


Growth, Phenotypic Plasticity and Fruit Quality in Tomato: A Study under High Temperature and Elevated CO2

 ,
,  ,
,  , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Location, Plant Material and Crop Management
2.2. Growth Chambers and Treatments
2.3. Seedling Emergence and Growth Parameters
2.4. Phenotypic Plasticity
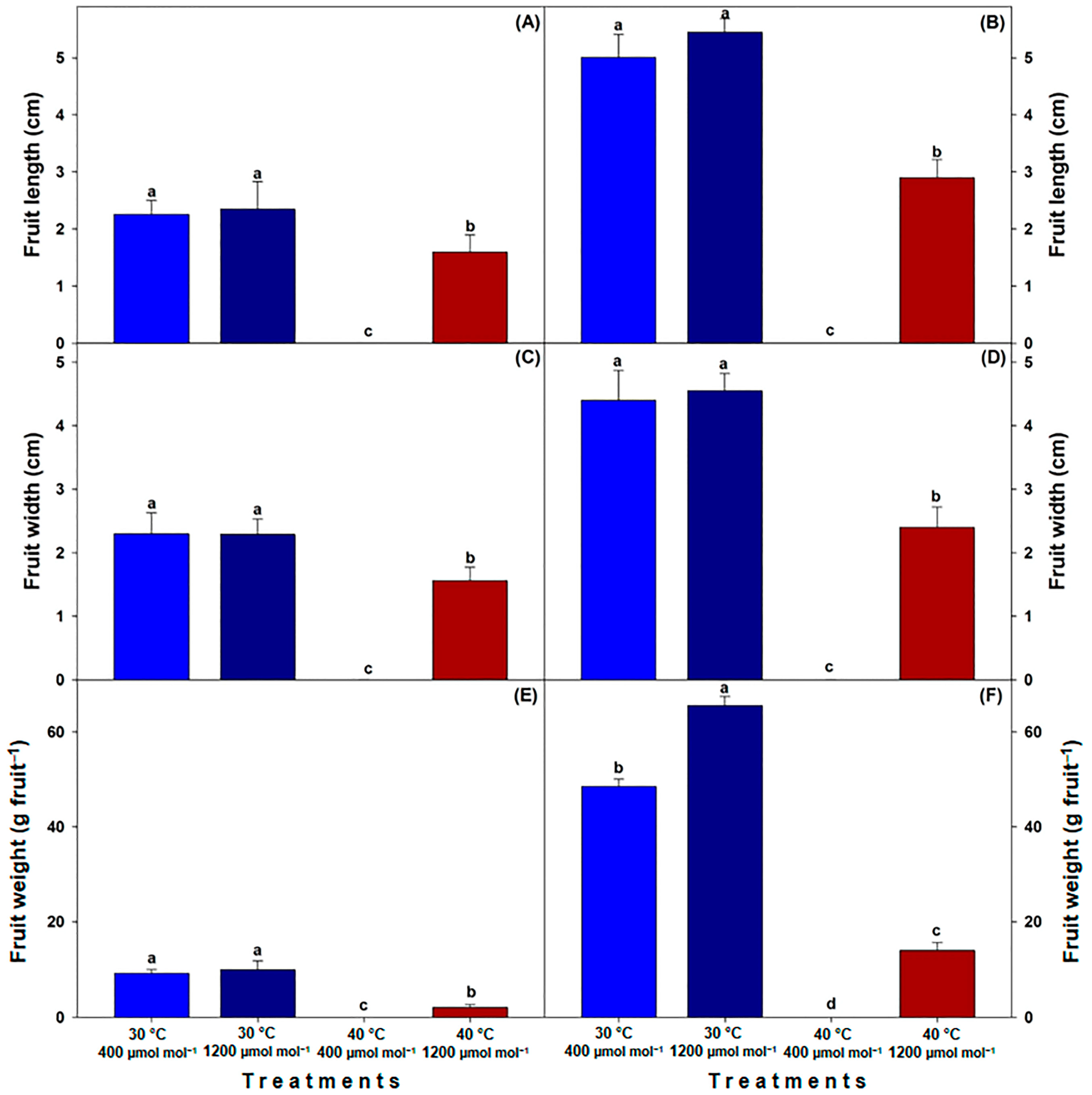
2.5. Fruit Traits
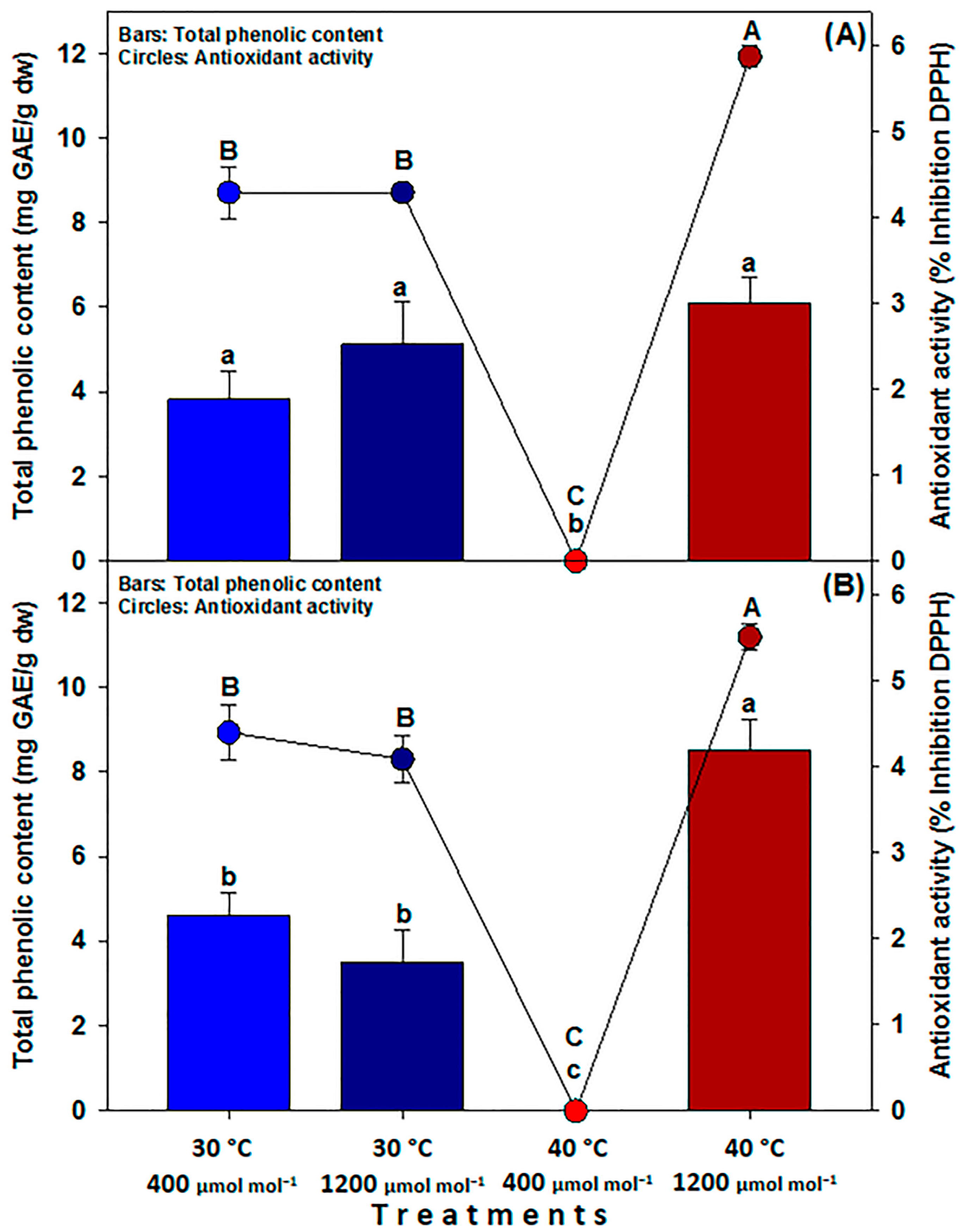
2.6. Extraction and Determination of Phenolic Compounds
2.7. Determination of Antioxidant Activity by DPPH
2.8. Experimental Design and Statistical Analyses
3. Results
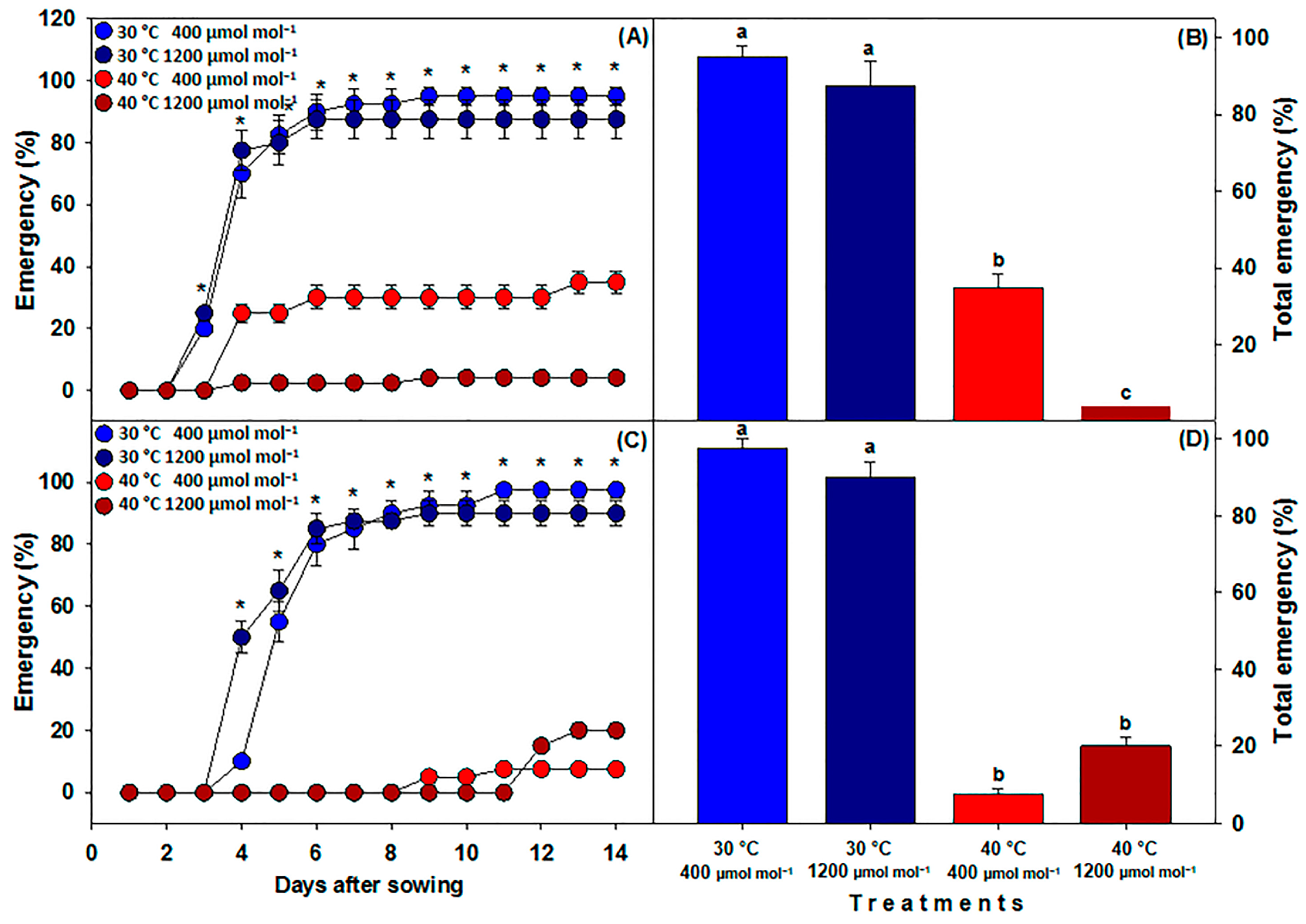
3.1. Seedling Emergence
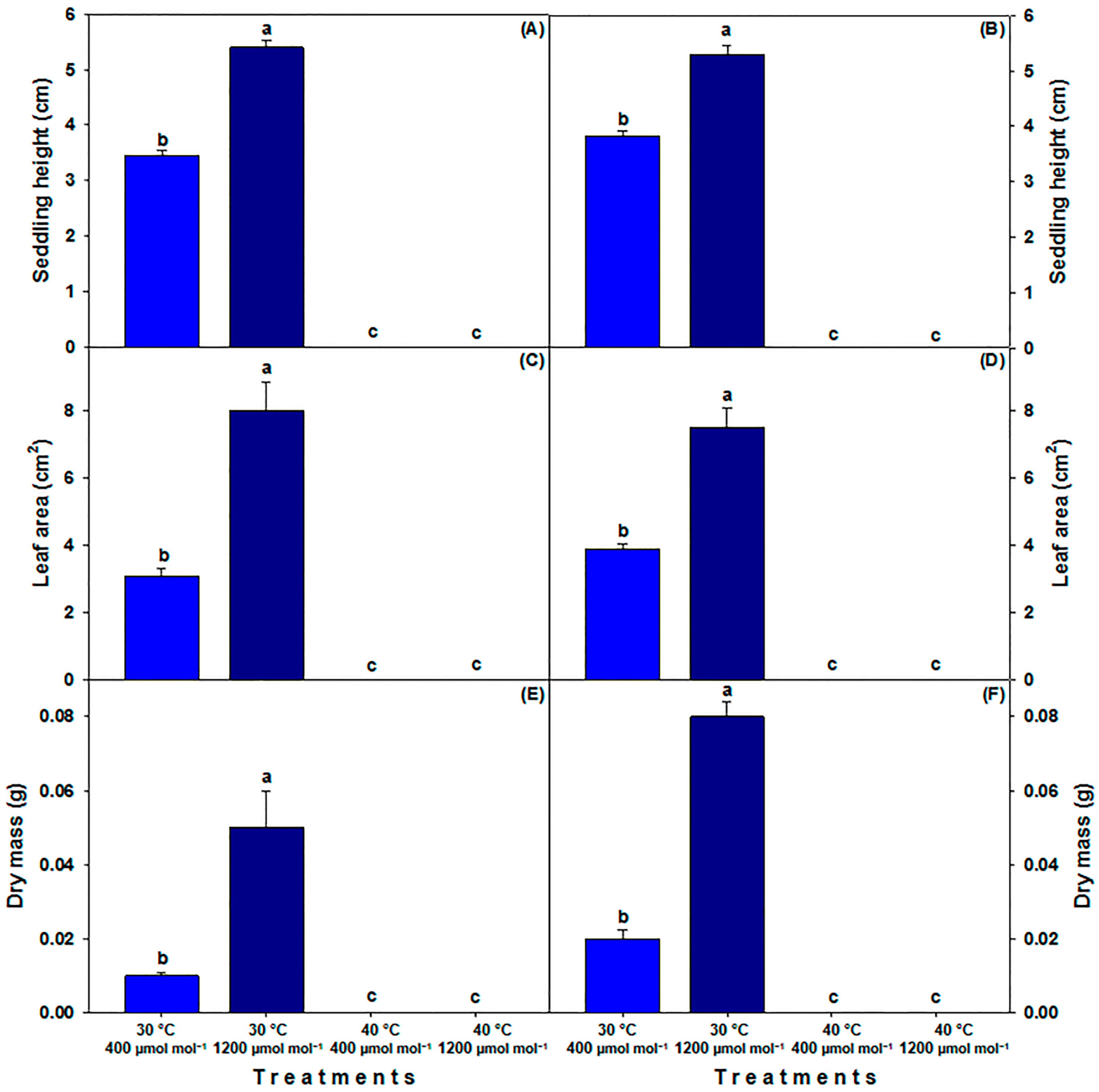
3.2. Growth Parameters in Seedling and Plant
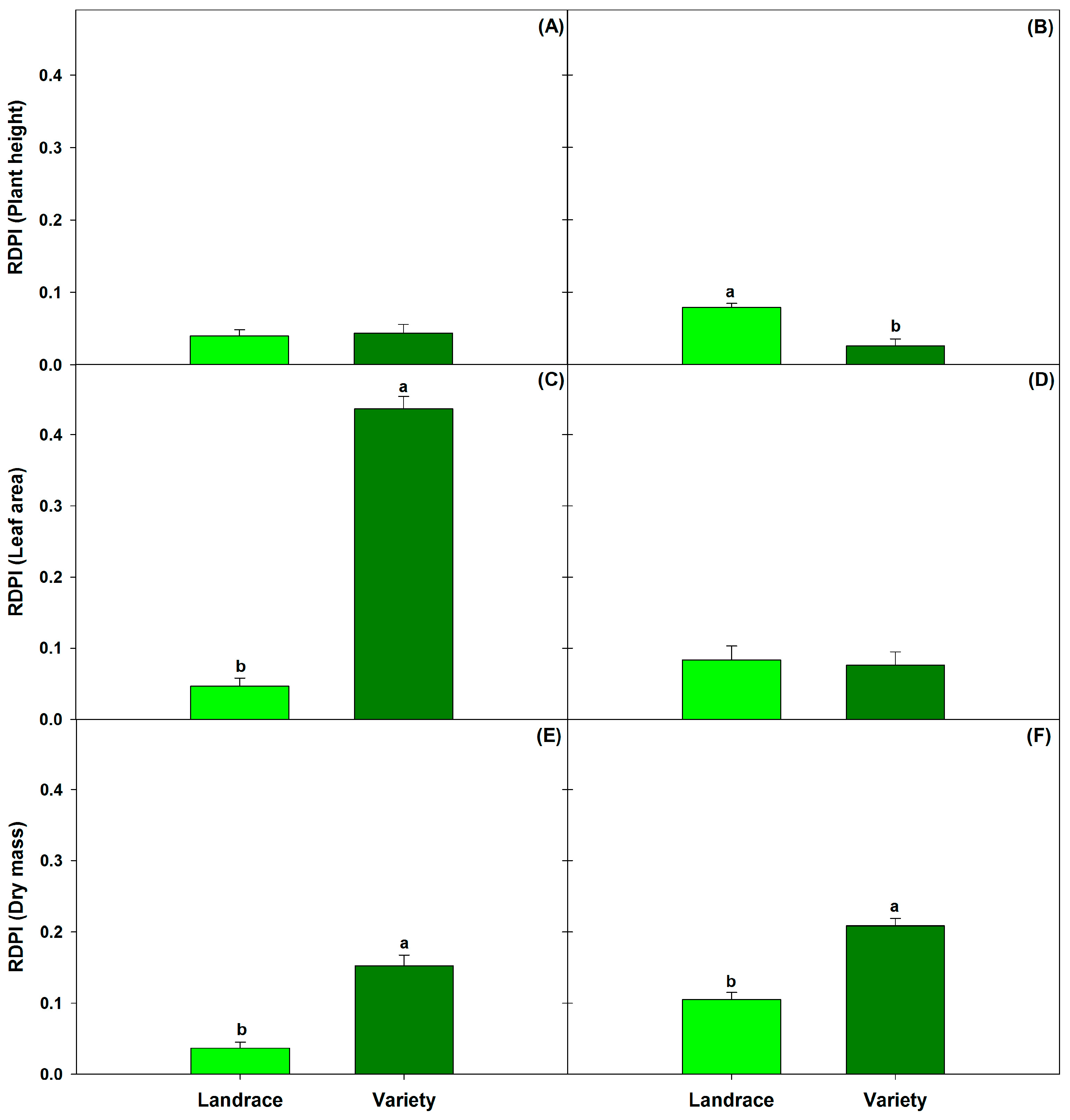
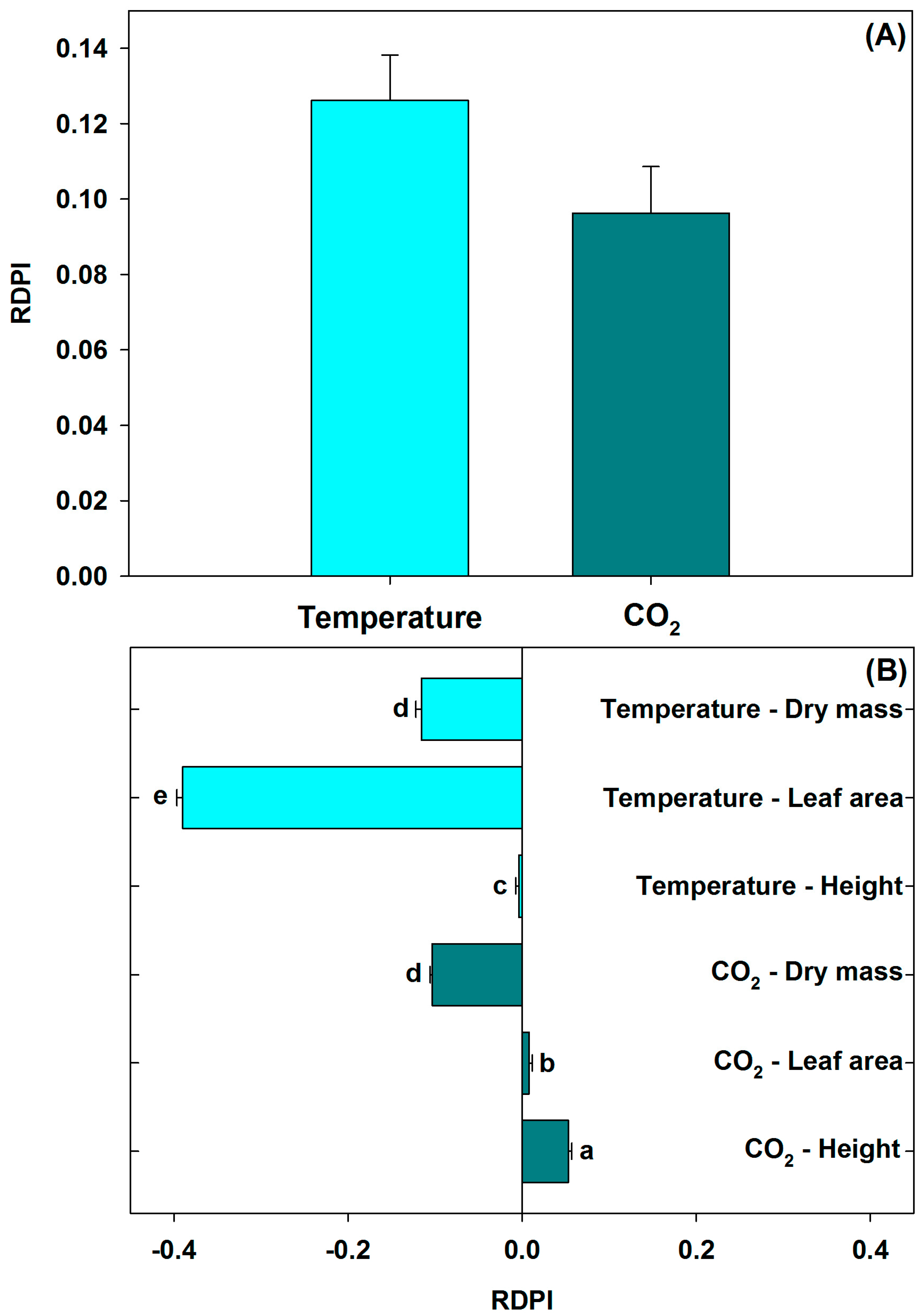
3.3. Relative Distance Plasticity Index (RDPI)
3.4. Fruit Quality
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, Z.; Shimizu, H.; Yagasaki, Y.; Ito, S.; Zheng, Y.; Zhou, G. Interactive effects of elevated CO2, drought, and warming on plants. J. Plant Growth Regul. 2013, 32, 692–707. [Google Scholar] [CrossRef]
- IPCC. Intergovernmental Panel on Climate Change. Available online: https://www.ipcc.ch/ (accessed on 12 May 2023).
- Garruña-Hernández, R.; Orellana, R.; Larque-Saavedra, A.; Canto, A. Understanding the physiological responses of a tropical crop (Capsicum Chinense Jacq.) at high temperature. PLoS ONE 2014, 9, e111402. [Google Scholar] [CrossRef]
- Pereyda-González, J.M.; De-la-Peña, C.; Tezara, W.; Zamora-Bustillos, R.; Andueza-Noh, R.H.; Noh-Kú, J.G.; Carrera-Marín, M.; Garruña, R. High temperature and elevated CO2 modify phenology and growth in pepper plants. Agronomy 2022, 12, 1836. [Google Scholar] [CrossRef]
- Jarvis, A.; Lane, A.; Hijmans, R.J. The effect of climate change on crop wild relatives. Agric. Ecosyst. Environ. 2008, 126, 13–23. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Garruña-Hernández, R.; Pereyda-González, J.; Oliva-Ruíz, M.; Castillo-Collí, M.; Ríos-Bolívar, F.; Cetina-Escalante, R. Hortalizas tropicales: Súper plantas ante el cambio climático. Bioagrociencias 2021, 14, 46–55. [Google Scholar] [CrossRef]
- O’Carrigan, A.; Hinde, E.; Lu, N.; Xu, X.Q.; Duan, H.; Huang, G.; Mak, M.; Bellotti, B.; Chen, Z.H. Effects of light irradiance on stomatal regulation and growth of tomato. Environ. Exp. Bot. 2014, 98, 65–73. [Google Scholar] [CrossRef]
- Desneux, N.; Luna, M.G.; Guillemaud, T.; Urbaneja, A. The invasive South American tomato pinworm, Tuta absoluta, continues to spread in Afro-Eurasia and beyond: The new threat to tomato world production. J. Pest. Sci. (2004) 2011, 84, 403–408. [Google Scholar] [CrossRef]
- Atif, M.J.; Jellani, G.; Malik, M.H.A.; Saleem, N.; Ullah, H.; Khan, M.Z.; Ikram, S. Different growth media effect the germination and growth of tomato seedlings. Sci. Technol. Dev. 2016, 35, 123–127. [Google Scholar] [CrossRef]
- van Dam, B.; Goffau, M.; van Lidt de Jeude, J.; Naika, S. Cultivation of Tomato: Production, Processing and Marketing, 4th ed.; Agromisa/CTA: Wageningen, The Netherlands, 2005; pp. 6–84. [Google Scholar]
- Li, X.; Ahammed, G.J.; Zhang, Y.Q.; Zhang, G.Q.; Sun, Z.H.; Zhou, J.; Zhou, Y.H.; Xia, X.J.; Yu, J.Q.; Shi, K. Carbon dioxide enrichment alleviates heat stress by improving cellular redox homeostasis through an ABA-independent process in tomato plants. Plant Biol. 2015, 17, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Valladares, F.; Sanchez-Gomez, D.; Zavala, M.A. Quantitative estimation of phenotypic plasticity: Bridging the gap between the evolutionary concept and its ecological applications. J. Ecol. 2006, 94, 1103–1116. [Google Scholar] [CrossRef]
- Hernández-Pinto, C.; Garruña, R.; Andueza-Noh, R.; Hernández-Núñez, E.; Zavala-León, M.J.; Pérez-Gutiérrez, A. Almacenamiento postcosecha de frutos: Alternativa para mejorar la calidad fisiológica de semillas de chile habanero. Biociencias 2020, 7, 1–15. [Google Scholar] [CrossRef]
- Valladares, F.; Chico, J.; Aranda, I.; Balaguer, L.; Dizengremel, P.; Manrique, E.; Dreyer, E. The greater seedling high-light tolerance of Quercus robur over Fagus sylvatica is linked to a greater physiological plasticity. Trees 2002, 16, 395–403. [Google Scholar] [CrossRef]
- Covarrubias-Cárdenas, A.G.; Martínez-Castillo, J.I.; Medina-Torres, N.; Ayora-Talavera, T.; Espinosa-Andrews, H.; García-Cruz, N.U.; Pacheco, N. Antioxidant capacity and Uplc-Pda Esi-Ms phenolic profile of Stevia rebaudiana dry powder extracts obtained by ultrasound assisted extraction. Agronomy 2018, 8, 170. [Google Scholar] [CrossRef]
- Folin, O.; Ciocalteu, V. On tyrosine and tryptophane determinations in proteins. J. Biol. Chem. 1927, 73, 627–650. [Google Scholar] [CrossRef]
- Vázquez-Flota, F.; Miranda-Ham, M.d.L.; Monforte-González, M.; Gutiérrez-Carbajal, G.; Velázquez-García, C.; Nieto-Pelayo, Y. La biosíntesis de capsaicinoides, el principio picante del chile. Rev. Fitotec. Mex. 2007, 30, 353–360. [Google Scholar] [CrossRef]
- Leakey, A.D.B.; Ainsworth, E.A.; Bernacchi, C.J.; Rogers, A.; Long, S.P.; Ort, D.R. Elevated CO2 Effects on plant carbon, nitrogen, and water relations: Six important lessons from FACE. Proc. J. Exp. Bot. 2009, 60, 2859–2876. [Google Scholar] [CrossRef]
- Mamatha, H.; Srinivasa Rao, N.K.; Laxman, R.H.; Shivashankara, K.S.; Bhatt, R.M.; Pavithra, K.C. Impact of elevated CO2 on growth, physiology, yield, and quality of tomato (Lycopersicon Esculentum Mill) cv. Arka Ashish. Photosynthetica 2014, 52, 519–528. [Google Scholar] [CrossRef]
- Vallejo-Cabrera, F.A.; Estrada-Salazar, E.I. Producción de Hortalizas de Clima Cálido, 1st ed.; Universidad Nacional de Colombia: Cali, Colombia, 2004; pp. 51–63. [Google Scholar]
- Vierling, E. The roles of heat shock proteins in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 579–620. [Google Scholar] [CrossRef]
- Blum, A.; Klueva, N.; Nguyen, H.T. Wheat cellular thermotolerance is related to yield under heat stress. Euphytica 2001, 117, 117–123. [Google Scholar] [CrossRef]
- Thomas, J.M.G.; Prasad, P.V.V.; Boote, K.J.; Allen, L.H. Seed composition, seedling emergence and earl seedling vigour of red kidne bean seed produced at elevated temperature and carbon dioxide. J. Agron. Crop. Sci. 2009, 195, 148–156. [Google Scholar] [CrossRef]
- Way, D.A.; Ladeau, S.L.; McCarthy, H.R.; Clark, J.S.; Oren, R.; Finzi, A.C.; Jackson, R.B. Greater seed production in elevated CO2 is not accompanied by reduced seed quality in Pinus taeda L. Glob. Chang. Biol. 2010, 16, 1046–1056. [Google Scholar] [CrossRef]
- Ziska, L.H.; Bunce, J.A. The Influence of elevated CO2 and temperature on seed germination and emergence from soil. Field Crop. Res. 1993, 34, 147–157. [Google Scholar] [CrossRef]
- Andalo, C.; Godelle, B.; Lefranc, M.; Mousseau, M.; Till-Bottraud, I. Elevated CO2 decreases seed germination in Arabidopsis thaliana. Glob. Chang. Biol. 1996, 2, 129–135. [Google Scholar] [CrossRef]
- Ayenan, M.A.T.; Danquah, A.; Hanson, P.; Ampomah-Dwamena, C.; Sodedji, F.A.K.; Asante, I.K.; Danquah, E.Y. Accelerating breeding for heat tolerance in tomato (Solanum lycopersicum L.): An integrated approach. Agronomy 2019, 9, 720. [Google Scholar] [CrossRef]
- Pazzagli, P.T.; Weiner, J.; Liu, F. Effects of CO2 elevation and irrigation regimes on leaf gas exchange, plant water relations, and water use efficiency of two tomato cultivars. Agric. Water Manag. 2016, 169, 26–33. [Google Scholar] [CrossRef]
- Delgado, V.; Magdaleno, J.; Ayala, O.; Garfias, D. Calidad de semillas de tres variedades nativas y una comercial de tomate producidas bajo temperaturas altas. Rev. Chapingo Ser. Hortic. 2018, 24, 215–227. [Google Scholar] [CrossRef]
- Zhou, R.; Yu, X.; Wen, J.; Jensen, N.B.; dos Santos, T.M.; Wu, Z.; Rosenqvist, E.; Ottosen, C.-O. Interactive effects of elevated CO2 concentration and combined heat and drought stress on tomato photosynthesis. BMC Plant Biol. 2020, 20, 260. [Google Scholar] [CrossRef]
- Giri, A.; Heckathorn, S.; Mishra, S.; Krause, C. Heat stress decreases levels of nutrient-uptake and-assimilation proteins in tomato roots. Plants 2017, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Heckathorn, S.A.; Giri, A.; Mishra, S.; Bista, D. Heat stress and roots. Clim. Chang. Plant Abiotic Stress Toler. 2013, 6, 109–136. [Google Scholar] [CrossRef]
- Jayawardena, D.M.; Heckathorn, S.A.; Bista, D.R.; Mishra, S.; Boldt, J.K.; Krause, C.R. Elevated CO2 plus chronic warming reduce nitrogen uptake and levels or activities of nitrogen-uptake and -assimilatory proteins in tomato roots. Physiol. Plant 2017, 159, 354–365. [Google Scholar] [CrossRef]
- Diouf, I.; Derivot, L.; Koussevitzky, S.; Carretero, Y.; Bitton, F.; Moreau, L.; Causse, M. Genetic basis of phenotypic plasticity and genotype x environment interaction in a multi-parental tomato population. J. Exp. Bot. 2020, 71, 5365–5376. [Google Scholar] [CrossRef]
- Scheiner, S.M. Genetics and Evolution of Phenotypic Plasticity. Annu. Rev. Ecol. Syst. 1993, 24, 35–68. [Google Scholar] [CrossRef]
- Via, S.; Gomulkiewicz, R.; De Jong, G.; Scheiner, S.M.; Schlichting, C.D.; Van Tienderen, P.H. Adaptive phenotypic plasticity: Consensus and controversy. Trends Ecol. Evol. 1995, 5, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Weng, L.; Li, M.; Xiao, H. Response of gene expression and alternative splicing to distinct growth environments in tomato. Int. J. Mol. Sci. 2017, 18, 475. [Google Scholar] [CrossRef]
- Pan, T.; Ding, J.; Qin, G.; Wang, Y.; Xi, L.; Yang, J.; Li, J.; Zhang, J.; Zou, Z. Interaction of Supplementary Light and CO2 Enrichment Improves Growth, Photosynthesis, Yield, and Quality of Tomato in Autumn through Spring Greenhouse Production. HortScience 2019, 54, 246–252. [Google Scholar] [CrossRef]
- Angmo, P.; Phuntsog, N.; Namgail, D.; Chaurasia, O.P.; Stobdan, T. Effect of shading and high temperature amplitude in greenhouse on growth, photosynthesis, yield and phenolic contents of tomato (Lycopersicum Esculentum Mill.). Physiol. Mol. Biol. Plants 2021, 27, 1539–1546. [Google Scholar] [CrossRef] [PubMed]
- Riga, P.; Anza, M.; Garbisu, C. Tomato quality is more dependent on temperature than on photosynthetically active radiation. J. Sci. Food Agric. 2008, 88, 158–166. [Google Scholar] [CrossRef]
- Maršić-Kacjan, N.; Gašperlin, L.; Abram, V.; Budič, M.; Vidrih, R. Quality parameters and total phenolic content in tomato fruits regarding cultivar and microclimatic conditions. Turk. J. Agric. For. 2011, 35, 185–194. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliva-Ruiz, M.; Pacheco, N.; Cuevas-Bernardino, J.C.; Tezara, W.; De-la-Peña, C.; Andueza-Noh, R.H.; Pierre, J.F.; Garruña, R. Growth, Phenotypic Plasticity and Fruit Quality in Tomato: A Study under High Temperature and Elevated CO2. Horticulturae 2023, 9, 1266. https://doi.org/10.3390/horticulturae9121266
Oliva-Ruiz M, Pacheco N, Cuevas-Bernardino JC, Tezara W, De-la-Peña C, Andueza-Noh RH, Pierre JF, Garruña R. Growth, Phenotypic Plasticity and Fruit Quality in Tomato: A Study under High Temperature and Elevated CO2. Horticulturae. 2023; 9(12):1266. https://doi.org/10.3390/horticulturae9121266
Chicago/Turabian StyleOliva-Ruiz, Miguel, Neith Pacheco, Juan Carlos Cuevas-Bernardino, Wilmer Tezara, Clelia De-la-Peña, Rubén H. Andueza-Noh, Jacques Fils Pierre, and René Garruña. 2023. "Growth, Phenotypic Plasticity and Fruit Quality in Tomato: A Study under High Temperature and Elevated CO2" Horticulturae 9, no. 12: 1266. https://doi.org/10.3390/horticulturae9121266
APA StyleOliva-Ruiz, M., Pacheco, N., Cuevas-Bernardino, J. C., Tezara, W., De-la-Peña, C., Andueza-Noh, R. H., Pierre, J. F., & Garruña, R. (2023). Growth, Phenotypic Plasticity and Fruit Quality in Tomato: A Study under High Temperature and Elevated CO2. Horticulturae, 9(12), 1266. https://doi.org/10.3390/horticulturae9121266