Abstract
Pleurotus ostreatus (Jacq.) P. Kumm is the second most cultivated edible mushroom worldwide. Temperature is one of the main environmental factors affecting the growth and fructification of mushrooms. However, the effects of ambient temperature on the growth, development, and production of bag-cultured P. ostreatus are still unclear. In this study, eight ambient temperatures (15 °C, 18 °C, 20 °C, 22 °C, 25 °C, 28 °C, 30 °C, and 32 °C) were used to cultivate P. ostreatus mycelia with cottonseed hull as substrate. Then, the agronomic traits and the activities of substrate-degrading and antioxidant enzymes were determined. The mycelial growth and fructification of P. ostreatus differed significantly under different temperatures, and 22 °C was the optimum temperature for mycelial culture when we comprehensively analyzed all the agronomic traits. The mycelial growth rate and the biological efficiency under 22 °C were 0.56 cm/d and 99.53%, respectively. Furthermore, there were also significant differences in the activities of substrate-degrading enzymes and antioxidant enzymes under different ambient temperatures. The highest activities of lignocellulose-degrading enzymes (laccase, xylanase, and cellulase), amylase, and proteinase at the vegetative growth stage occurred at 15–22 °C, 20–22 °C, and 22–30 °C, respectively. The highest activities of superoxide dismutase, peroxidase, and catalase at the vegetative growth stage occurred at 28–30 °C. This study provides a reference temperature for bag-cultured P. ostreatus.
1. Introduction
Edible mushrooms, possessing high nutritional values and medicinal properties, can degrade various lignocellulosic wastes; the cultivation of edible mushrooms produces protein-rich food and contributes to reducing environmental pollution [1,2,3,4]. For example, Pleurotus ostreatus var. florida (Jacq.) P. Kumm can be cultivated with wheat straw and three kinds of wetland plants (lake sedge, water hyacinth, and sacred lotus), and the maximum significant growth, yield, bio efficiency, proximate, and biochemical parameters were obtained from wheat straw, followed by lake sedge, sacred lotus, and water hyacinth biomass [2]. Pleurotus ostreatus can grow on cotton seed, paper waste, sawdust, and wheat straw, and the highest biological and economic yield was obtained from cotton seed [3]. Rice bran, wheat bran, barley bran, Chinese cabbage, eggshell, and soybean powder are all suitable for cultivating Hericium [4]. Nowadays, mushroom production is considered the second most important commercial microbial technology after yeast [5]. Owing to their flavor and economic, medicinal, and ecological values, edible mushrooms have been consumed and appreciated for many years. However, the cultivation of edible mushrooms is influenced by many environmental factors (e.g., temperature, humidity, illumination, CO2 density, and pH value) and the chemical composition of the medium (e.g., carbon and nitrogen sources, C/N ratio, and lignocellulose content), among which the temperature cannot be easily controlled under the mode of agricultural production, seriously affects the cultivation of edible mushrooms [6,7]. For example, Hericium grown on PDA (potato dextrose agar) medium at different temperatures (10–35 °C) showed significantly varying growth performances. The optimal temperature for Hericium growth was 25–30 °C [4]. The mycelial growth of P. ostreatus var. florida cultivated on a petri dish containing PDA medium, PDA medium with olive leaf extract and/or green tea leaf extract at 22 °C, 25 °C, and 28 °C inoculation temperatures showed significant differences, and mycelia grew the fastest at 28 °C [6]. In a previous study, maximum mycelial growth was reported at 25 °C for most of the Pleurotus strains tested, whereas the maximum growth of many strains was at 30 °C [7].
P. ostreatus (Jacq.) P. Kumm is the second most cultivated edible mushroom worldwide after Agaricus bisporus. The production of bag-cultured P. ostreatus can be divided into two periods (the vegetative growth period and the reproductive period). Since the growth of P. ostreatus is greatly affected by seasonal temperature changes, its production (especially inoculation) is mainly carried out in autumn and spring in China. As previously reported, the effects of different temperatures on the mycelial growth of edible mushrooms grown on agar plates have been widely studied; however, the exact effects and biochemical mechanisms of the ambient temperature on the mycelial growth of bag-cultured P. ostreatus and other edible mushrooms during the vegetative growth period, and the effects of the mycelial culture temperature on the fructification and yield of P. ostreatus, were all still unclear. In this study, eight ambient temperatures (15 °C, 18 °C, 20 °C, 22 °C, 25 °C, 28 °C, 30 °C, and 32 °C) were used to cultivate P. ostreatus mycelia with cottonseed hull (the optimal substrate for P. ostreatus cultivation [3]) as substrate. Several agronomic traits (including the mycelial growth rate, growth vigor, primordial formation time, fruiting body morphology, and biological efficiency, etc.) and the activities of substrate-degrading enzymes and antioxidant enzymes were determined. The results of this study will provide reference temperatures for the production of bag-cultured P. ostreatus.
2. Materials and Methods
2.1. Biological Material and Culture Conditions
P. ostreatus strain New 831 was obtained from the College of Life Science, Henan Agricultural University. Pure culture of P. ostreatus was obtained on the mushroom complete medium (5 g L−1 yeast extract, 20 g L−1 glucose, 0.01 g L−1 thiamine hydrochloride, 0.5 g L−1 magnesium sulfate heptahydrate [MgSO4·7H2O], 1 g L−1 potassium dihydrogen phosphate [KH2PO4], and 2% Agar) at 25 °C for 7 d [8]. Then, the activated P. ostreatus mycelia were inoculated in the breathable polypropylene bags (18 cm × 36 cm × 0.004 cm) containing sterilized cottonseed hull substrates (97% cottonseed hull, 2% lime, 1% calcium carbonate, and 65% water). After inoculation, the bags were incubated at 25 °C for about 30 d until the mycelia fully colonized.
2.2. Temperature Treatment
The cottonseed hull substrates mentioned above were prepared and transferred to the breathable polypropylene bags (18 cm × 36 cm × 0.004 cm) with 0.36 kg of dry material per bag. The bags were then autoclaved for sterilization at 0.14 MPa and 126 °C for 2.5 h. When the temperature of the bags dropped below 30 °C, they were fumigated with sodium dichloroisocyanurate (10 g m−3) in the inoculation room for 30 min and inoculated through the sterile end with the same inoculation amount (about 50 g per bag) [3]. After inoculation, the bags were incubated at different temperatures (15 °C, 18 °C, 20 °C, 22 °C, 25 °C, 28 °C, 30 °C, and 32 °C) to initiate mycelium growth. The germination of strains and mycelial growth were observed regularly. The experiment was performed independently in triplicate with 30 bags per treatment.
2.3. Mycelial Linear Growth Rate Measurements
The mycelial linear growth distance, expressed in cm, was measured when the mycelia fully colonized the substrate. Then, the mycelial linear growth rate of P. ostreatus, expressed in cm/d, was calculated following the equation mentioned by Werghemmi et al. [6]. The experiments were performed independently in triplicate with 10 bags per treatment.
2.4. Biological Efficiency Measurements
After the mycelia fully colonized the substrates, the mushroom bags were transferred to the mushroom house to induce primordia formation with the conditions of 15 °C temperature, 90% relative humidity, scattering light, and fresh air. The first three flushes of the fruiting body were harvested once they reached remarkable size and color, and the fresh weight of the fruiting body harvested from one bag was weighed. Then, the biological efficiency (%) was calculated following the equation mentioned by Elbagory et al. [3]. The experiment was performed independently in triplicate with 30 bags per experiment.
2.5. Enzyme Activity Assays
The enzymes’ activities were analyzed using the hyphae samples taken from the substrate at different stages: I, the stage of substrate half-colonized by mycelia; II, the stage of substrate fully colonized by mycelia; III, the primordial stage; IV, the first harvest stage; V, the second harvest stage. The extracellular enzymes were extracted as follows: 5 g of mycelial colonized substrate was mixed with 25 mL of 100 mol L−1 phosphate buffer (pH 6.5) and incubated at 4 °C for about 3 h with a speed of 150 r min−1, then the mixture was centrifugated at 4 °C for 25 min with a speed of 15,000× g. The supernatant was transferred into a new tube and used to determine the activities of substrate-degrading enzymes. The intracellular enzymes were prepared according to the following protocol: 1 g of the mycelial colonized substrate was frozen in liquid nitrogen and ground in powder, 5 mL of 100 mol L−1 phosphate buffer (pH 6.5) was added and mixed thoroughly for 5 min, and then the mixture was centrifugated at 4 °C for 25 min with a speed of 15,000× g. The supernatant was transferred into a new tube and used to determine the activities of antioxidant enzymes. The protein concentration of the extracellular and intracellular enzymes was determined using the Bradford method with bovine serum albumin as a standard [9].
The activities of superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD) enzymes were detected using the corresponding activity assay kits (Nanjing Jiancheng Bioengineering Institute): Superoxide Dismutase [SOD] assay kit [WST-1 method], A001–3–2; Catalase [CAT] assay kit [Visible light], A007–1–1; and Peroxidase assay kit, A084–3–1, following the manufacturer’s protocol. The absorption values were observed at 560 nm to indicate the enzyme levels required to induce 50% inhibition of nitro blue tetrazolium reduction and calculate the SOD activity. The rate of hydrogen peroxide (H2O2) decomposition was measured at 240 nm to calculate the CAT activity. POD activity in the enzyme extracts was measured based on changes in absorbance at 470 nm in the mixtures consisting of 50 mmol L−1 phosphate buffer (pH 5.0), 40 mmol L−1 H2O2, 20 mmol L−1guaiacol, and 100 µL of enzyme extract. The specific activities of the above enzymes were calculated and defined based on the protein levels (U mg−1 protein) according to the method given by Abbas et al. [10]. All experiments were conducted in triplicate.
The specific activities of protease, amylase, laccase, xylanase, and carboxymethyl cellulase (CMCase) were measured according to the previous protocols described and defined based on the protein levels (U mg−1 protein), with some modifications [11,12,13,14]. Briefly, laccase activity was measured using ABTS 2,2′-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) (Sigma-Aldrich, St. Louis, MI, USA) as the substrate [11,12]. The reaction mixture contained 2.7 mL of 100 mmol/L sodium acetate buffer (pH 5.0), 200 μL of 1.0 mmol/L ABTS, and 100 μL of extracellular enzymes, and the increase in absorbance at 420 nm within 1 min was determined to monitor the oxidation of ABTS. The molar extinction coefficient (ε420 nm = 36,000 L (mol cm)−1) of ABTS radical was used to calculate the laccase activity. The activities of xylanase, CMCase, and amylase were determined using carboxylmethyl cellulose, oat spelt xylan, and soluble starch (Sigma-Aldrich, USA) as substrates, respectively [13]. The reaction mixture was prepared by mixing 80 μL of extracellular enzymes, 0.5 mL of substrate solution (10 g L−1), and 0.5 mL of 50 mmol L−1 sodium acetate buffer (pH 5.0), then incubated at 37 °C for 30 min. Subsequently, 1.5 mL of DNS (3,5-dinitrosalicylic acid) solution was added and boiled for 10 min to stop the reaction. The absorbance at 540 nm was measured, and the production of glucose or xylose was calculated using the glucose or xylose standard curves. The reaction mixture containing inactivated enzymes (boiled for 10 min), substrate, and buffer was used as the control. Protease activity was determined according to the casein-Folin method [14]. The reaction mixture containing 0.1 mL of extracellular enzymes, 0.9 mL of 100 mmol L−1 sodium phosphate buffer (pH 7.0), and 2 mL of 0.5% casein was incubated at 37 °C for 10 min, then 3 mL of 10% TCA (Trichloroacetic acid) solution was added and incubated at 37 °C for 20 min to stop the reaction. After centrifugation at 12,000× g for 5 min, 0.1 mL of supernatant was mixed with 0.5 mL of 0.4 mol L−1 sodium carbonate solution and 0.1 mL of Folin-Ciocalteu’s phenol reagent (Sangon Biotech, Shanghai, China), then incubated at 37 °C for 20 min. Finally, the absorbance at 680 nm was measured and used to calculate the production of tyrosine using the tyrosine standard curve. All analyses were performed in triplicate.
2.6. Statistical Analysis
GraphPad Prism 6.0 (GraphPad Software, Inc., San Diego, CA, USA) was used for the statistical analyses in this study. The error bars indicate the standard deviation from the mean of the triplicates. The differences at p <0.05 were considered significant according to Turkey’s test (SPSS, Version 26.0, IBM Corporation, Armonk, NY, USA) and were marked with different letters.
3. Results
3.1. Effects of Mycelial Culture Temperature on the Mycelial Growth of P. ostreatus
The cultivation bags incubated at different temperatures showed significant variations in the mycelium germination time, the mycelial growth vigor and growth rate, and the time required for full mycelial colonization (Table 1). The mycelia of P. ostreatus germinated rapidly under 22–32 °C cultivation temperatures, and germination occurred one day after inoculation. Germination occurred within two days when mycelia were incubated at 15 °C and 18 °C. With increasing temperatures, the mycelial growth rate increased first and then decreased. The fastest mycelial growth rate occured at 28 °C, which was 0.65 cm d−1, followed by 25 °C, 30 °C, 22 °C, 20 °C, 32 °C, 18 °C, and 15 °C. The time required for full mycelial colonization decreased first and then increased with increasing temperatures. The shortest time required for full mycelial colonization occurred at 28 °C, followed by 25 °C, 30 °C, 22 °C, 20 °C, 32 °C, 18 °C, and 15 °C. This variation was consistent with the mycelial growth rate. The culture temperature also had a significant effect on the mycelial growth vigor (Table 1). The mycelia grew more vigorously under low culture temperatures. The mycelia were white, dense, robust, and with neat tips when cultured at 15–25 °C. When the culture temperature was above 28 °C, the mycelial growth vigor gradually weakened. By comprehensive comparison, a temperature of 20–25 °C was determined as the suitable temperature for the mycelial growth of P. ostreatus.

Table 1.
Effects of mycelial culture temperature on the mycelial growth of P. ostreatus.
3.2. Effects of Mycelial Culture Temperature on Fructification of P. ostreatus
The fructification and yield of edible mushrooms are closely related to mycelial growth. To investigate the effects of mycelial culture temperature on the fructification of P. ostreatus, the mycelia fully colonized mushroom bags cultivated at different ambient temperatures were moved to the mushroom house to induce fructification, and the primordial formation time of the first flush, the fruiting body morphology, and the biological efficiency were determined. Results showed that the primordial formation time of the first flush and the biological efficiency significantly varied under different temperatures (Table 2). The primordial formation time was closely related to the time required for full mycelial colonization. The fastest primordial formation time was recorded at 25 °C and 28 °C, followed by 22 °C and 30 °C, 20 °C, 32 °C, 18 °C, and 15 °C. The culture temperature had no significant effects on the fruiting body morphology, which were gray, thick pileus, and short stipe.
Table 2.
Effects of mycelial culture temperature on fructification of P. ostreatus.
However, the biological efficiency of P. ostreatus gradually decreased with the increasing mycelial culture temperature. The highest biological efficiency (109.58%) was obtained at 15 °C, and the lowest (79.73%) occurred at 35 °C. The biological efficiency of P. ostreatus cultivated at 20 °C, 22 °C, and 25 °C were 102.16%, 99.53%, and 97.15%, respectively. Comprehensive analyses showed that for high biological efficiency and shorter fruiting body formation time, 22 °C was the most optimal temperature for the mycelial culture.
3.3. Effects of Mycelial Culture Temperature on the Activities of Substrate-Degrading Enzymes
P. ostreatus can obtain carbon and nitrogen sources from the substrate by degrading lignocelluose, starch, and protein, so the activities of substrate-degrading enzymes are closely related to the mycelial growth and yield of P. ostreatus. Therefore, the activities of the main substrate-degrading enzymes (including laccase, xylanase, cellulase, amylase, and protease) produced by P. ostreatus mycelia cultivated at different temperatures were determined. As shown in Figure 1, the activities of lignocelluose-degrading enzymes (laccase, xylanase, and CMCase) were significantly affected by the mycelial culture temperature.
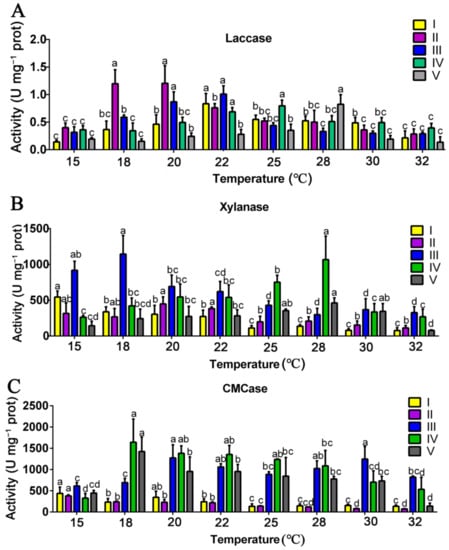
Figure 1.
Effects of the mycelial culture temperature on the activities of lignocellulose-degrading enzymes. Activities of laccase (A), xylanase (B), and CMCase (C) at different mycelial culture temperatures at different growth and developmental stages of P. ostreatus. I, the stage of substrate half colonized by mycelia; II, the stage of substrate fully colonized by mycelia; III, the primordial; IV, the first harvest stage; V, the second harvest stage. The data are presented as the means from three independent measurements ± the standard deviations (indicated by the error bars). Different letters indicate significant differences between strains (p < 0.05, as determined by Tukey’s test).
The laccase activity at stages I to V increased at first and then decreased with the increasing mycelial culture temperature (Figure 1A). At stage I, the highest laccase activity occurred at 22 °C (0.73 U mg−1 prot) and the lowest one occurred at 15 °C and 32 °C. At stage II, the highest laccase activity occurred at 18 °C (1.19 U mg−1 prot) and 20 °C (1.20 U mg−1 prot), and the lowest one occurred at 15 °C, 30 °C, and 32 °C. At stage III, the highest laccase activity occurred at 20 °C (0.87 U mg−1prot) and 22 °C (1.01 U mg−1 prot), and the lowest one occurred at 15 °C, 28 °C, 30 °C, and 32 °C. At stage IV, the highest laccase activity occurred at 22 °C (0.70 U mg−1 prot) and 25 °C (0.79 U mg−1 prot). At stage V, the highest laccase activity occurred at 28 °C (0.82 U mg−1 prot).
With the increasing mycelial culture temperature, the xylanase activity at stage I gradually decreased, while those at stages II to V almost all increased first and then decreased (Figure 1B). The highest xylanase activity at stage II occurred at 20 °C (447.49 U mg−1 prot) and 22 °C (423.49 U mg−1 prot), that at stage III occurred at 18 °C (1144.12 U mg−1 prot), that at stage IV occurred at 28 °C (1067.29 U mg−1 prot), and that at stage V also occurred at 28 °C (462.01 U mg−1 prot).
The effects of the mycelial culture temperature on cellulase were determined by measuring the CMCase activity. With the increasing mycelial culture temperature, the CMCase activity at stage I and stage II all decreased, while those at stages III to V almost all increased first and then decreased. The highest CMCase activity at stage III occurred at 20 °C (1273.13 U mg−1 prot) and 30 °C (1246.77 U mg−1 prot), while those at stages IV and V all occurred at 18 °C (1642.47 and 1424.14 U mg−1 prot).
Based on the present results, it can be seen that the mycelia at the vegetative growth stages (stages I and II) cultivated at 15–22 °C had relatively higher activities of laccase, xylanase, and CMCase, which may be related to the higher biological efficiency at those temperatures.
As shown in Figure 2, the activities of amylase and protease were also significantly affected by the mycelial culture temperature. The amylase activity at stages I, II, IV, and V all increased first and then decreased with the increasing mycelial culture temperature (Figure 2A). The peak of amylase activity at stage I occurred at 20 °C and 22 °C (296.43 and 300.65 U mg−1 prot, respectively), and those at stages II, IV, and V occurred at 22 °C (667.73 U mg−1 prot), 28 °C (479.23 U mg−1 prot), and 18 °C (807.03 U mg−1 prot), respectively. The protease activity at stages I, II, IV, and V also increased first and then decreased. The highest protease activity at stages I, II, IV, and V occurred at 22 °C (825.45 U mg−1 prot), 28 °C (814.27 U mg−1 prot), 25 °C (992.65 U mg−1 prot), and 30 °C (210.36 U mg−1 prot), respectively. Based on the present results, mycelia at the vegetative growth stages (stages I and II) cultivated at 20 and 22 °C had relatively higher amylase activity, and those cultivated at 22–30 °C had relatively higher protease activity.
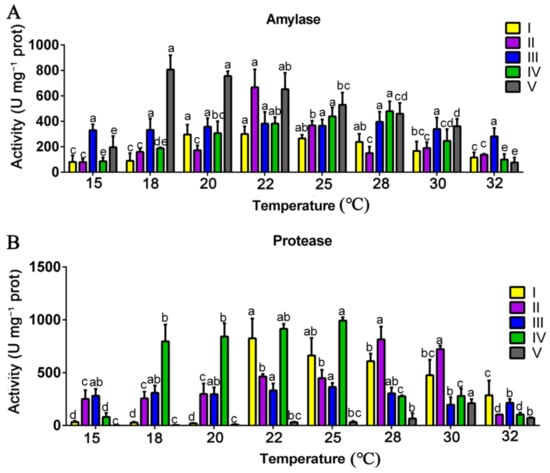
Figure 2.
Effects of the mycelial culture temperature on the activities of other hydrolytic enzymes. Activities of amylase (A) and protease (B) at different mycelial culture temperatures at different growth and developmental stages of P. ostreatus. I, the stage of substrate half colonized by mycelia; II, the stage of substrate fully colonized by mycelia; III, the primordial stage; IV, the first harvest stage; V, the second harvest stage. The data are presented as the means from three independent measurements ± the standard deviations (indicated by the error bars). Different letters indicate significant differences between strains (p < 0.05, as determined by Tukey’s test).
3.4. Effects of Mycelial Culture Temperature on the Activities of Antioxidant Enzymes
An unfavorable high ambient temperature can stimulate the antioxidant system (including antioxidant enzymes and antioxidant compounds) in P. ostreatus to resist the adverse environment [8]. Therefore, the activities of antioxidant enzymes (SOD, POD, and CAT) in mycelia cultivated at different ambient temperatures were determined. The activities of three antioxidant enzymes were all significantly affected by different mycelial growth temperatures (Figure 3). As shown in Figure 3A, the SOD activity at stages I and III remained stable at 15–22 °C and increased significantly at 25–32 °C. At stage II, the SOD activity remained stable at 15–25 °C, increased significantly at 28–30 °C, and then gradually decreased to a low level. The SOD activity at stage IV was the highest at 18 °C, 20 °C, and 22 °C, and that at stage V was the highest at 30 °C and 32 °C. As shown in Figure 3B, the POD activity at the vegetative growth stages (stages I and II) was significantly higher at 28–30 °C. It was relatively stable at the reproductive growth stages (stages III to V). The highest POD activity at stage I occurred at 30 °C, while that at stage II occurred at 28 °C. As shown in Figure 3C, the CAT activity at stage I was significantly higher at 25–30 °C, and that at stage II was significantly improved when the mycelial culture temperature was above 22 °C. The CAT activity was relatively stable at the reproductive growth stages (stages III to V). Based on the present results, it can be seen that the mycelia at the vegetative growth stages (stages I and II) cultivated at 28–30 °C had relatively higher activities of antioxidant enzymes.
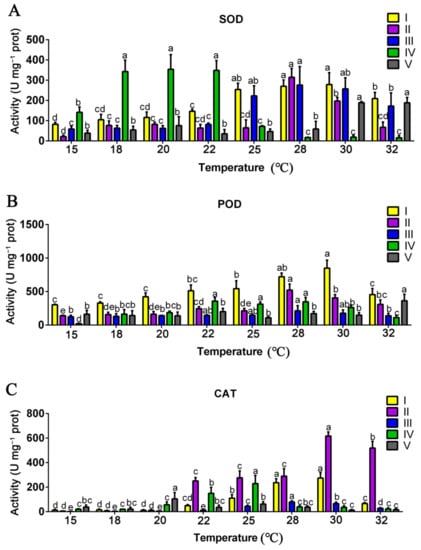
Figure 3.
Effect of mycelial culture temperature on the activities of antioxidant enzymes. Activities of SOD (A), POD (B), and CAT (C) at different mycelial culture temperatures at different growth and developmental stages of P. ostreatus. CAT: catalase; SOD: superoxide dismutase; POD: guaiacol peroxidase. I, the stage of substrate half colonized by mycelia; II, the stage of substrate fully colonized by mycelia; III, the primordial stage; IV, the first harvest stage; V, the second harvest stage. The data are presented as the means from three independent measurements ± the standard deviations (indicated by the error bars). Different letters indicate significant differences between strains (p < 0.05, as determined by Tukey’s test).
4. Discussion
Temperature is an important environmental factor that affects the growth and productivity of edible mushrooms. Temperatures higher or lower than the optimum temperature can affect the physiological and biochemical processes of plants and edible mushrooms [15,16,17,18]. However, the effects and mechanism of the mycelial culture temperature on the mycelial growth and fructification of bag-cultured mushrooms have not been systematically studied. In this study, P. ostreatus, the second most cultivated edible mushroom worldwide, was used as the research object, and the effects of different mycelial culture temperatures (15–32 °C) on the main agronomic traits, and the activities of substrate-degrading enzymes and antioxidant enzymes of bag-cultured P. ostreatus, were determined.
The mycelial growth rate and growth vigor of P. ostreatus were all significantly affected by the mycelial culture temperature; the mycelial growth rates of P. ostreatus cultivated at 20–30 °C were all above 0.5 cm/d, and the highest growth rate occurred at 28 °C. The highest growth rate of P. ostreatus var. florida, P. ostreatus, and P. cystidiosus cultivated on PDA medium were also obtained at 28 °C [6,19], which indicates that the optimum temperature for mycelial growth of Pleurotus spp. cannot be affected by the medium and cultivation method. This contradicts previous studies that outlined the direct effect of raw materials on the substrate temperature, mycelial colonization, and yield [20]. Additionally, the mycelial growth vigor, another important index for the mycelial growth of edible mushrooms, changed significantly when P. ostreatus was cultivated at different temperatures. The mycelial growth vigor of P. ostreatus cultivated at 15–25 °C was significantly stronger than that cultivated at high temperatures (28–32 °C), which is somewhat different from the mycelial growth vigor of P. ostreatus cultivated on PDA medium [19]. Such differences may be caused by different culture media or species. This can be because the mycelial growth vigor was different under different cultivation substrates. By comprehensively comparing the mycelial growth rate and growth vigor, it can be seen that the suitable temperature for mycelium culture was 20–25 °C.
The primordial formation time and the biological efficiency were significantly different under different mycelial culture temperatures. The primordial formation time of P. ostreatus was negatively related to the mycelial growth rate, and the shortest primordial formation time (35 d in this study) was recorded at 25 °C and 28 °C. Therefore, a high mycelial growth rate can significantly shorten the whole production time of P. ostreatus and reduce the energy consumption and cost under industrial production. The biological efficiency of P. ostreatus gradually decreased with the increase of the mycelial culture temperature, which decreased the economic efficiency of P. ostreatus production. Based on the agronomic traits, it can be seen that the optimum mycelial culture temperature for bag-cultured P. ostreatus was 22 °C, because P. ostreatus showed a relatively higher growth rate, stronger growth vigor, shorter primordial formation time, and higher biological efficiency when cultivated at this temperature. Therefore, the mycelial culture temperature of bag-cultured P. ostreatus under industrial production should be controlled at 22 °C.
Various studies have been performed to study the chemical composition of medium (types and amounts of carbon and nitrogen, C/N ratio, metals, xenobiotics, lignocellulose content and composition, and pH, etc.) on the production of substrate-degrading enzymes [21,22,23,24,25,26,27,28] and the reaction temperatures on the activities of such enzymes [29]. However, the effects of the mycelial culture temperature on the production (or the specific activities) of substrate-degrading enzymes in bag-cultured P. ostreatus have not been revealed until now. The results of this study showed that the peak of laccase activity was obtained at the vegetative growth stage, while the peaks of xylanase, CMCase, amylase, and protease all occurred at the reproductive growth stage, which indicated that the expression of laccase was earlier than other substrate-degrading enzymes; this phenomenon has been reported elsewhere [24,30]. The polymer lignin compounds from the substrate are broken down into simpler carbon molecules which are further taken by mycelia and help in mushroom growth [31]. Therefore, the laccase activity is important to the mycelial growth of P. ostreatus. The present results also showed that the laccase activity at the vegetative growth stage was significantly higher when P. ostreatus was cultivated at 18–22 °C, which indicated that the relatively lower mycelial culture temperature is conducive to the high expression of laccase at such stage and can improve the selectivity of delignification of P. ostreatus. Xylanase is a major hemicellulase that acts on the backbone of xylan to produce xylooligosaccharides [32], which are one kind of carbon and energy source for the growth and development of edible mushrooms. The xylanase activity at the vegetative growth stage was higher when P. ostreatus was cultivated at 15–22 °C. Cellulose serves as the main carbon and energy source for the growth and development of P. ostreatus [24,30], and high cellulose degradation during the reproductive stage is positively correlated with the yield of edible mushrooms [23,33]. The present results showed that the CMCase activity at the reproductive growth stage was significantly higher than that at the vegetative growth stage, which is consistent with the findings on Morchella importuna [34]. Similar to the xylanase activity, the CMCase activity at the vegetative growth stage was also higher when P. ostreatus was cultivated at 15–22 °C, which suggests that the expression of xylanase and CMCase in bag-cultured P. ostreatus can be enhanced by low mycelial culture temperature. Therefore, the high biological efficiency of P. ostreatus under 15–22 °C of mycelial culture temperatures may be related to the high activities of xylanase and CMCase under those temperatures.
Starch in cultivation substrates can be degraded by amylase to provide carbon and energy for microbes [26], and protein in cultivation substrates can be degraded by protease to provide a nitrogen source for microbes [27]. The present findings showed that the activities of amylase and protease at the vegetative growth stage all increased first and then decreased with the increase of mycelial culture temperature; the highest activities of amylase and protease were obtained at 20–22 °C and 22–30 °C, respectively.
Excessive ROS (reactive oxygen species) may be accumulated in edible mushrooms under unfavorable high ambient temperatures. Then, the antioxidant enzymes will be activated to remove excessive ROS [8,17]. SOD can disproportionate the superoxide radical to hydrogen peroxide, and then CAT and peroxidases (such as POD) convert the toxic hydrogen peroxide to water [10,35]. The present findings showed that the activities of SOD, POD, and CAT in P. ostreatus were relatively higher at the vegetative growth stage when cultivated at 28–30 °C. This may be related to the excess accumulation of ROS under high temperatures. It is noteworthy that the SOD activity was significantly improved at the first harvest stage when P. ostreatus mycelia were cultivated at 15–22 °C, which may be related to the fast development of fruiting bodies. The relationship between the expression of SOD isoenzymes and the fruiting body development of Volvariella volvacea, and the relationship between the SOD activity and the fruiting body development of Hypsizygus marmoreus have been previously reported [17,36]. Therefore, SOD plays important roles both in protecting P. ostreatus mycelia against the adverse effect of high temperature and in the development of the fruiting body.
5. Conclusions
Temperature is one of the most important environmental factors that affect the growth and development of edible mushrooms. The effects and mechanism of the mycelial culture temperature on the mycelial growth and fructification of bag-cultured P. ostreatus were systematically studied in this study. The mycelial growth vigor and growth rate, primordial formation time, biological efficiency, and other agronomic traits of P. ostreatus were all significantly affected by the mycelial culture temperature, and P. ostreatus cultivated at 22 °C performed best with relatively higher growth rate, stronger growth vigor, shorter primordial formation time, and higher biological efficiency. Additionally, the activities of substrate-degrading enzymes and antioxidant enzymes were also significantly affected by the mycelial culture temperature. At the vegetative growth stage, P. ostreatus cultivated at low temperatures (15–22 °C) had significantly higher activities of lignocellulose-degrading enzymes, which may be the reason for the high biological efficiency of P. ostreatus under those temperatures. However, the activities of antioxidant enzymes at the vegetative growth stage were improved by high mycelial culture temperatures. This study provides a reference temperature for the mycelia cultivaiton of bag-cultured P. ostreatus. However, further research is suggested with molecular analysis to have a detailed insight into the mycelial culture temperature-induced changes responsible for the enhancement of growth and yield in Pleurotus ostreatus (Jacq.) P. Kumm.
Author Contributions
Q.W. and Y.H. designed and directed the experiment. W.Z. and Y.C. performed the experiments. Y.H., Y.Q. and W.Z. analyzed the data. Q.W. and J.S. provided materials, instruments, and funds. Q.W., Y.H. and F.W. wrote and modified the manuscript. Validation and Data curation, F.X. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the China Agriculture Research System of MOF and MARA (No. CARS-20), the National Natural Science Foundation of China (No.32202571), the Major Public Welfare Special Projects in Henan Province (No. 201300110700), the Joint Research Project for Improved Agricultural Variety in Henan Province (No. 2022030101), and the Key Program for Science and Technology Development of Henan Province (No. 212102110395).
Institutional Review Board Statement
This article does not contain any studies with human participants or animals performed by any of the authors.
Data Availability Statement
Not applicable.
Conflicts of Interest
All authors declare that they have no conflict of interest.
References
- Gupta, S.; Summuna, B.; Gupta, M.; Annepu, S.K. Edible mushrooms: Cultivation, bioactive molecules, and health benefits. In Bioactive Molecules in Food; Reference series in Phytochemistry; Springer: Berlin/Heidelberg, Germany, 2018; pp. 1–33. [Google Scholar]
- Elbagory, M.; El-Nahrawy, S.; Omara, A.E.D.; Eid, E.M.; Bachheti, A.; Kumar, P.; Abou Fayssal, S.; Adelodun, B.; Bachheti, R.K.; Kumar, P.; et al. Sustainable bioconversion of wetland plant biomass for Pleurotus ostreatus var. flsorida cultivation: Studies on proximate and biochemical characterization. Agriculture 2022, 12, 2095. [Google Scholar] [CrossRef]
- Girmay, Z.; Gorems, W.; Birhanu, G.; Zewdie, S. Growth and yield performance of Pleurotus ostreatus (Jacq. Fr.) Kumm (oyster mushroom) on different substrates. AMB Express 2016, 6, 87. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.G.; Park, H.G.; Park, S.H.; Choi, C.W.; Kim, S.H.; Park, W.M. Comparative study of mycelial growth and basidiomata formation in seven different species of the edible mushroom genus Hericium. Bioresour. Technol. 2005, 96, 1439–1444. [Google Scholar] [CrossRef] [PubMed]
- Pathak, R.; Joshi, N.; Dwivedi, R.R. Eco-friendly production of Agaricus bisporus (lange) imbach (white button mushroom). Nat. Sci. 2009, 6, 57–60. [Google Scholar]
- Werghemmi, M.; Fayssal, S.A.; Mazouz, H.; Hajjaj, H.; Hajji, L. Olive and green tea leaves extract in Pleurotus ostreatus var. florida culture media: Effect on mycelial linear growth rate, diameter and growth induction index. IOP Conf. Ser. Earth Environ. Sci. 2022, 1090, 012020. [Google Scholar] [CrossRef]
- Zharare, G.E.; Kabanda, S.M.; Poku, J.Z. Effects of temperature and hydrogen peroxide on mycelial growth of eight Pleurotus strains. Sci. Hortic. 2010, 125, 95–102. [Google Scholar] [CrossRef]
- Hu, Y.R.; Wang, Y.; Chen, Y.J.; Chai, Q.Q.; Dong, H.Z.; Shen, J.W.; Qi, Y.C.; Wang, F.Q.; Wen, Q. Salicylic acid enhances heat stress resistance of Pleurotus ostreatus (Jacq.) P. Kumm through metabolic rearrangement. Antioxidants 2022, 11, 968. [Google Scholar] [CrossRef]
- Hammond, J.B.; Kruger, N.J. The bradford method for protein quantitation. Methods Mol. Biol. 1988, 3, 25–32. [Google Scholar]
- Abbas, M.; Imran, F.; Iqbal Khan, R.; Zafar-ul-Hye, M.; Rafique, T.; Jameel Khan, M.; Taban, S.; Danish, S.; Datta, R. Gibberellic acid induced changes on growth, yield, superoxide dismutase, catalase, and peroxidase in fruits of bitter gourd (Momordica charantia L.). Horticulturae 2020, 6, 72. [Google Scholar] [CrossRef]
- Patrick, F.; Mtui, G.; Mshandete, A.M.; Kivaisi, A. Optimized production of lignin peroxidase, manganese peroxidase and laccase in submerged cultures of Trametes trogii using various growth media compositions. Tanzan J. Sci. 2010, 36, 1–18. [Google Scholar]
- Baldrian, P. Increase of laccase activity during interspecific interactions of white-rot fungi. FEMS. Microbiol. Ecol. 2004, 50, 245–253. [Google Scholar] [CrossRef]
- Giraldo, L.A.; Tejido, M.L.; Ranilla, M.J.; Carro, M.D. Effects of exogenous fibrolytic enzymes on in vitro ruminal fermentation of substrates with different forage:concentrate ratios. Anim. Feed Sci. Technol. 2008, 141, 306–325. [Google Scholar] [CrossRef]
- Kasana, R.C.; Salwan, R.; Yadav, S.K. Microbial proteases: Detection, production, and genetic improvement. Crit. Rev. Microbiol. 2011, 37, 262–276. [Google Scholar] [CrossRef]
- Almeselmani, M.; Deshmukh, P.S.; Sairam, R.K.; Kushwaha, S.R.; Singh, T.P. Protective role of antioxidant enzymes under high temperature stress. Plant Sci. 2006, 171, 382–388. [Google Scholar] [CrossRef]
- Lee, D.H.; Lee, C.B. Chilling stress-induced changes of antioxidant enzymes in the leaves of cucumber: In gel enzyme activity assays. Plant Sci. 2000, 159, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.J.; Zhang, L.; Wang, R.Q.; Xie, B.; Li, X.; Chen, R.L.; Guo, L.X.; Xie, B.G. The sequence characteristics and expression models reveal superoxide dismutase involved in cold response and fruiting body development in Volvariella volvacea. Int. J. Mol. Sci. 2016, 17, 34. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Zhao, M.; Wu, X.; Zhang, J. Metabolic response of Pleurotus ostreatus to continuous heat stress. Front. Microbiol. 2020, 10, 3148. [Google Scholar] [CrossRef]
- Hoa, H.T.; Wang, C.L. The effects of temperature and nutritional conditions on mycelium growth of two oyster mushrooms (Pleurotus ostreatus and Pleurotus cystidiosus). Mycobiology 2015, 43, 14–23. [Google Scholar] [CrossRef]
- Abou Fayssal, S.; Alsanad, M.A.; Yordanova, M.H.; El Sebaaly, Z.; Najjar, R.; Sassine, Y.N. Effect of olive pruning residues on substrate temperature and production of oyster mushroom (Pleurotus ostreatus). Acta Hortic. 2021, 1327, 245–252. [Google Scholar] [CrossRef]
- Sadh, P.K.; Duhan, S.; Duhan, J.S. Agro-industrial wastes and their utilization using solid state fermentation: A review. Bioresour. Bioprocess. 2018, 5, 1. [Google Scholar] [CrossRef]
- Economou, C.N.; Diamantopoulou, P.A.; Philippoussis, A.N. Valorization of spent oyster mushroom substrate and laccase recovery through successive solid state cultivation of Pleurotus, Ganoderma, and Lentinula strains. Appl. Microbiol. Biot. 2017, 101, 5213–5222. [Google Scholar] [CrossRef]
- Kurt, S.; Buyukalaca, S. Yield performances and changes in enzyme activities of Pleurotus spp. (P. ostreatus and P. sajorcaju) cultivated on different agricultural wastes. Bioresource Technol. 2010, 101, 3164–3169. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Ma, F.; Li, Y.; Yu, H.; Li, C.; Zhang, X. Differential proteomic profiles of Pleurotus ostreatus in response to lignocellulosic components provide insights into divergent adaptive mechanisms. Front. Microbiol. 2017, 8, 480. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Xu, L.; Zhao, L.; Ding, Z.; Ma, H.; Terry, N. Fungal laccase production from lignocellulosic agricultural wastes by solid-state fermentation: A review. Microorganisms 2019, 7, 665. [Google Scholar] [CrossRef] [PubMed]
- Ergun, S.O.; Urek, R.O. Production of ligninolytic enzymes by solid state fermentation using Pleurotus ostreatus. Ann. Agrar. Sci. 2017, 15, 273–277. [Google Scholar] [CrossRef]
- Dosoretz, C.G.; Chen, H.C.; Grethlein, H.E. Effect of environmental conditions on extracellular protease activity in ligninolytic cultures of Phanerochaete chrysosporium. Appl. Environ. Microbiol. 1990, 56, 395–400. [Google Scholar] [CrossRef]
- Haider, A.; Alam, M.M.; Khan, A.A.; Zulfiqar, M.A. Optimization of cultural conditions for the treatment of pulp and paper industrial effluent by Pleurotus ostreatus (L.). Pak. J. Agric. Res. 2019, 32, 507–513. [Google Scholar] [CrossRef]
- Zhuo, R.; Yu, H.; Qin, X.; Ni, H.; Jiang, Z.; Ma, F.; Zhang, X. Heterologous expression and characterization of a xylanase and xylosidase from white rot fungi and their application in synergistic hydrolysis of lignocellulose. Chemosphere 2018, 212, 24–33. [Google Scholar] [CrossRef]
- Pandey, V.K.; Singh, M.P. Biodegradation of wheat straw by Pleurotus ostreatus. Cell Mol. Biol. 2014, 60, 29–34. [Google Scholar]
- Kumar, P.; Eid, E.M.; Al-Huqail, A.A.; Širić, I.; Adelodun, B.; Abou Fayssal, S.; Kumar, V. Kinetic Studies on Delignification and Heavy Metals Uptake by Shiitake (Lentinula edodes) Mushroom Cultivated on Agro-Industrial Wastes. Horticulturae 2022, 8, 316. [Google Scholar] [CrossRef]
- Jiang, W.; Pei, R.; Zhou, S.F. 3D-printed xylanase within biocompatible polymers as excellent catalyst for lignocellulose degradation. Chem. Eng. J. 2020, 400, 125920. [Google Scholar] [CrossRef]
- Öztürk, C.; Atila, F. Changes in lignocellulosic fractions of growing substrates during the cultivation of Hypsizygus ulmarius mushroom and its effects on mushroom productivity. Sci. Hortic. 2021, 288, 110403. [Google Scholar] [CrossRef]
- Hao, H.; Zhang, J.; Wang, H.; Wang, Q.; Chen, M.; Juan, J.; Chen, H. Comparative transcriptome analysis reveals potential fruiting body formation mechanisms in Morchella importuna. AMB Express 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mirshekari, M.; Einali, A.; Jafar Valizadeh, J. Metabolic changes and activity pattern of antioxidant enzymes induced by salicylic acid treatment in green microalga Dunaliella salina under nitrogen deficiency. J. Appl. Phycol. 2019, 31, 1709–1719. [Google Scholar] [CrossRef]
- Chen, H.; Hao, H.B.; Han, C.C.; Wan, G.H.; Wang, Q.; Chen, M.J.; Juan, J.X.; Feng, Z.Y.; Zhang, J.J. Exogenous L-ascorbic acid regulates the antioxidant system to increase the regeneration of damaged mycelia and induce the development of fruiting bodies in Hypsizygus marmoreus. Fungal Biol. 2020, 124, 551–561. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).