

Priming Watermelon Resistance by Activating Physiological Response and Defense Gene Expression to Alleviate Fusarium Wilt in Wheat-Watermelon Intercropping

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Design
2.2. Plant Growth Analysis
2.3. Assessment of Disease Incidence
2.4. Soil DNA Extraction and qPCR Amplification
2.5. Total RNA Extraction and Gene Expression Analysis
2.6. Defensive Enzyme Assays
2.7. Determination of Total Phenolic and Flavonoid Contents
2.8. Determination of MDA and H2O2 Contents
2.9. Statistical Analysis
3. Results
3.1. Effects of Wheat and Watermelon Root Exudates on Mycelial Growth of FON
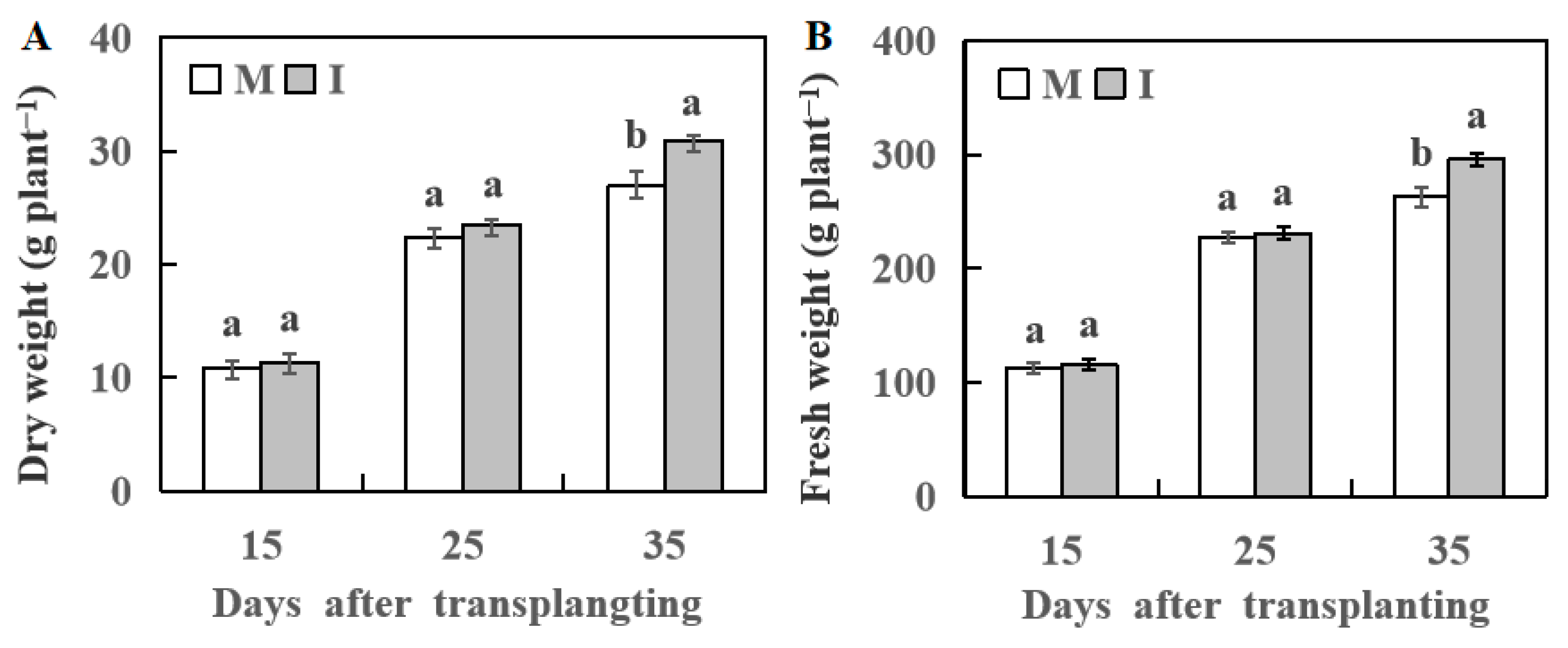
3.2. Effect of Wheat Intercropping on Plant Growth of Watermelon
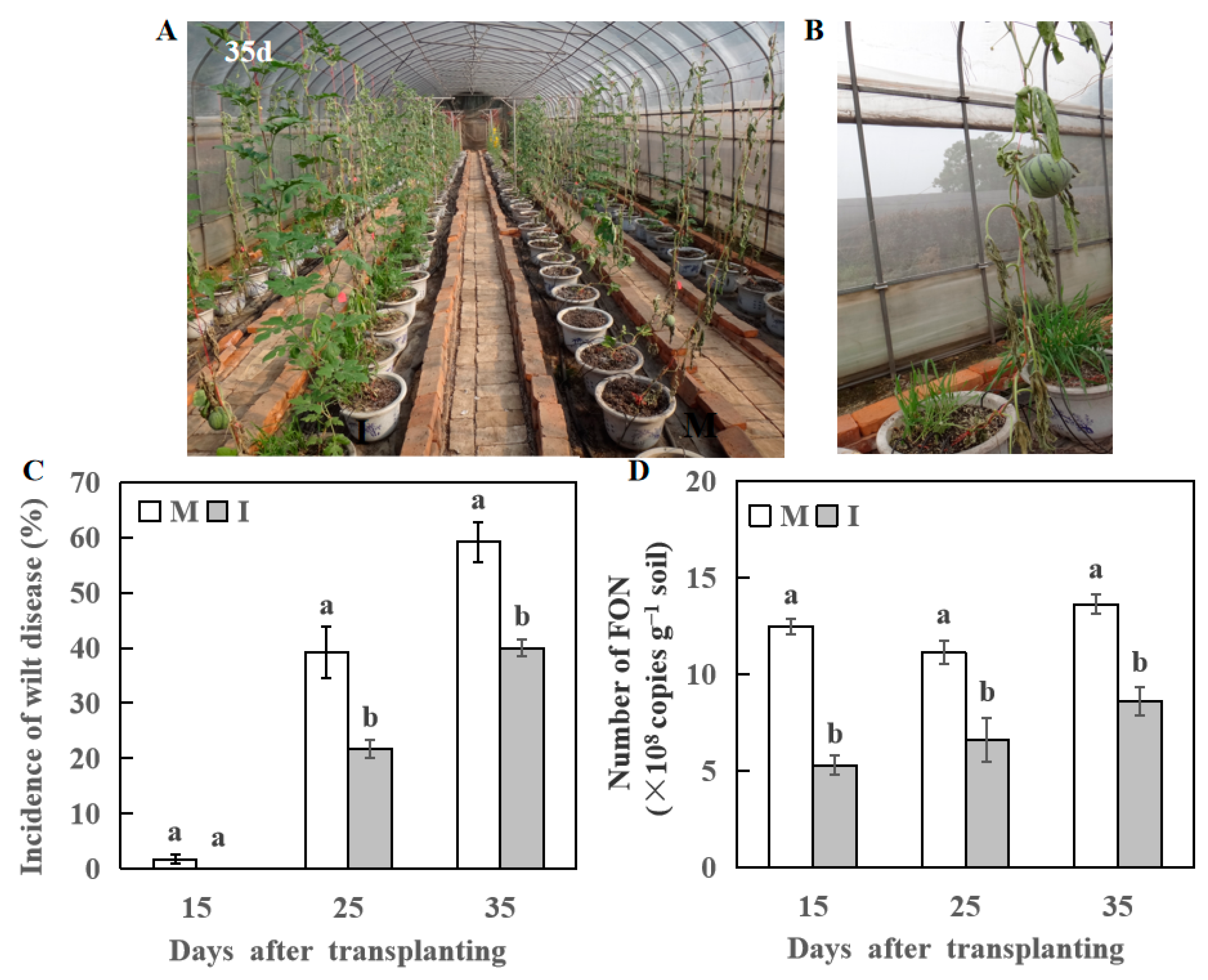
3.3. Effect of Wheat Intercropping on Incidence Rate of Fusarium Wilt of Watermelon
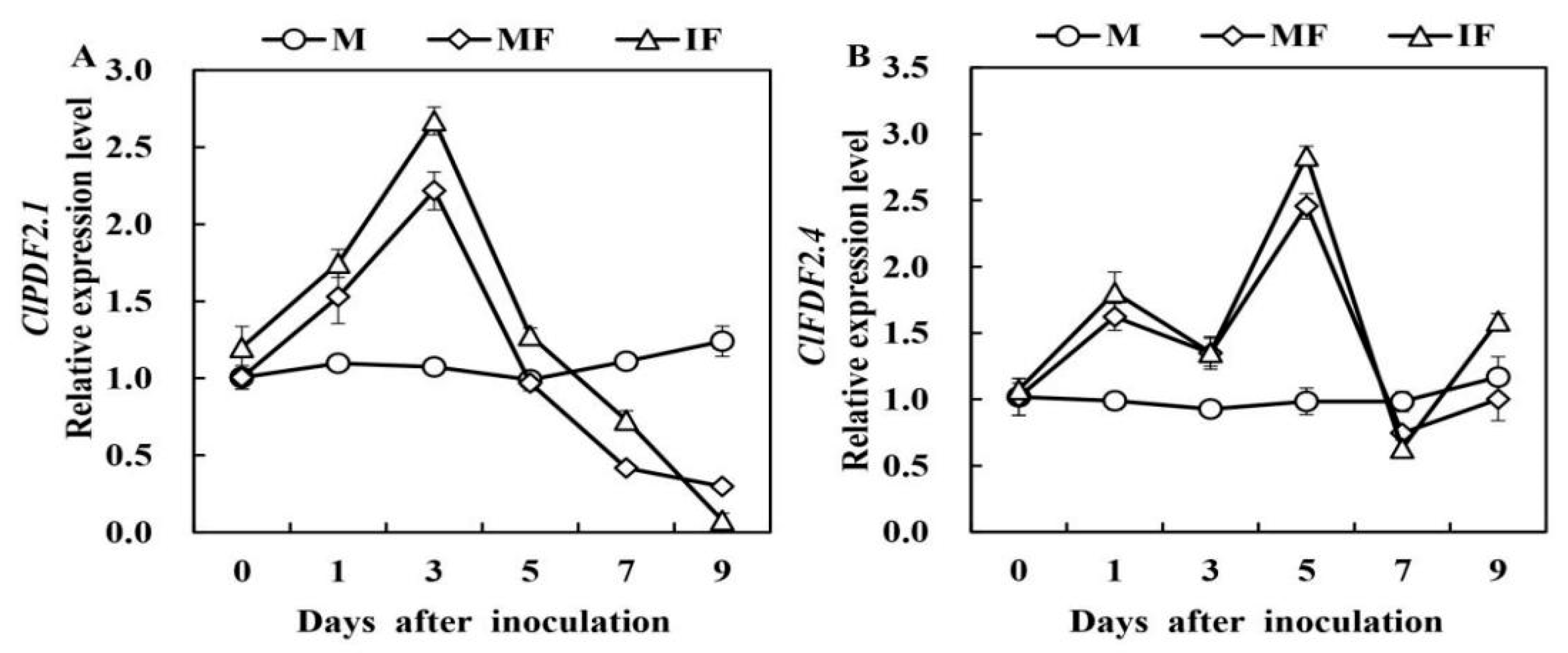
3.4. Effect of Wheat Intercropping on the Expression of Defensive Enzyme Genes and Defensin-Like Genes
3.5. Effect of Wheat Intercropping on Defensive Enzyme Activities
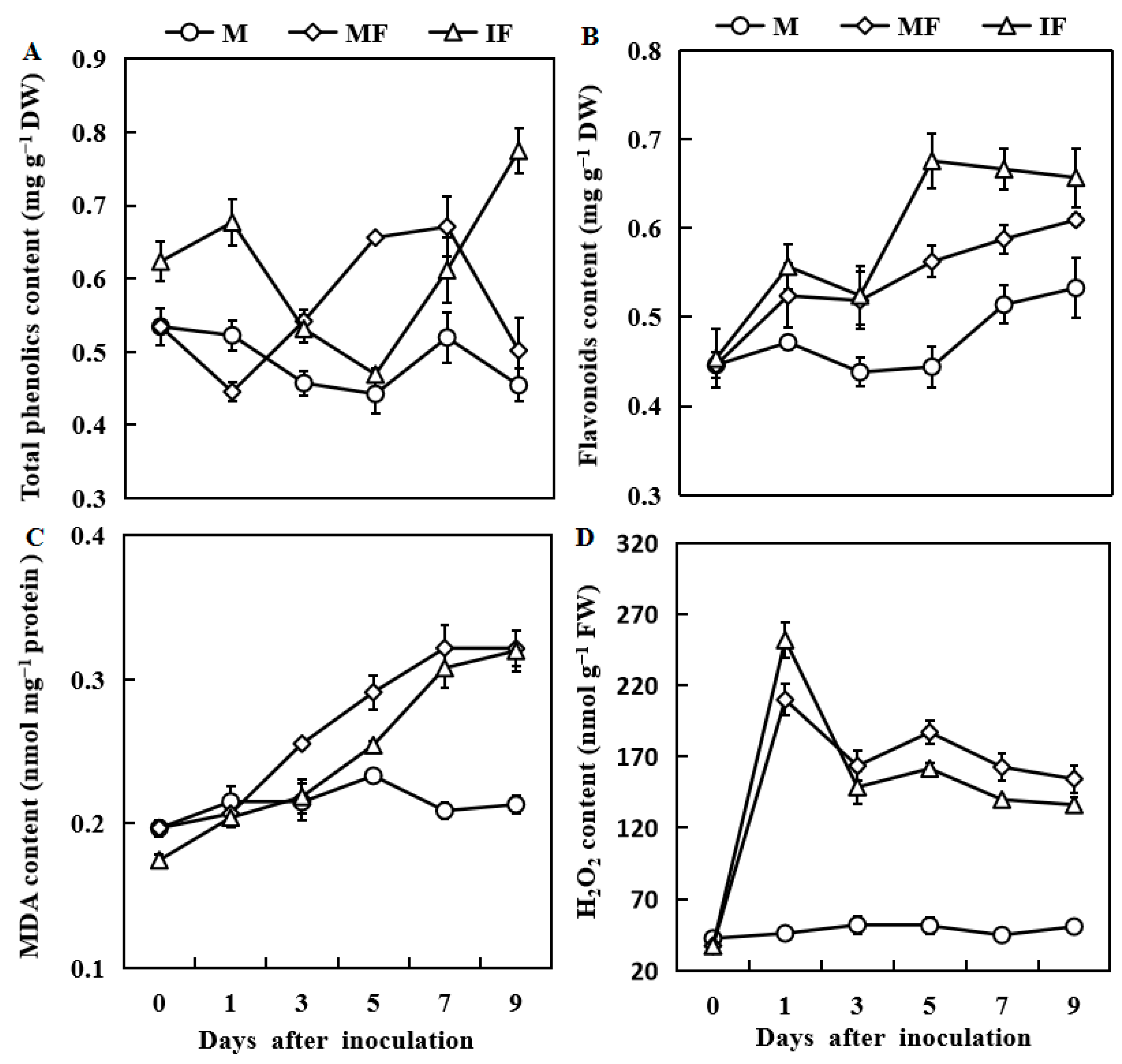
3.6. Effect of Wheat Intercropping on Total Phenolic, Flavonoid, MDA and H2O2 Contents
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ren, L.X.; Su, S.M.; Yang, X.M.; Xu, Y.C.; Huang, Q.W.; Shen, Q.R. Intercropping with aerobic rice suppressed Fusarium wilt in watermelon. Soil. Biol. Biochem. 2008, 40, 834–844. [Google Scholar] [CrossRef]
- Xiao, X.M.; Cheng, Z.H.; Meng, H.W.; Liu, L.H.; Li, H.Z.; Dong, Y.X. Intercropping of green garlic (Allium sativum L.) induces nutrient concentration changes in the soil and plants in continuously cropped cucumber (Cucumis sativus L.) in a plastic tunnel. PLoS ONE 2013, 8, e62173. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.F.; Cao, H.S.; Nawaz, M.A.; Sohail, H.; Huang, Y.; Cheng, F.; Kong, Q.S.; Bie, Z.L. Wheat intercropping enhances the resistance of watermelon to Fusarium wilt. Front. Plant Sci. 2018, 9, 696. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhang, Y.; Qi, L.; Mei, X.; Liao, J.; Ding, X.; Deng, W.; Fan, L.; He, X.; Vivanco, J.M.; et al. Plant-plant-microbe mechanisms involved in soil-borne disease suppressionon a maize and pepper intercropping system. PLoS ONE 2014, 9, e115052. [Google Scholar] [CrossRef]
- Gao, X.; Wu, M.; Xu, R.; Wang, X.; Pan, R.; Kim, H.J.; Liao, H. Root interactions in amaize/soybean intercropping system control soybean soil-borne disease, red crownrot. PLoS ONE 2014, 9, e95031. [Google Scholar]
- Li, X.; Zhang, T.; Wang, X.; Hua, K.; Zhao, L.; Han, Z. The composition of root exudates from two different resistant peanut cultivars and their effects on the growth of soil-borne pathogen. Int. J. Biol. Sci. 2013, 9, 164–173. [Google Scholar] [CrossRef]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell. Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef]
- Ling, N.; Raza, W.; Ma, J.; Huang, Q.; Shen, Q. Identification and role of organic acids in watermelon root exudates for recruiting Paenibacillus polymyxa SQR-21 in the rhizosphere. Eur. J. Soil. Biol. 2011, 47, 374–379. [Google Scholar] [CrossRef]
- Dixon, R.A. Natural products and plant disease resistance. Nature 2001, 411, 843–847. [Google Scholar] [CrossRef]
- D’Auria, J.C.; Gershenzon, J. The secondary metabolism of Arabidopsis thaliana: Growing like a weed. Curr. Opin. Plant Biol. 2005, 8, 308–316. [Google Scholar] [CrossRef]
- Xu, W.H.; Liu, D.; Wu, F.Z.; Liu, S.W. Root exudates of wheat are involved in suppression of Fusarium wilt in watermelon in watermelon-wheat companion cropping. Eur. J. Plant Pathol. 2015, 141, 209–216. [Google Scholar] [CrossRef]
- Morsya, M.R.; Jouve, L.; Hausman, J.F.; Hoffmann, L.; Stewart, J.M. Alteration of oxidative and carbohydrate metabolism under abiotic stress in two rice (Oryza sativa L.) genotypes contrasting in chilling tolerance. J. Plant Physiol. 2007, 164, 157e67. [Google Scholar] [CrossRef] [PubMed]
- Heber, U.; Miyake, C.; Mano, J.; Ohno, C.; Aada, K. Monodehydroascorbate radical detected by electron paramagnetic resonance spectrometry is a sensitive probe of oxidative stress in intact leaves. Plant Cell. Physiol. 1996, 37, 1066–1072. [Google Scholar] [CrossRef]
- Zhao, H.C.; Zhao, H.; Wang, B.C.; Wang, J.B. Effect of local stress induction on resistance-related enzymes in cucumber seeding. Colloid. Surface. B 2005, 43, 37–42. [Google Scholar] [CrossRef]
- Rossum, M.W.P.C.; Alberda, M.; Plas, L.H.W. Role of oxidative damage in tulip bulb scale micropropagation. Plant Sci. 1997, 130, 207–216. [Google Scholar] [CrossRef]
- Bian, S.M.; Jiang, Y.W. Reactive oxygen species, antioxidant enzyme and gene expression patterns in leaves and roots of Kentucky bluegrass in response to drought stress and recovery. Sci. Hortic. 2009, 120, 264–270. [Google Scholar] [CrossRef]
- Liu, C.H.; Zeng, H.L.; He, L.; Ye, P.S.; Wei, S.G.; Zhang, Q.F.; Li, Q.Y. Effect of Trichoderma harzianum T23 on defence enzymes in leaves of eggplants. Southwest. China J. Agri. Sci. 2014, 27, 1945–1948. [Google Scholar]
- Bali, S.G.; Kaur, P.; Jamwal, V.L.; Gandhi, S.G.; Sharma, A.; Ohri, P.; Bhardwaj, R.; Ali, M.A.; Ahmad, P. Seed priming with jasmonic acid counteracts root knot nematode infection in tomato by modulating the activity and expression of antioxidative enzymes. Biomolecules 2020, 10, 98. [Google Scholar] [CrossRef] [PubMed]
- Shigeoka, S.; Ishikawa, T.; Tamoi, M.; Miyagawa, Y.; Takeda, T.; Yabuta, Y.; Yoshimura, K. Regulation and function of ascorbate peroxidase isoenzymes. J. Exp. Bot. 2002, 53, 1305–1319. [Google Scholar] [CrossRef]
- Zhang, F.L.; Ge, H.L.; Zhang, F.; Guo, N.; Wang, Y.C.; Chen, L.; Li, C.W. Biocontrol potential of Trichoderma harzianum isolate T-aloe against Sclerotinia sclerotiorum in soybean. Plant Physiol. Biochem. 2016, 100, 64–74. [Google Scholar] [CrossRef]
- Hao, W.Y.; Ren, L.X.; Ran, W.; Shen, Q.R. Allelopathic effects of root exudates from watermelon and rice plants on Fusarium oxysporum f.sp. niveum. Plant Soil. 2010, 336, 485–497. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.S. The water culture method for growing plants without soil. Calif. Agric. Exp. Stat. Circ. 1950, 347, 1–32. [Google Scholar]
- Wu, H.S.; Yang, X.N.; Fan, J.Q.; Miao, W.G.; Ling, N.; Xu, Y.C.; Huang, Q.W.; Shen, Q.R. Suppression of Fusarium wilt of watermelon by a bio-organic fertilizer containing combinations of antagonistic microorganisms. Biocontrol. 2009, 54, 287–300. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, L. The WRKY transcription factor superfamily: Its origin in eukaryotes and expansion in plants. BMC Evol. Biol. 2005, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.G.; Wu, F.Z. Dynamics of the diversity of fungal and Fusarium communities during continuous cropping of cucumber in the greenhouse. FEMS Microbiol. Ecol. 2012, 80, 469–478. [Google Scholar] [CrossRef]
- Wakelin, S.A.; Warren, R.A.; Kong, L.; Harvey, P.R. Management factors affffecting size and structure of soil Fusarium communities under irrigated maize in Australia. Appl. Soil. Ecol. 2008, 39, 201–209. [Google Scholar] [CrossRef]
- Cheng, F.; Lu, J.Y.; Gao, M.; Shi, K.; Kong, Q.S.; Huang, Y.; Bie, Z.L. Redox signaling and CBF-responsive pathway are involved in salicylic acid-improved photosynthesis and growth under chilling stress in watermelon. Front. Plant Sci. 2016, 7, 1519. [Google Scholar] [CrossRef]
- Kong, Q.S.; Yuan, J.X.; Gao, L.Y.; Zhao, S.; Jiang, W.; Huang, Y.; Bie, Z.L. Identifification of suitable reference genes for gene expression normalization in qRT-PCR analysis in watermelon. PLoS ONE 2014, 9, e90612. [Google Scholar]
- Zhang, M.; Xu, J.H.; Liu, G.; Yao, X.F.; Li, P.F.; Yang, X.P. Characterization of the watermelon seeding infection process by Fusarium oxysporum f. sp. niveum. Plant Pathol. 2015, 64, 1076–1084. [Google Scholar] [CrossRef]
- Livak, K.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2−11 CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Patra, H.K.; Kar, M.; Mishra, D. Catalase activity in leaves and cotyledons during plant development and senescence. Biochem. Physiol. Pflanz. 1978, 172, 385–390. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence:correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp.Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbatespecific peroxidase in spinach chloroplasts. Plant Cell. Physiol. 1981, 22, 867–880. [Google Scholar]
- Wang, Y.S.; Tian, S.P.; Xu, Y.; Qin, G.Z.; Yao, H.J. Changes in the activities of pro-and anti-oxidant enzymes in peach fruit inoculated with Cryptococcus laurentii or Penicillium expansum at 0 or 20 °C. Postharvest Biol. Technol. 2004, 34, 21–28. [Google Scholar] [CrossRef]
- Lagrimini, L.M.; Rothstein, S. Tissue specificity of tobacco peroxidase isozymes and their induction by wounding and tobacco mosaic virus infection. Plant Physiol. 1987, 84, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Arnaldos, T.L.; Muñoz, R.; Ferrer, M.A.; Calderón, A.A. Changes in phenol content during strawberry (Frabaria annassa cv. Chandler) callus culture. Plant Physiol. 2001, 113, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.M.; Garcia, P.C.; Rivero, R.M.; Romero, L. Response of phenolic metabolism to the application of carbendazin plus boron in tobacco. Plant Physiol. Biochem. 1999, 106, 151–157. [Google Scholar]
- Tekel’ova, D.; Repcak, M.; Zemkova, E.; Toth, J. Quantitative changes of dianthrones, hyperforin and favonoids content in the flower ontogenesis of Hypericum perforatum. Planta Med. 2000, 66, 778–780. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Willekens, H.; Chamnongpol, S.; Davey, M.; Schraudner, M.; Langebartels, C.; Montagu, M.V.; Inzé, D.; Camp, W.V. Catalase is a sink for H2O2 and is indispensable for stress defence in C3 plants. EMBO J. 1997, 16, 4806. [Google Scholar] [CrossRef]
- Zhou, X.G.; Everts, K.L. Suppression of Fusarium wilt of watermelon by soil amendment with hairy vetch. Plant Dis. 2004, 88, 1357–1365. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.H.; Wang, Z.G.; Wu, F.Z. Companion cropping with wheat increases resistance to Fusarium wilt in watermelon and the roles of root exudates in watermelon root growth. Physiol. Mol. Plant Pathol. 2015, 90, 12–20. [Google Scholar] [CrossRef]
- Ling, N.; Zhang, W.; Wang, D.; Mao, J.; Huang, Q.; Guo, S.; Shen, Q.R. Root exudates from grafted-root watermelon showed a certain contribution in inhibiting Fusarium oxysporum f.sp. niveum. PLoS ONE 2013, 8, e63383. [Google Scholar]
- Wu, X.; Wu, F.Z.; Zhou, X.G.; Fu, X.P.; Tao, Y.; Xu, W.H.; Pan, K.; Liu, S.W. Effects of Intercropping with Potato Onion on the Growth of Tomato and Rhizosphere Alkaline Phosphatase Genes Diversity. Front. Plant Sci. 2016, 7, 846. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Jeond, S.; Lee, C. Antioxidant capacity of phenolic phytochemicals from various cultivars of plums. Food. Chem. 2003, 81, 321e6. [Google Scholar] [CrossRef]
- Li, C.X.; Fu, X.P.; Zhou, X.G.; Liu, S.W.; Xia, Y.; Li, N.H.; Zhang, X.X.; Wu, F.Z. Treatment with Wheat Root Exudates and Soil Microorganisms from Wheat/Watermelon Companion Cropping Can Induce Watermelon Disease Resistance against Fusarium oxysporum f. sp. niveum. Plant Dis. 2019, 103, 1693–1702. [Google Scholar] [CrossRef]
- Zvirin, T.; Herman, R.; Brotman, Y. Differential colonization and defence responses of resistant and susceptible melon lines infected by Fusarium oxysporum race 1.2. Plant Pathol. 2010, 59, 576–585. [Google Scholar] [CrossRef]
- Broglie, K.; Chet, I.; Holliday, M.; Cressman, R.; Biddle, P.; Knowlton, S.; Mauvais, C.J.; Broglie, R. Transgenic plants with enhanced resistance to the fungal pathogen Rhizoctonia solani. Science 1991, 254, 1194–1197. [Google Scholar] [CrossRef]
- Kaur, N.; Chen, W.B.; Zheng, Y.; Hasegawa, D.K.; Ling, K.S.; Fei, Z.J.; Wintermantel, W.M. Transcriptome analysis of the white fly, Bemisia tabaci MEAM1 during feeding on tomato infected with the crinivirus, Tomato chlorosis virus, identi-fies a temporal shift in gene expression and differential reg-ulation of novel orphan genes. BMC Genom. 2017, 18, 370. [Google Scholar] [CrossRef]
- Mittler, R.; Herr, E.H.; Orvar, B.L.; Van Camp, W.; Willekens, H.; Inzé, D.; Ellis, B.E. Transgenic tobacco plants with reduced capability to detoxify reactive oxygen intermediates are hyperresponsive to pathogen infection. Proc. Natl. Acad. Sci. USA 1999, 96, 14165–14170. [Google Scholar] [CrossRef]
- Li, L.; Steffens, J.C. Overexpression of polyphenol oxidase in transgenic tomato plants results in enhanced bacterial disease resistance. Planta 2002, 215, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Gechev, T.; Willekens, H.; Montagu, M.V.; Inzé, D.; Camp, W.V.; Toneva, V.; Minkov, I. Different responses of tobacco antioxidant enzymes to light and chilling stress. J. Plant Physiol. 2003, 160, 509e15. [Google Scholar] [CrossRef] [PubMed]
- De Ascensao, A.R.; Dubery, I.A. Panama disease: Cell wall reinforcement in banana roots in response to elicitors from Fusarium oxysporum f. sp. cubense race four. Phytopathology 2000, 90, 1173–1180. [Google Scholar] [CrossRef] [PubMed]
- Kavino, M.; Kumar, N.; Damodaran, T.; Harish, S.; Saravanakumar, D. Biochemical markers as a useful tool for the early identifification of Fusarium oxysporum f.sp. cubense, race 1 resistance banana clones. Arch. Phytopathol. Plant Prot. 2009, 42, 1069–1078. [Google Scholar] [CrossRef]
- Wu, Y.L.; Yi, G.J.; Peng, X.X.; Huang, B.Z.; Liu, E.E.; Zhang, J.J. Systemic acquired resistance in Cavendish banana induced by infection with an incompatible strain of Fusarium oxysporum f. sp. cubense. J. Plant Physiol. 2013, 170, 1039–1046. [Google Scholar] [CrossRef]
- Hüttermann, A.; Mai, C.; Kharazipour, A. Modifification of lignin for the production of new compounded materials. Appl. Microbiol. Biotechnol. 2001, 55, 387–394. [Google Scholar]
- Sudhamoy, M.; Adinpunya, M. Reinforcement of cell wall in roots of Lycopersicon esculentum through induction of phenolic compounds and lignin by elicitors. Physiol. Mol. Plant Pathol. 2007, 71, 201–209. [Google Scholar]
- Cushnie, T.P.; Lamb, A.J. Recent Advances in Understanding the Antibacterial Properties of Flavonoids. Int. J. Antimicrob. Agents 2011, 38, 99–107. [Google Scholar] [CrossRef]
- Rosell, G.; Quero, C.; Coll, J.; Guerrero, A. Biorational insecticides in pest management. J. Pestic. Sci. 2008, 33, 103–121. [Google Scholar] [CrossRef]
- Mandavia, M.K.; Gajera, H.P.; Andharia, J.H.; Khandar, R.R.; Parameswaran, M. Cell wall degrading enzymes in host pathogen interaction of Fusarium wilt of chickpea: Inhibitory effect of phenolic compounds. Indian Phytopathol. 1999, 52, 285–288. [Google Scholar]
- Bahraminejad, S.; Asenstorfer, R.E.; Riley, I.T.; Schutz, C.J. Analysis of antimicrobial activity of flflavonoids and saponins isolated from the shoots of oats (Avena sativa L.). J. Phytopathol. 2008, 156, 1–7. [Google Scholar]
- Zhou, Y.; Dahler, J.M.; Underhill, S.J.R.; Wills, R.B.H. Enzymes associated with blackheart development in pineapple fruit. Food Chem. 2003, 80, 565–572. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| ClCAC | 5′-AATTGTGGTTGATGCTGCAC-3′ | 5′-TGACAGCTGTACCTGGCATC-3′ |
| ClTUA | 5′-CTTGCTGGGAGCTCTATTGC-3′ | 5′-AACGGATTAAAAGCGTCGTG-3′ |
| ClAPX | 5′-CACCCTGGTAGAGAGGACAAAC-3′ | 5′-TCAAAGATGAGAGGGTTGGAAG-3′ |
| ClPPO | 5′-GCAAGAAGGAGAAAGAGGATGA-3′ | 5′-CTTCAAGCAATTCGGTTATTCC-3′ |
| ClPOD | 5′-AGTGGGTGGGTTGACCTTTCT-3′ | 5′-ATCACAAAGGGCTTCTCCAAA-3′ |
| ClCAT | 5′-GCTCACCATGCCGAGAGGTATC-3′ | 5′-CGTTCTTGCCTGTCTGATGTCC-3′ |
| ClSOD | 5′-TCTCAAGCTACCTCGCCACTC-3′ | 5′-AGCGTGACGACGCCTTCAAC-3′ |
| ClCHI | 5′-CTGAATTCTTGGAGTCAGTGGA-3′ | 5′-ACGCCTTGCTCCATAACATAAC-3′ |
| ClPDF2.1 | 5′-ATGAAGTTCTTTTCCGCTGC-3′ | 5′-TCAAACGCAGTGCTTTGTGCAGAAG-3′ |
| ClPDF2.4 | 5′-ATGAAGTTTCTTTTTCAGCTGC-3′ | 5′-TCAAACGCAGTGCTTTGTG-3′ |
| Cultivars | Colony Growth Diameters of FON (cm) | |||
|---|---|---|---|---|
| 0.5 mL Plant−1 | 1 mL Plant−1 | 5 mL Plant−1 | 10 mL Plant−1 | |
| Control | 3.34 ± 0.044 b | 3.34 ± 0.044 b | 3.34 ± 0.044 b | 3.34 ± 0.044 b |
| E18 | 3.02 ± 0.026 c | 3.04 ± 0.018 d | 3.04 ± 0.009 d | 3.12 ± 0.040 c |
| E352 | 3.06 ± 0.029 c | 3.10 ± 0.004 cd | 3.14 ± 0.041 c | 3.16 ± 0.027 c |
| Z9023 | 3.09 ± 0.016 c | 3.15 ± 0.015 c | 3.18 ± 0.017 c | 3.18 ± 0.010 c |
| Watermelon | 3.63 ± 0.012 a | 3.53 ± 0.039 a | 3.46 ± 0.031 a | 3.44 ± 0.018 a |
| Treatment | Days | Length (cm) | SurfArea (cm2) | Volume (cm3) | AvgDiam (mm) | Number | Dry Weight (g) |
|---|---|---|---|---|---|---|---|
| M | 15 | 863.14 ± 40.25 a | 123.49 ± 1.75 a | 1.43 ± 0.06 a | 0.36 ± 0.02 a | 4916.00 ± 55.64 a | 0.17 ± 0.01 a |
| I | 977.77 ± 13.77 a | 154.02 ± 12.24 a | 1.68 ± 0.07 a | 0.39 ± 0.01 a | 5638.33 ± 262.76 a | 0.18 ± 0.01 a | |
| M | 25 | 1786.3 ± 27.19 a | 241.38 ± 7.32 b | 2.66 ± 0.10 b | 0.49 ± 0.01 b | 9496.33 ± 237.45 b | 0.28 ± 0.01 a |
| I | 1900.97 ± 46.07 a | 270.76 ± 6.43 a | 3.16 ± 0.09 a | 0.54 ± 0.01 a | 10425.67 ± 59.53 a | 0.31 ± 0.01 a | |
| M | 35 | 2190.48 ± 31.17 b | 315.39 ± 18.34 b | 3.51 ± 0.15 b | 0.55 ± 0.01 a | 13428.33 ± 252.2 b | 0.54 ± 0.02 b |
| I | 2451.63 ± 53.57 a | 390.05 ± 15.44 a | 5.09 ± 0.42 a | 0.60 ± 0.01 a | 14295.00 ± 117.41 a | 0.60 ± 0.01 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, H.; Lu, J.; Huang, Y.; Wang, M.; Yan, C.; Bie, Z. Priming Watermelon Resistance by Activating Physiological Response and Defense Gene Expression to Alleviate Fusarium Wilt in Wheat-Watermelon Intercropping. Horticulturae 2023, 9, 27. https://doi.org/10.3390/horticulturae9010027
Lv H, Lu J, Huang Y, Wang M, Yan C, Bie Z. Priming Watermelon Resistance by Activating Physiological Response and Defense Gene Expression to Alleviate Fusarium Wilt in Wheat-Watermelon Intercropping. Horticulturae. 2023; 9(1):27. https://doi.org/10.3390/horticulturae9010027
Chicago/Turabian StyleLv, Huifang, Junyang Lu, Yuan Huang, Mingxia Wang, Congsheng Yan, and Zhilong Bie. 2023. "Priming Watermelon Resistance by Activating Physiological Response and Defense Gene Expression to Alleviate Fusarium Wilt in Wheat-Watermelon Intercropping" Horticulturae 9, no. 1: 27. https://doi.org/10.3390/horticulturae9010027
APA StyleLv, H., Lu, J., Huang, Y., Wang, M., Yan, C., & Bie, Z. (2023). Priming Watermelon Resistance by Activating Physiological Response and Defense Gene Expression to Alleviate Fusarium Wilt in Wheat-Watermelon Intercropping. Horticulturae, 9(1), 27. https://doi.org/10.3390/horticulturae9010027