
OfSPL11 Gene from Osmanthus fragrans Promotes Plant Growth and Oxidative Damage Reduction to Enhance Salt Tolerance in Arabidopsis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Salt Treatment
2.2. Gene Cloning and Bioinformatics Analysis
2.3. Subcellular Localization and Transcriptional Autoactivation
2.4. Transient Transformation in Leaves of Tobacco and Callus of O. fragrans
2.5. Generation of Transgenic Arabidopsis Plants
2.6. Salt Tolerance Assay
2.7. Measurement of Plant Physiological Index
2.8. Yeast One-Hybrid Assays
2.9. Transient Expression Assay
2.10. Quantitative Real-Time PCR and Statistical Analysis
3. Results
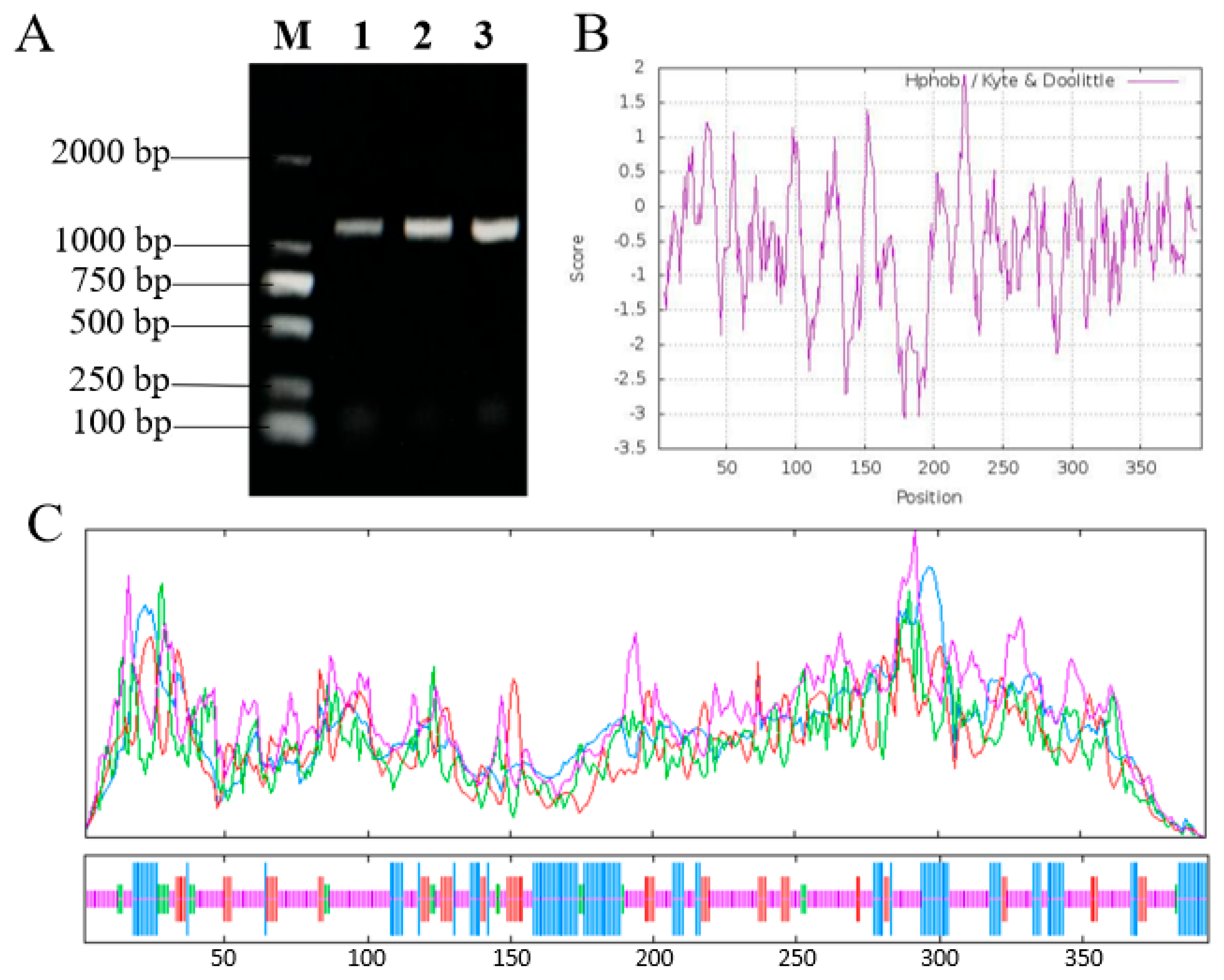
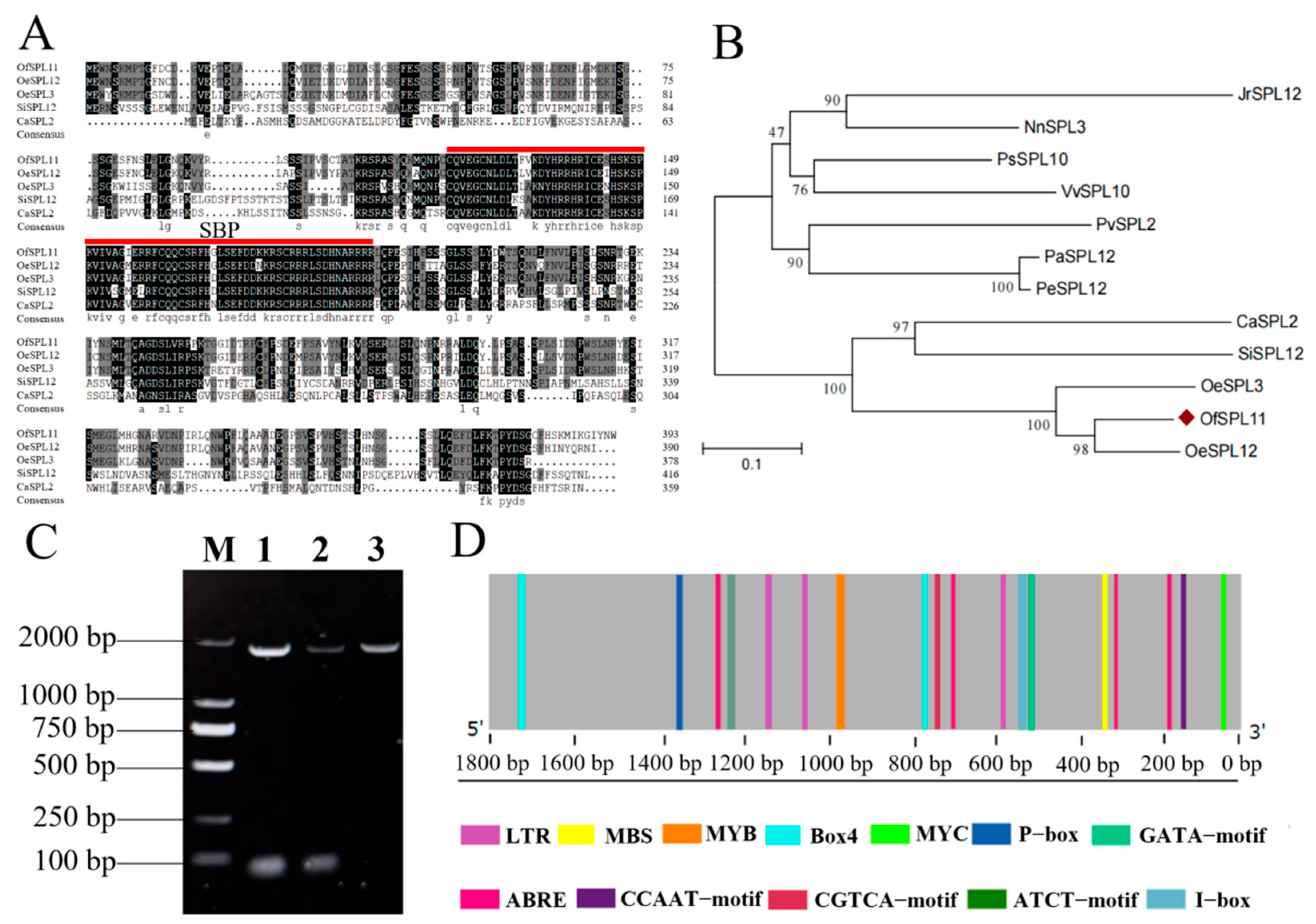
3.1. OfSPL11 Clone and Sequence Analysis
3.2. Cis-Acting Element Analysis of OfSPL11 Promoter
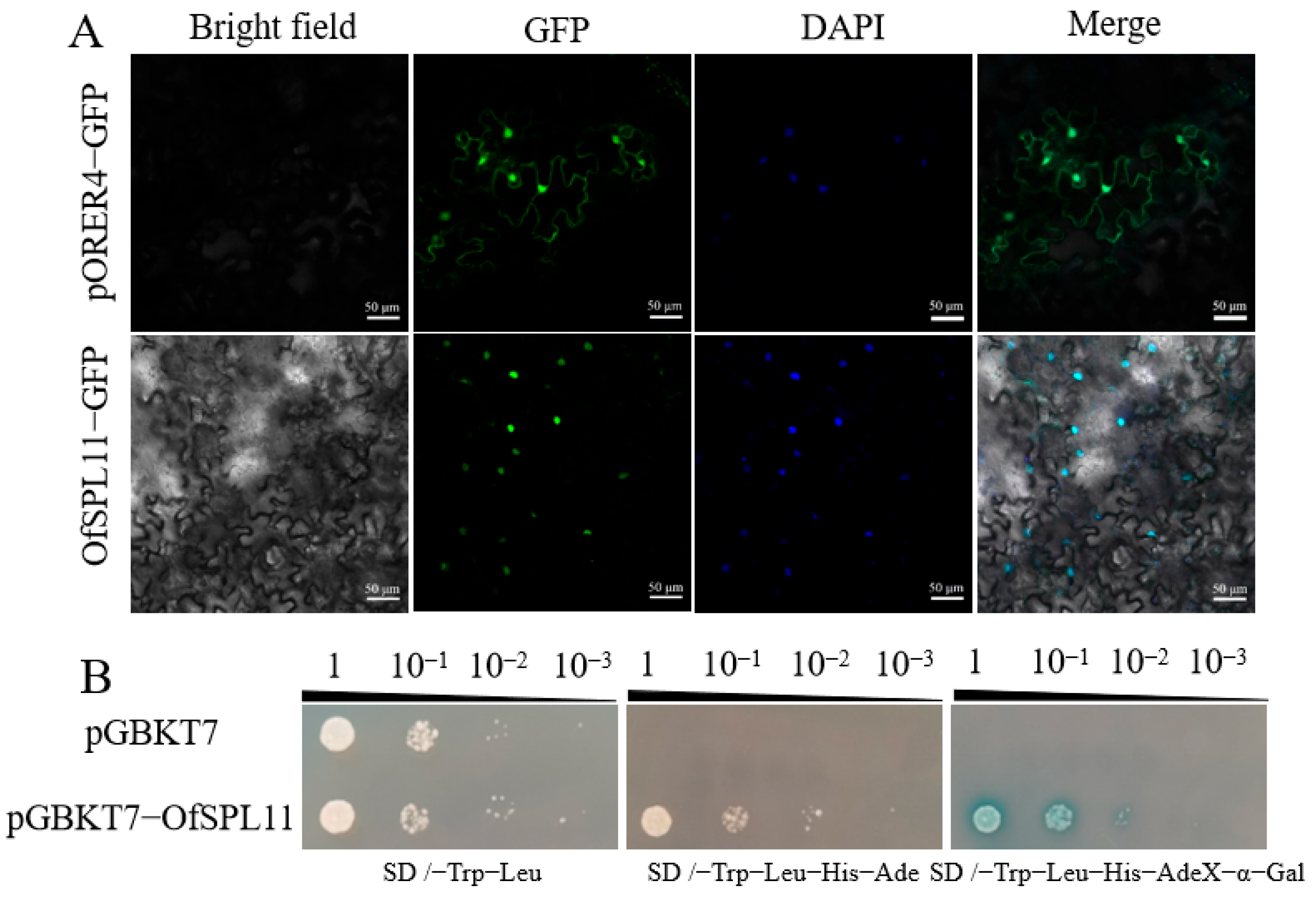
3.3. Subcellular Localization and Transcriptional Autoactivation
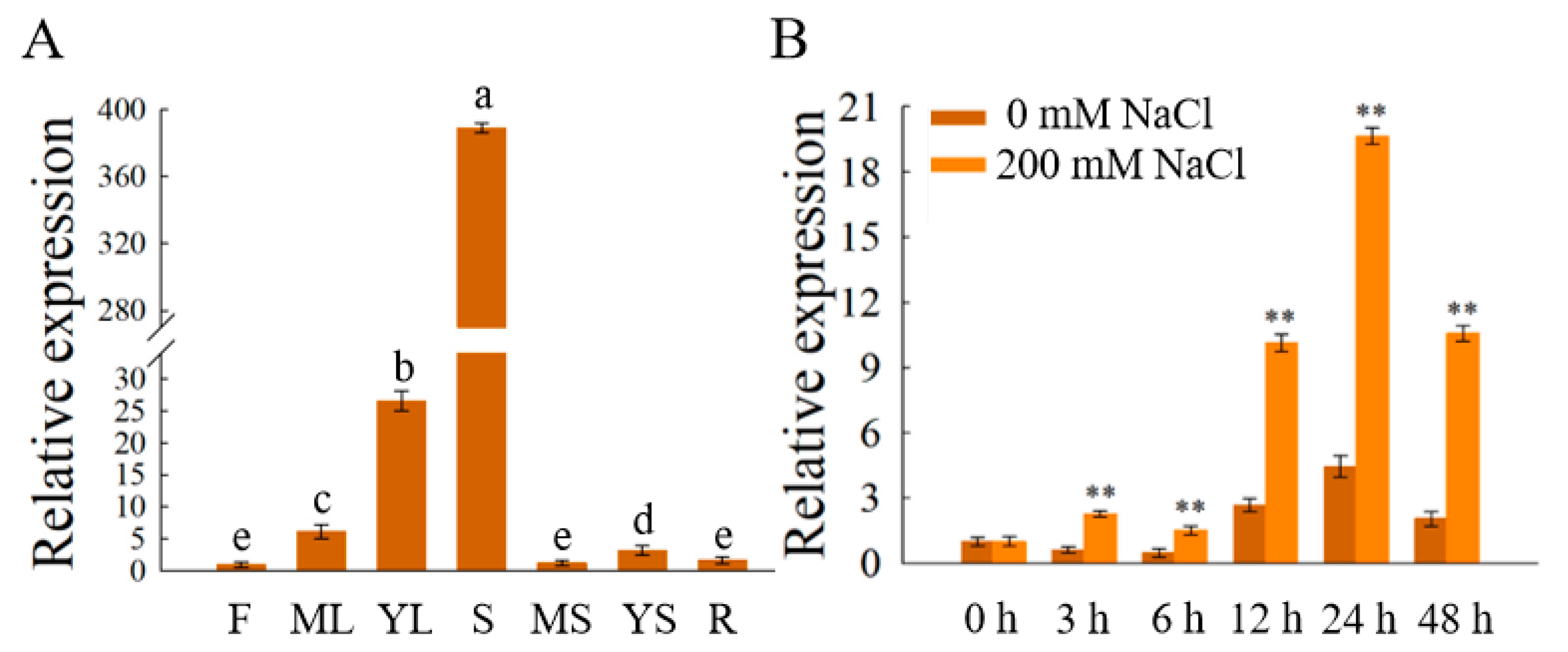
3.4. Tissue and Salt Stress Expression Profiling of OfSPL11
3.5. The Promoter of OfSPL11 Was Sensitive to Salinity
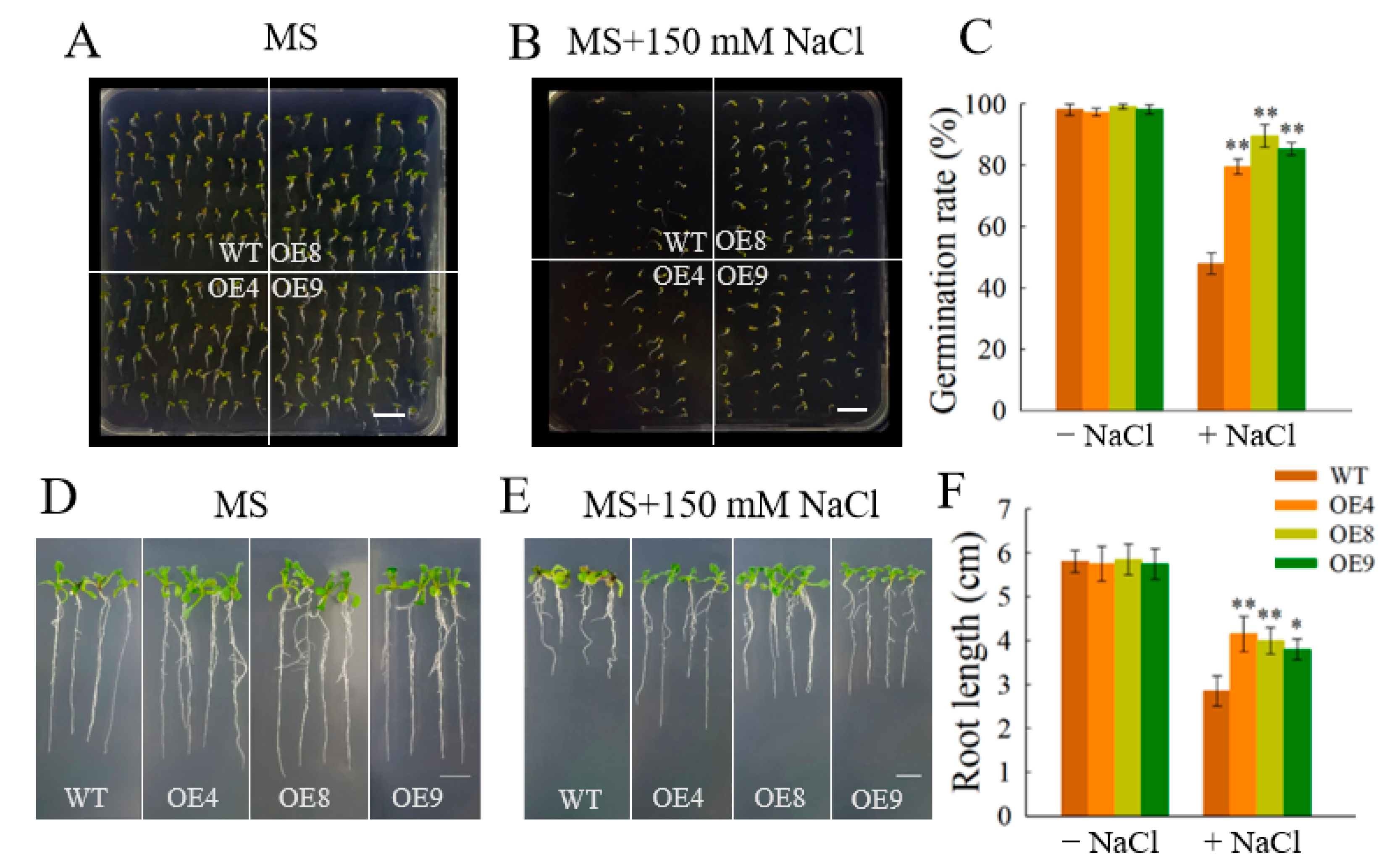
3.6. Phenotypic Characteristics of Arabidopsis Lines Overexpressing OfSPL11
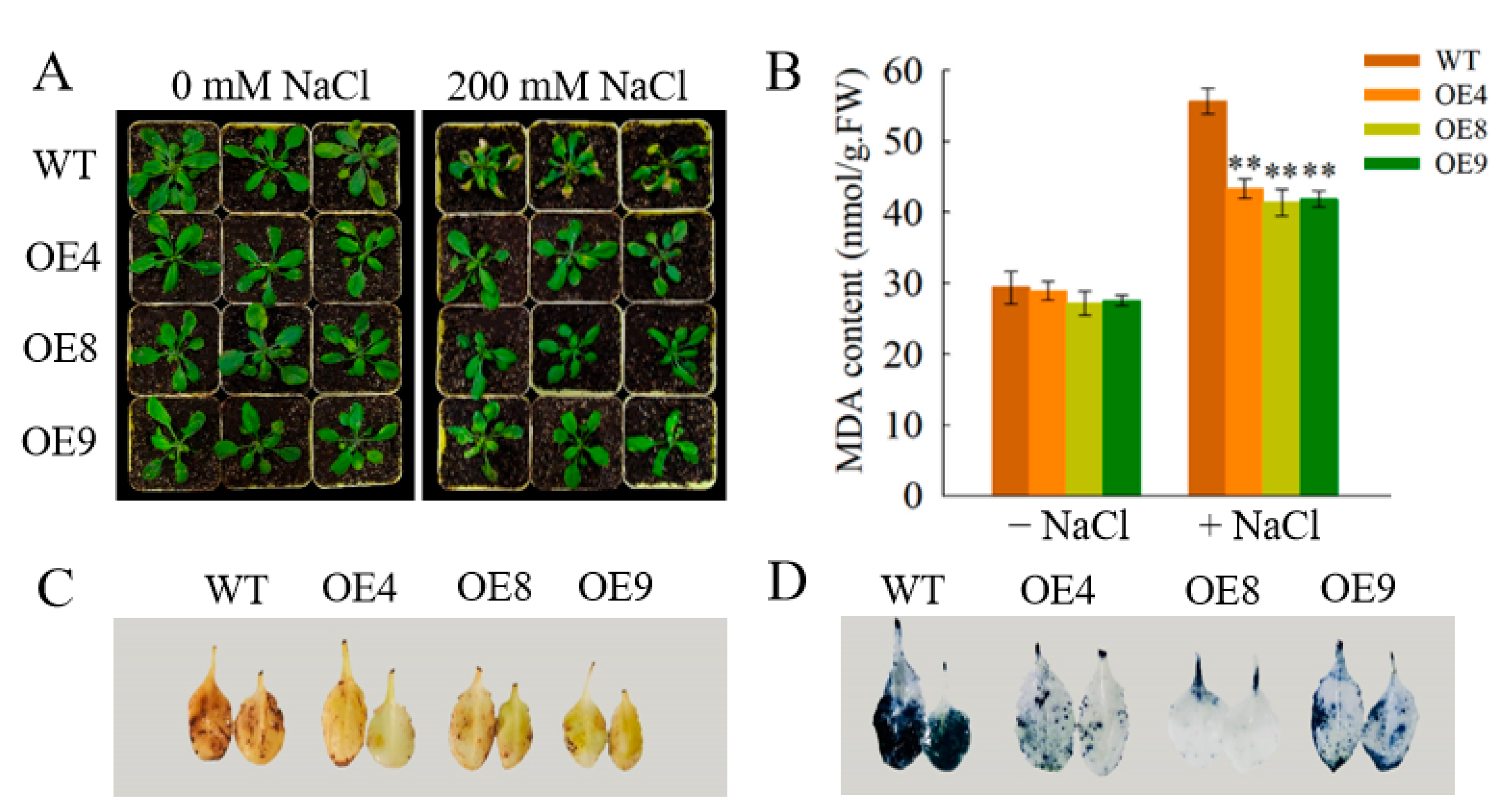
3.7. OfSPL11 Enhances Salt Tolerance in Arabidopsis
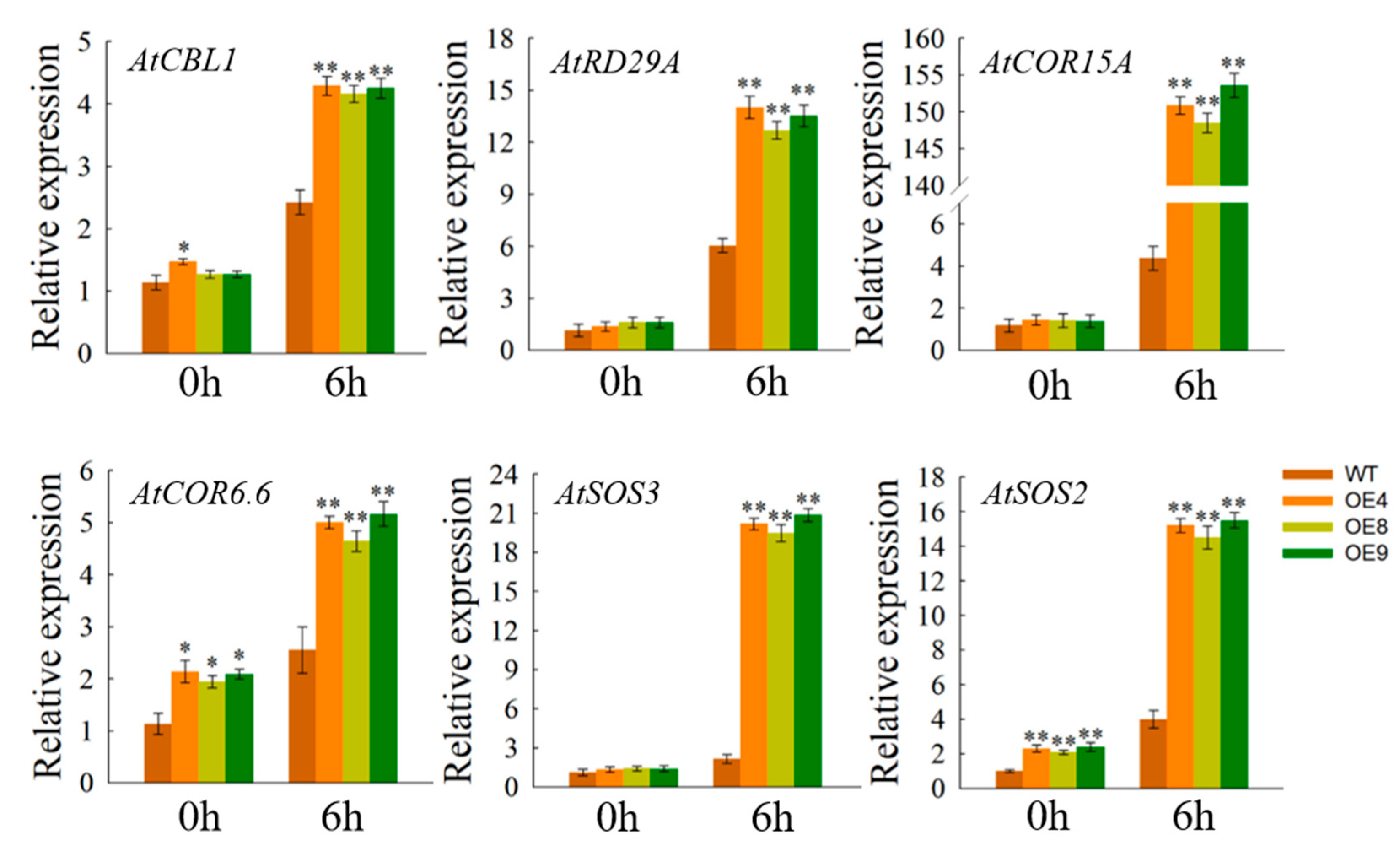
3.8. Expression of Abiotic Stress Response Genes in OfSPL11 Overexpression Lines
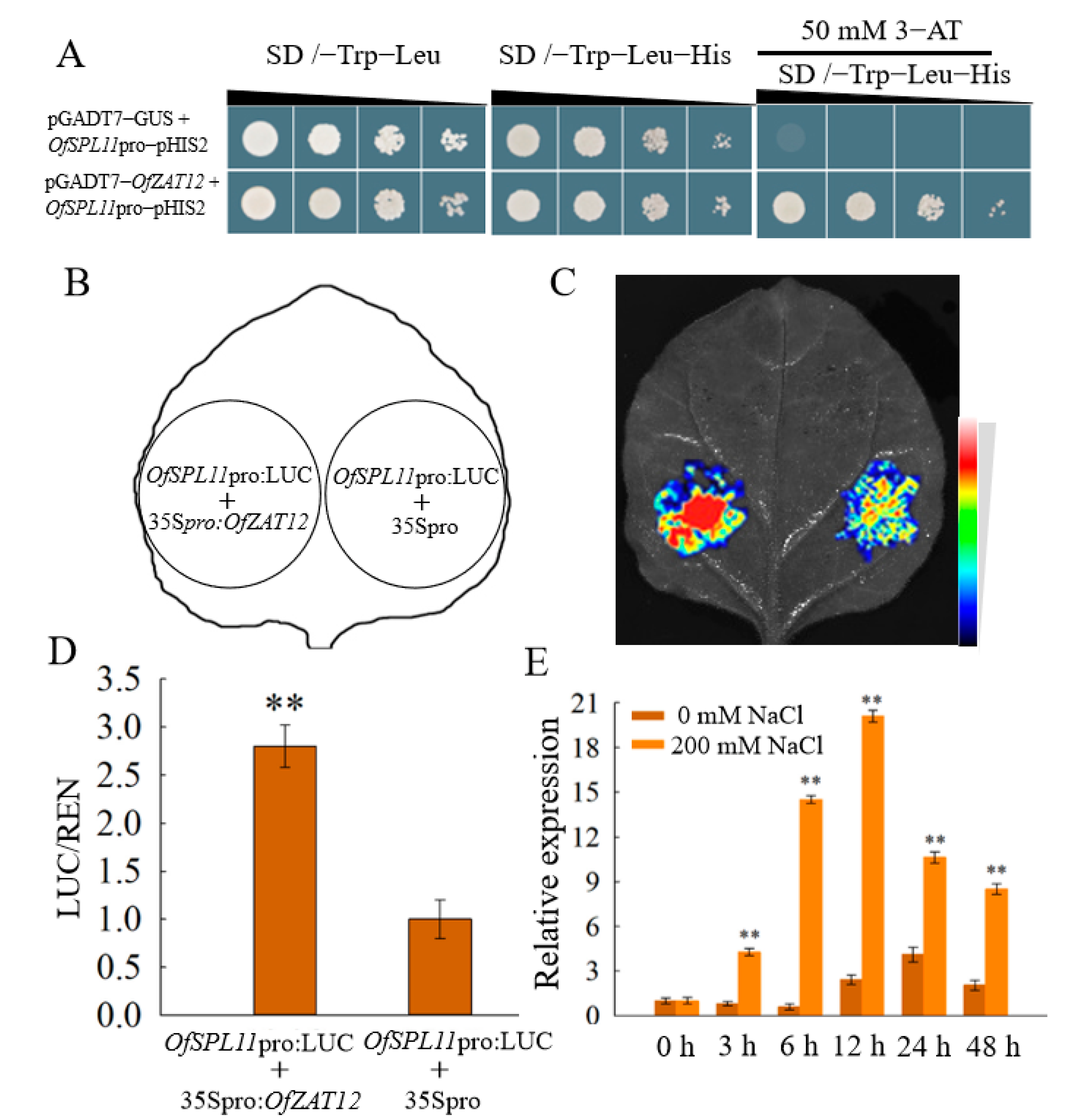
3.9. OfZAT12 Regulated the Expression of OfSPL11
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kong, C.C.; Ren, C.G.; Li, R.Z.; Xie, Z.H.; Wang, J.P. Hydrogen Peroxide and Strigolactones Signaling Are Involved in Alleviation of Salt Stress Induced by Arbuscular Mycorrhizal Fungus in Sesbania cannabina Seedlings. J. Plant Growth Regul. 2017, 36, 734–742. [Google Scholar] [CrossRef]
- Moles, T.M.; Pompeiano, A.; Huarancca, R.T.; Scartazza, A.; Guglielminetti, L. The efficient physiological strategy of a tomato landrace in response to short-term salinity stress. Plant Physiol. Biochem. 2016, 109, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441–445. [Google Scholar] [CrossRef]
- Chen, M.; Yang, Z.; Liu, J.; Zhu, T.; Wei, X.; Fan, H.; Wang, B. Adaptation Mechanism of Salt Excluders under Saline Conditions and Its Applications. Int. J. Mol. Sci. 2018, 19, 3688. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Liu, J.; Zhu, T.; Zhao, C.; Li, L.; Chen, M. The Role of Melatonin in Salt Stress Responses. Int. J. Mol. Sci. 2019, 20, 1735. [Google Scholar] [CrossRef] [Green Version]
- Zhao, K.F.; Song, J.; Fan, H.; Zhou, S.; Zhao, M. Growth response to ionic and osmotic stress of NaCl in salt-tolerant and salt-sensitive maize. J. Integr. Plant Biol. 2010, 52, 468–475. [Google Scholar] [CrossRef]
- Wu, Y.; Fan, X.; Zhang, Y.; Jiang, J.; Sun, L.; Rahman, F.U.; Liu, C. VvSNAT1 overexpression enhances melatonin production and salt tolerance in transgenic Arabidopsis. Plant Physiol. Biochem. 2021, 166, 485–494. [Google Scholar] [CrossRef]
- Wen, X.; Geng, F.; Cheng, Y.; Wang, J. Ectopic expression of CsMYB30 from Citrus sinensis enhances salt and drought tolerance by regulating wax synthesis in Arabidopsis thaliana. Plant Physiol. Biochem. 2021, 166, 777–788. [Google Scholar] [CrossRef]
- Klein, J.; Saedler, H.; Huijser, P. A new family of DNA binding proteins includes putative transcriptional regulators of the Antirrhinum majus floral meristem identity gene SQUAMOSA. Mol. Gen. Genet. 1996, 250, 7–16. [Google Scholar] [CrossRef]
- Yamasaki, K.; Kigawa, T.; Inoue, M.; Tateno, M.; Yamasaki, T.; Yabuki, T.; Aoki, M.; Seki, E.; Matsuda, T.; Nunokawa, E. A novel zinc-binding motif revealed by solution structures of DNA-binding domains of Arabidopsis SBP-family transcription factors. J. Mol. Biol. 2004, 337, 49–63. [Google Scholar] [CrossRef]
- Moreno, A.M.; Harper, C.L.; Krueger, W.R.; Dellaporta, L.S.; Michael, F. liguleless1 encodes a nuclear-localized protein required.for induction of ligules and auricles during maize leaf organogenesis. Genes Dev. 1997, 11, 616–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preston, J.C.; Hileman, L. Functional evolution in the plant SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE (SPL) gene family. Front. Plant Sci. 2013, 4, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, L.M.; Liu, Y.Q.; Chen, D.Y.; Xue, X.Y.; Mao, Y.B.; Chen, X.Y. Arabidopsis Transcription Factors SPL1 and SPL12 Confer Plant Thermotolerance at Reproductive Stage. Mol. Plant 2017, 10, 735–748. [Google Scholar] [CrossRef] [PubMed]
- Ning, K.; Chen, S.; Huang, H.; Jiang, J.; Yuan, H.; Li, H. Molecular characterization and expression analysis of the SPL gene family with BpSPL9 transgenic lines found to confer tolerance to abiotic stress in Betula platyphylla Suk. Plant Cell Tissue Organ Cult. 2017, 130, 469–481. [Google Scholar] [CrossRef]
- Clough, J.S.; Bent, F.A. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Saleki, R.; Young, G.P.; Lefebvre, D.D. Mutants of Arabidopsis thaliana Capable of Germination under Saline Conditions. Plant Physiol. 1993, 101, 839–845. [Google Scholar] [CrossRef] [Green Version]
- Kotchoni, O.S.; Kuhns, C.; Ditzer, A.; Kirch, H.H.; Bartels, D. Over-expression of different aldehyde dehydrogenase genes in Arabidopsis thaliana confers tolerance to abiotic stress and protects plants against lipid peroxidation and oxidative stress. Plant Cell Environ. 2006, 29, 1033–1048. [Google Scholar] [CrossRef]
- Kim, S.H.; Woo, D.H.; Kim, J.M.; Lee, S.Y.; Chung, W.S.; Moon, Y.H. Arabidopsis MKK4 mediates osmotic-stress response via its regulation of MPK3 activity. Biochem. Biophys. Res. Commun. 2011, 412, 150–154. [Google Scholar] [CrossRef]
- Livak, J.K.; Schmittgen, D.T. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Du, Q.; Tomkinson, E.A.; Gardner, D.P. Transcriptional Regulation of Neuronal Nicotinic Acetylcholine Receptor Genes. J. Biol. Chem. 1997, 272, 14990–14995. [Google Scholar] [CrossRef] [Green Version]
- Arzani, A. Improving salinity tolerance in crop plants: A biotechnological view. In Vitro Cell. Dev. Biol. Plant. 2008, 44, 373–383. [Google Scholar] [CrossRef]
- Cui, L.G.; Shan, J.X.; Shi, M.; Gao, J.P.; Lin, H.X. The miR156-SPL9-DFR pathway coordinates the relationship between development and abiotic stress tolerance in plants. Plant J. 2014, 80, 1108–1117. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M.P.; Bressan, A.R.; Zhu, J.K.; Bohnert, J.H. Plant cellular and molecular responses to high salinity. Plant Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [Green Version]
- Mittova, V.; Tal, M.; Volokita, M.; Guy, M. Salt stress induces up-regulation of an efficient chloroplast antioxidant system in the salt-tolerant wild tomato species Lycopersicon pennellii but not in the cultivated species. Physiol. Plant. 2002, 115, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Wen, D.; Yang, W.; Meng, Q.; Shi, Q.; Gong, B. Overexpression of Caffeic Acid O-Methyltransferase 1 (COMT1) Increases Melatonin Level and Salt Stress Tolerance in Tomato Plant. J. Plant Growth Regul. 2019, 39, 1221–1235. [Google Scholar] [CrossRef]
- Zheng, X.; Tan, X.D.; Allan, C.A.; Zuo, B.; Zhao, Y.; Reiter, J.R.; Wang, L.; Wang, Z.; Guo, Y.; Zhou, J. Chloroplastic biosynthesis of melatonin and its involvement in protection of plants from salt stress. Sci. Rep. 2017, 7, 41236. [Google Scholar] [CrossRef]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, Z.; Zhang, H.; Wang, X.; Huang, R. Transcriptional modulation of ethylene response factor protein JERF3 in the oxidative stress response enhances tolerance of tobacco seedlings to salt, drought, and freezing. Plant Physiol. 2008, 148, 1953–1963. [Google Scholar] [CrossRef] [Green Version]
- Yang, A.; Dai, X.; Zhang, W. A R2R3-type MYB gene, OsMYB2, is involved in salt, cold, and dehydration tolerance in rice. J. Exp. Bot. 2012, 63, 2541–2556. [Google Scholar] [CrossRef]
- Sholpan, D.; Karen, S.; Jesse, C.; Ron, M. Zinc-Finger Protein Zat12 Plays a Central Role in Reactive Oxygen and Abiotic Stress Signaling in Arabidopsis. Plant Physiol. 2005, 139, 847–856. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, S.; Fang, Q.; Wang, Y.; Zhong, S.; Dong, B.; Zhao, H. OfSPL11 Gene from Osmanthus fragrans Promotes Plant Growth and Oxidative Damage Reduction to Enhance Salt Tolerance in Arabidopsis. Horticulturae 2022, 8, 412. https://doi.org/10.3390/horticulturae8050412
Zhu S, Fang Q, Wang Y, Zhong S, Dong B, Zhao H. OfSPL11 Gene from Osmanthus fragrans Promotes Plant Growth and Oxidative Damage Reduction to Enhance Salt Tolerance in Arabidopsis. Horticulturae. 2022; 8(5):412. https://doi.org/10.3390/horticulturae8050412
Chicago/Turabian StyleZhu, Shoukuo, Qiu Fang, Yiguang Wang, Shiwei Zhong, Bin Dong, and Hongbo Zhao. 2022. "OfSPL11 Gene from Osmanthus fragrans Promotes Plant Growth and Oxidative Damage Reduction to Enhance Salt Tolerance in Arabidopsis" Horticulturae 8, no. 5: 412. https://doi.org/10.3390/horticulturae8050412
APA StyleZhu, S., Fang, Q., Wang, Y., Zhong, S., Dong, B., & Zhao, H. (2022). OfSPL11 Gene from Osmanthus fragrans Promotes Plant Growth and Oxidative Damage Reduction to Enhance Salt Tolerance in Arabidopsis. Horticulturae, 8(5), 412. https://doi.org/10.3390/horticulturae8050412