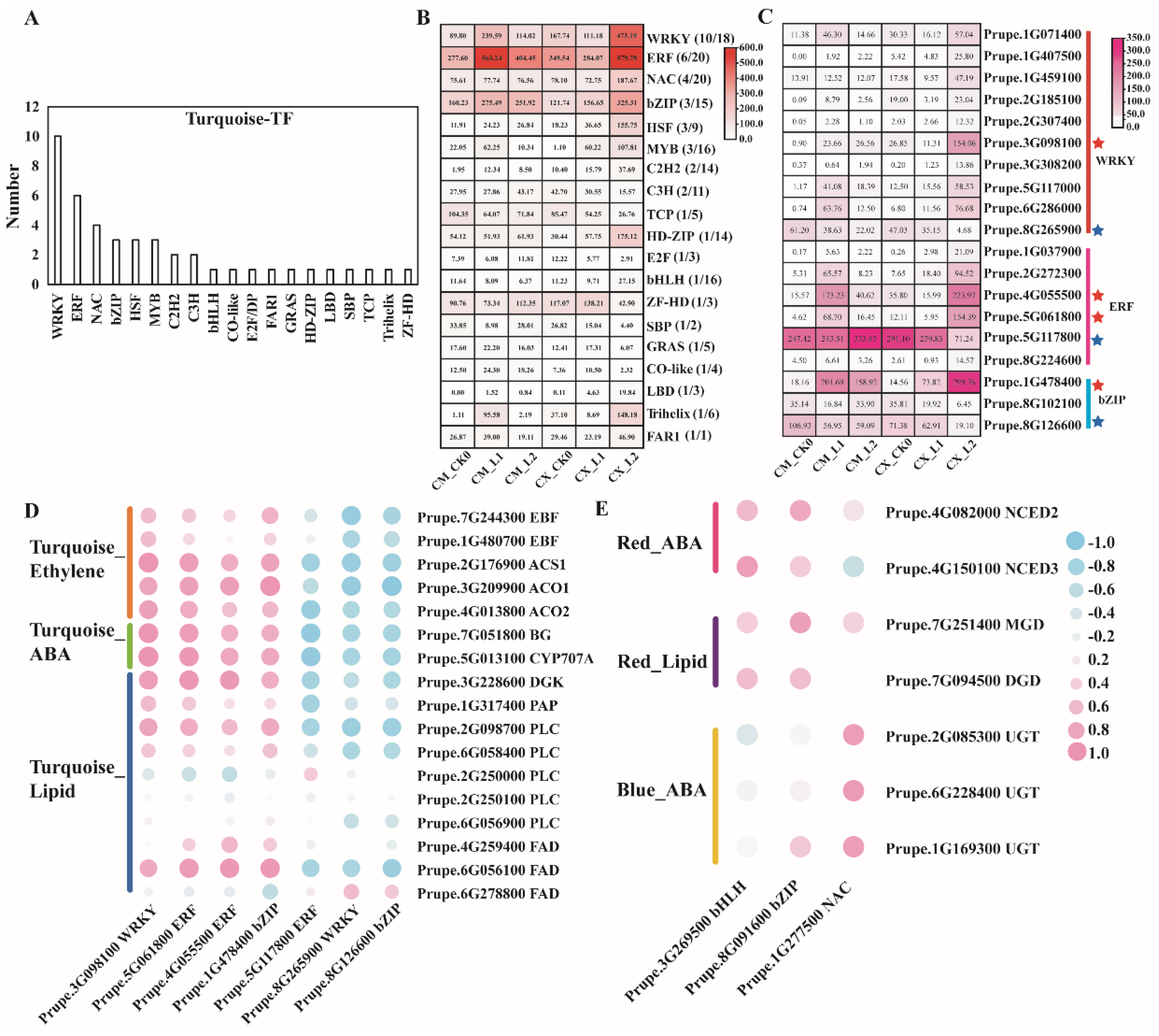
3.8. Analysis of Genes Related to Different Chilling Sensitivity Traits of Peach Fruits Based on WGCNA
3.9. Analysis of Cis-Acting Elements of Key Structural Gene Promoters in Different Cold-Sensitive Peaches during Low-Temperature Storage
4. Discussion
4.1. The Levels of Endogenous Hormones in Peach Fruits Change with the Occurrence of Chilling Injury Symptoms
4.2. The Changes in the Lipid Levels of Peach Fruits with Different Cold Sensitivities Were Different during Low-Temperature Storage
4.3. Transcription Factors Play a Key Regulatory Role in Low-Temperature Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Seymour, G.B.; Ostergaard, L.; Chapman, N.H.; Knapp, S.; Martin, C. Fruit development and ripening. Annu. Rev. Plant Biol. 2013, 64, 219–241. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, K.; Wu, C.; Zhao, Y.; Yin, X.; Zhang, B.; Grierson, D.; Chen, K.; Xu, C. Effect of ethylene on cell wall and lipid metabolism during alleviation of postharvest chilling injury in peach. Cells 2019, 8, 1612. [Google Scholar] [CrossRef] [PubMed]
- Brummell, D.A.; Dal Cin, V.; Crisosto, C.H.; Labavitch, J.M. Cell wall metabolism during maturation, ripening and senescence of peach fruit. J. Exp. Bot. 2004, 55, 2029–2039. [Google Scholar] [CrossRef] [PubMed]
- Ketsa, S.; Chidtragool, S.; Lurie, S. Prestorage heat treatment and poststorage quality of mango fruit. HortScience 2000, 35, 247–249. [Google Scholar] [CrossRef]
- Zhang, C.; Ding, Z.; Xu, X.; Wang, Q.; Qin, G.; Tian, S. Crucial roles of membrane stability and its related proteins in the tolerance of peach fruit to chilling injury. Amino Acids 2010, 39, 181–194. [Google Scholar] [CrossRef]
- Cabrera, R.M.; Saltveit, M.E. Physiological Response to Chilling Temperatures of Intermittently Warmed Cucumber Fruit. J. Am. Soc. Hortic. Sci. 2019, 115, 256–261. [Google Scholar] [CrossRef]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.Y.; Li, J.; Wang, P.Y.; Qin, F.; et al. Plant abiotic stress response and nutrient use efficiency. Sci. China Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef]
- Tang, J.; Zhao, Y.; Qi, S.; Dai, Q.; Lin, Q.; Duan, Y. Abscisic acid alleviates chilling injury in cold-stored peach fruit by regulating ethylene and hydrogen peroxide metabolism. Front. Plant Sci. 2022, 13, 987573. [Google Scholar] [CrossRef]
- Deluc, L.G.; Quilici, D.R.; Decendit, A.; Grimplet, J.; Wheatley, M.D.; Schlauch, K.A.; Mérillon, J.M.; Cushman, J.C.; Cramer, G.R. Water deficit alters differentially metabolic pathways affecting important flavor and quality traits in grape berries of Cabernet Sauvignon and Chardonnay. BMC Genom. 2009, 10, 212. [Google Scholar] [CrossRef]
- Jin, M.; Jiao, J.; Zhao, Q.; Ban, Q.; Gao, M.; Suo, J.; Zhu, Q.; Rao, J. Dose effect of exogenous abscisic acid on controlling lignification of postharvest kiwifruit (Actinidia chinensis cv. hongyang). Food Control 2021, 124, 107911. [Google Scholar] [CrossRef]
- Chen, B.; Yang, H. 6-Benzylaminopurine alleviates chilling injury of postharvest cucumber fruit through modulating antioxidant system and energy status. J. Sci. Food Agric. 2013, 93, 1915–1921. [Google Scholar] [CrossRef] [PubMed]
- Luengwilai, K.; Beckles, D.M.; Saltveit, M.E. Chilling-injury of harvested tomato (Solanum lycopersicum L.) cv. Micro-Tom fruit is reduced by temperature pre-treatments. Postharvest Biol. Technol. 2012, 63, 123–128. [Google Scholar] [CrossRef]
- Cao, S.; Zheng, Y.; Wang, K.; Jin, P.; Rui, H. Methyl jasmonate reduces chilling injury and enhances antioxidant enzyme activity in postharvest loquat fruit. Food Chem. 2009, 115, 1458–1463. [Google Scholar] [CrossRef]
- Wang, K.; Yin, X.R.; Zhang, B.; Grierson, D.; Xu, C.J.; Chen, K.S. Transcriptomic and metabolic analyses provide new insights into chilling injury in peach fruit. Plant Cell Environ. 2017, 40, 1531–1551. [Google Scholar] [CrossRef] [PubMed]
- Guy, C.; Kaplan, F.; Kopka, J.; Selbig, J.; Hincha, D.K. Metabolomics of temperature stress. Physiol. Plant. 2008, 132, 220–235. [Google Scholar] [CrossRef] [PubMed]
- Funnekotter, B.; Kaczmarczyk, A.; Turner, S.R.; Bunn, E.; Zhou, W.; Smith, S.; Flematti, G.; Mancera, R.L. Acclimation-induced changes in cell membrane composition and influence on cryotolerance of in vitro shoots of native plant species. Plant Cell Tissue Organ Cult. 2013, 114, 83–96. [Google Scholar] [CrossRef]
- Yamaki, S.; Uritani, I. Mechanism of chilling injury in sweet potatoes part V biochemical mechanism of chilling injury with special reference to mitochondrial lipid components. Agric. Biol. Chem. 1972, 36, 47–55. [Google Scholar] [CrossRef]
- Thalhammer, A.; Bryant, G.; Sulpice, R.; Hincha, D.K. Disordered cold regulated15 proteins protect chloroplast membranes during freezing through binding and folding, But do not stabilize chloroplast enzymes in vivo. Plant Physiol. 2014, 166, 190–201. [Google Scholar] [CrossRef]
- Ma, Y.; Hu, S.; Chen, G.; Zheng, Y.; Jin, P. Cold shock treatment alleviates chilling injury in peach fruit by regulating antioxidant capacity and membrane lipid metabolism. Food Qual. Saf. 2022, 6, fyab026. [Google Scholar] [CrossRef]
- Franzoni, G.; Spadafora, N.D.; Sirangelo, T.M.; Ferrante, A.; Rogers, H.J. Biochemical and molecular changes in peach fruit exposed to cold stress conditions. Mol. Hortic. 2023, 3, 24. [Google Scholar] [CrossRef]
- Wang, X.; Ding, Y.; Li, Z.; Shi, Y.; Wang, J.; Hua, J.; Gong, Z.; Zhou, J.M.; Yang, S. PUB25 and PUB26 Promote Plant Freezing Tolerance by Degrading the Cold Signaling Negative Regulator MYB15. Dev. Cell 2019, 51, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Thomashow, M.F. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [PubMed]
- Duan, W.; Yang, C.; Cao, X.; Wei, C.; Chen, K.; Li, X.; Zhang, B. Chilling-induced peach flavor loss is associated with expression and DNA methylation of functional genes. J. Adv. Res. 2022, 53, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Lurie, S.; Crisosto, C.H. Chilling injury in peach and nectarine. Postharvest Biol. Technol. 2005, 37, 195–208. [Google Scholar] [CrossRef]
- Wang, C.Y. Changes of polyamines and ethylene in cucumber seedlings in response to chilling stress. Physiol. Plant. 1987, 69, 253–257. [Google Scholar] [CrossRef]
- Barroso, C.; Romero, L.C.; Cejudo, F.J.; Vega, J.M.; Gotor, C. Salt-specific regulation of the cytosolic O-acetylserine(thiol)lyase gene from Arabidopsis thaliana is dependent on abscisic acid. Plant Mol. Biol. 1999, 40, 729–736. [Google Scholar] [CrossRef]
- Zuo, X.; Cao, S.; Zhang, M.; Cheng, Z.; Cao, T.; Jin, P.; Zheng, Y. High relative humidity (HRH) storage alleviates chilling injury of zucchini fruit by promoting the accumulation of proline and ABA. Postharvest Biol. Technol. 2021, 171, 111344. [Google Scholar] [CrossRef]
- Yoshida, T.; Christmann, A.; Yamaguchi-Shinozaki, K.; Grill, E.; Fernie, A.R. Revisiting the Basal Role of ABA—Roles Outside of Stress. Trends Plant Sci. 2019, 24, 625–635. [Google Scholar] [CrossRef]
- Lyons, J.M. Chilling Injury in Plants. Annu. Rev. Plant Physiol. 1973, 24, 445–466. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Biomembranes—Part C: Biological Oxidations. Methods Enzymol. 1978, 52, 302–310. [Google Scholar]
- Tan, W.J.; Yang, Y.C.; Zhou, Y.; Huang, L.P.; Xu, L.; Chen, Q.F.; Yu, L.J.; Xiao, S. Diacylglycerol acyltransferase and diacylglycerol kinase modulate triacylglycerol and phosphatidic acid production in the plant response to freezing stress. Plant Physiol. 2018, 177, 1303–1318. [Google Scholar] [CrossRef] [PubMed]
- Moellering, E.R.; Benning, C. Galactoglycerolipid metabolism under stress: A time for remodeling. Trends Plant Sci. 2011, 16, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Wallace, P.A.; Teakle, N.L.; Colmer, T.D. Measuring Soluble Ion Concentrations (Na+, K+, Cl−) in Salt-Treated. In Plants BT—Plant Stress Tolerance: Methods and Protocols; Springer: New York, NY, USA, 2010. [Google Scholar] [CrossRef]
- Song, C.; Wang, K.; Xiao, X.; Liu, Q.; Yang, M.; Li, X.; Feng, Y.; Li, S.; Shi, L.; Chen, W.; et al. Membrane lipid metabolism influences chilling injury during cold storage of peach fruit. Food Res. Int. 2022, 157, 111249. [Google Scholar] [CrossRef] [PubMed]
- Cao, K.; Yang, X.; Li, Y.; Zhu, G.; Fang, W.; Chen, C.; Wang, X.; Wu, J.; Wang, L. New high-quality peach (Prunus persica L. Batsch) genome assembly to analyze the molecular evolutionary mechanism of volatile compounds in peach fruits. Plant J. 2021, 108, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Tweneboah, S.; Oh, S.K. Biological roles of NAC transcription factors in the regulation of biotic and abiotic stress responses in solanaceous crops. J. Plant Biotechnol. 2017, 44, 1–11. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, L.; Xia, C.; Zhao, G.; Liu, J.; Jia, J.; Kong, X. A novel wheat bZIP transcription factor, TabZIP60, confers multiple abiotic stress tolerances in transgenic Arabidopsis. Physiol. Plant. 2015, 153, 538–554. [Google Scholar] [CrossRef]
- Jiao, C. PpCBF6 Is Involved in Phytosulfokine α-Retarded Chilling Injury by Suppressing the Expression of PpLOX5 in Peach Fruit. Front. Plant Sci. 2022, 13, 874338. [Google Scholar] [CrossRef]
- Luo, D.L.; Ba, L.J.; Shan, W.; Kuang, J.F.; Lu, W.J.; Chen, J.Y. Involvement of WRKY Transcription Factors in Abscisic-Acid-Induced Cold Tolerance of Banana Fruit. J. Agric. Food Chem. 2017, 65, 3627–3635. [Google Scholar] [CrossRef]
- Zhang, S.; Cao, K.; Wei, Y.; Jiang, S.; Ye, J.; Xu, F.; Chen, Y.; Shao, X. PpBZR1, a BES/BZR transcription factor, enhances cold stress tolerance by suppressing sucrose degradation in peach fruit. Plant Physiol. Biochem. 2023, 202, 107972. [Google Scholar] [CrossRef]
- Zheng, Y.; Liu, Z.; Wang, H.; Zhang, W.; Li, S.; Xu, M. Transcriptome and genome analysis to identify C2H2 genes participating in low-temperature conditioning-alleviated postharvest chilling injury of peach fruit. Food Qual. Saf. 2022, 6, fyac059. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhan, W.; Wang, Y.; Duan, W.; Li, A.; Miao, Y.; Wang, H.; Meng, J.; Liu, H.; Niu, L.; Pan, L.; et al. Preliminary Analysis, Combined with Omics of Chilling Injury Mechanism of Peach Fruits with Different Cold Sensitivities during Postharvest Cold Storage. Horticulturae 2024, 10, 46. https://doi.org/10.3390/horticulturae10010046
Zhan W, Wang Y, Duan W, Li A, Miao Y, Wang H, Meng J, Liu H, Niu L, Pan L, et al. Preliminary Analysis, Combined with Omics of Chilling Injury Mechanism of Peach Fruits with Different Cold Sensitivities during Postharvest Cold Storage. Horticulturae. 2024; 10(1):46. https://doi.org/10.3390/horticulturae10010046
Chicago/Turabian StyleZhan, Wenduo, Yan Wang, Wenyi Duan, Ang Li, Yule Miao, Hongmei Wang, Junren Meng, Hui Liu, Liang Niu, Lei Pan, and et al. 2024. "Preliminary Analysis, Combined with Omics of Chilling Injury Mechanism of Peach Fruits with Different Cold Sensitivities during Postharvest Cold Storage" Horticulturae 10, no. 1: 46. https://doi.org/10.3390/horticulturae10010046
APA StyleZhan, W., Wang, Y., Duan, W., Li, A., Miao, Y., Wang, H., Meng, J., Liu, H., Niu, L., Pan, L., Sun, S., Cui, G., Wang, Z., & Zeng, W. (2024). Preliminary Analysis, Combined with Omics of Chilling Injury Mechanism of Peach Fruits with Different Cold Sensitivities during Postharvest Cold Storage. Horticulturae, 10(1), 46. https://doi.org/10.3390/horticulturae10010046