
In Vitro Studies on Seed Germination of the Mediterranean Species Anthyllis barba-jovis to Facilitate Its Introduction into the Floriculture Industry


Abstract
1. Introduction
2. Materials and Methods
2.1. Seed Material
2.2. Seed Viability
2.3. Mechanical Scarification
2.4. In Vitro Germination
2.5. Experimental Design and Statistical Analysis
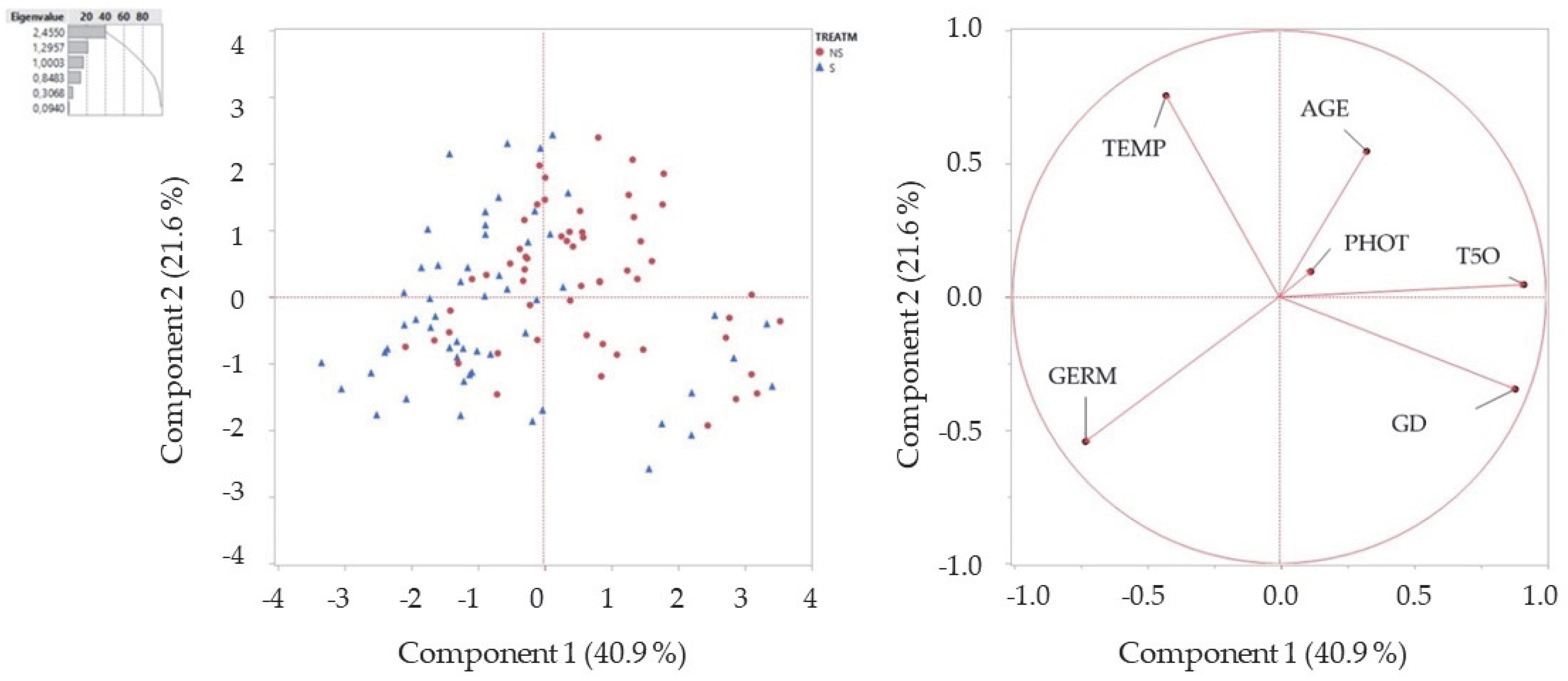
3. Results
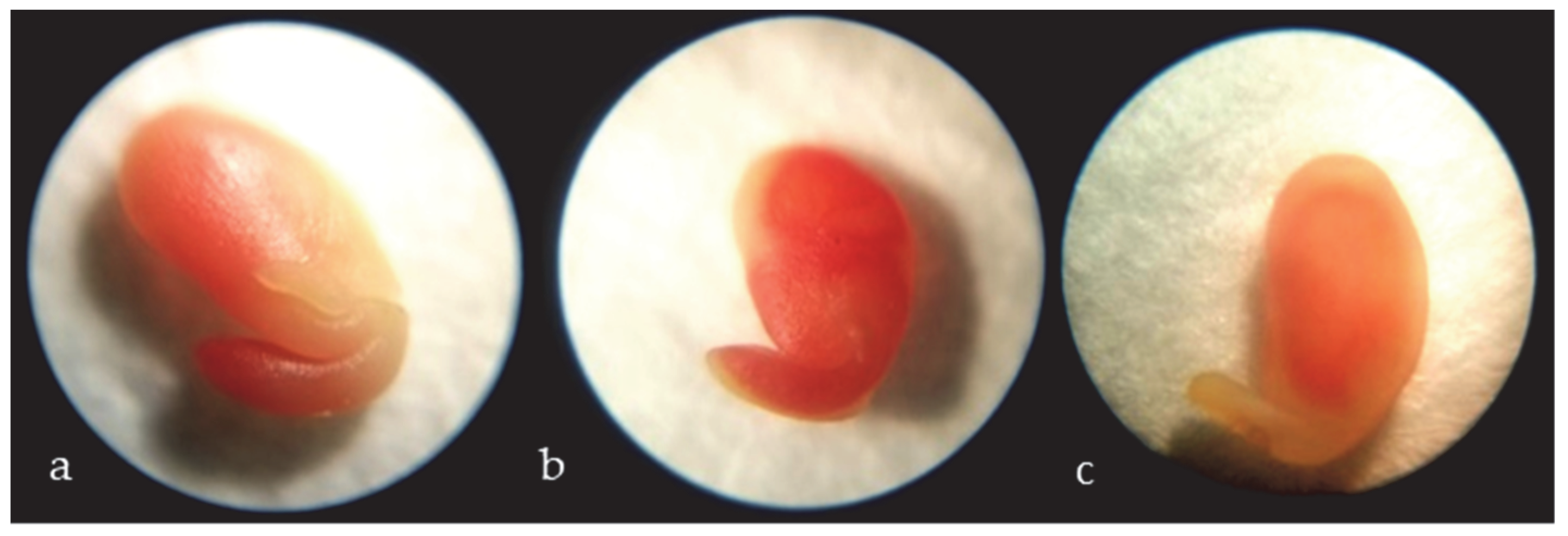
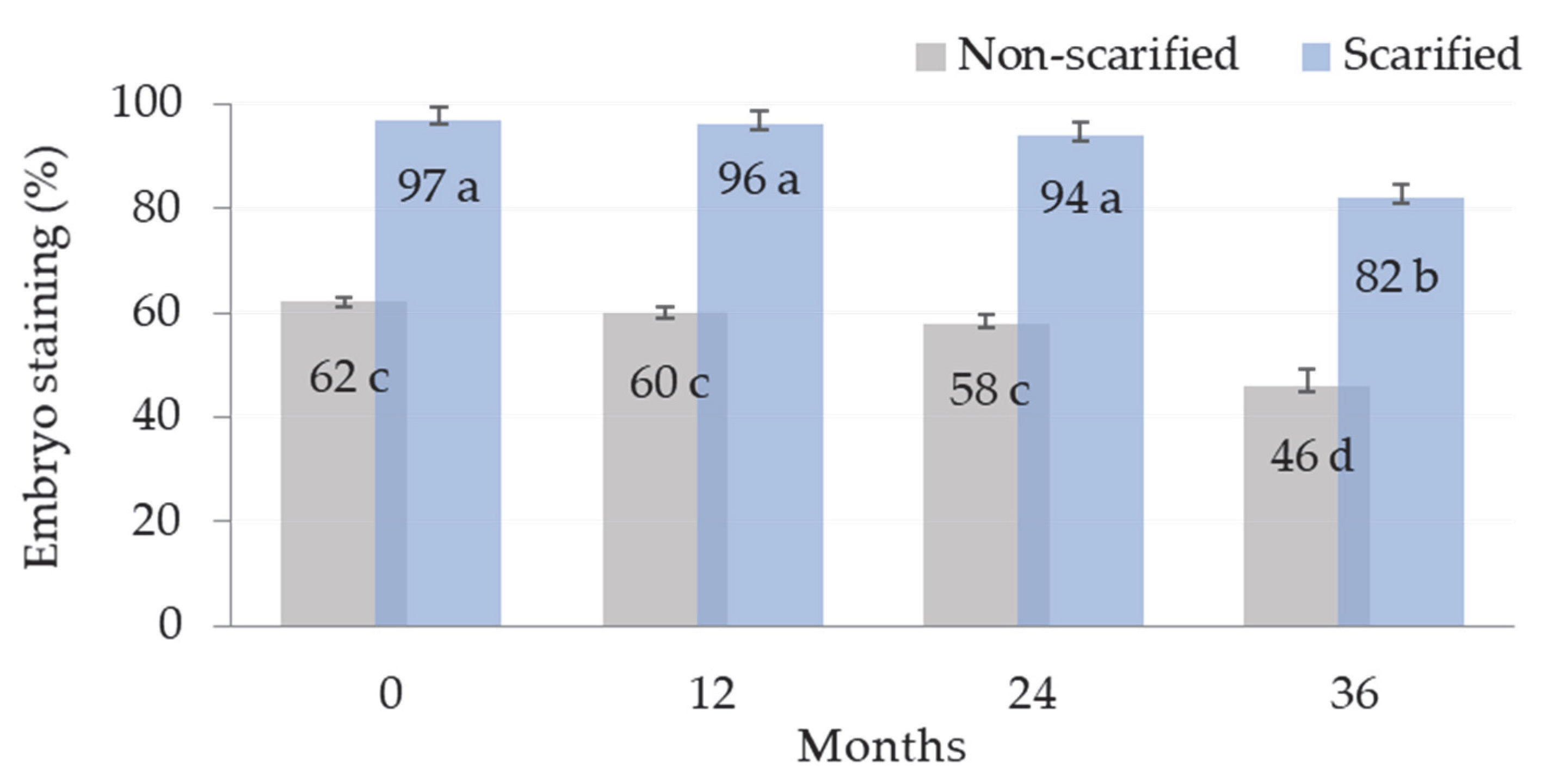
3.1. Seed Viability
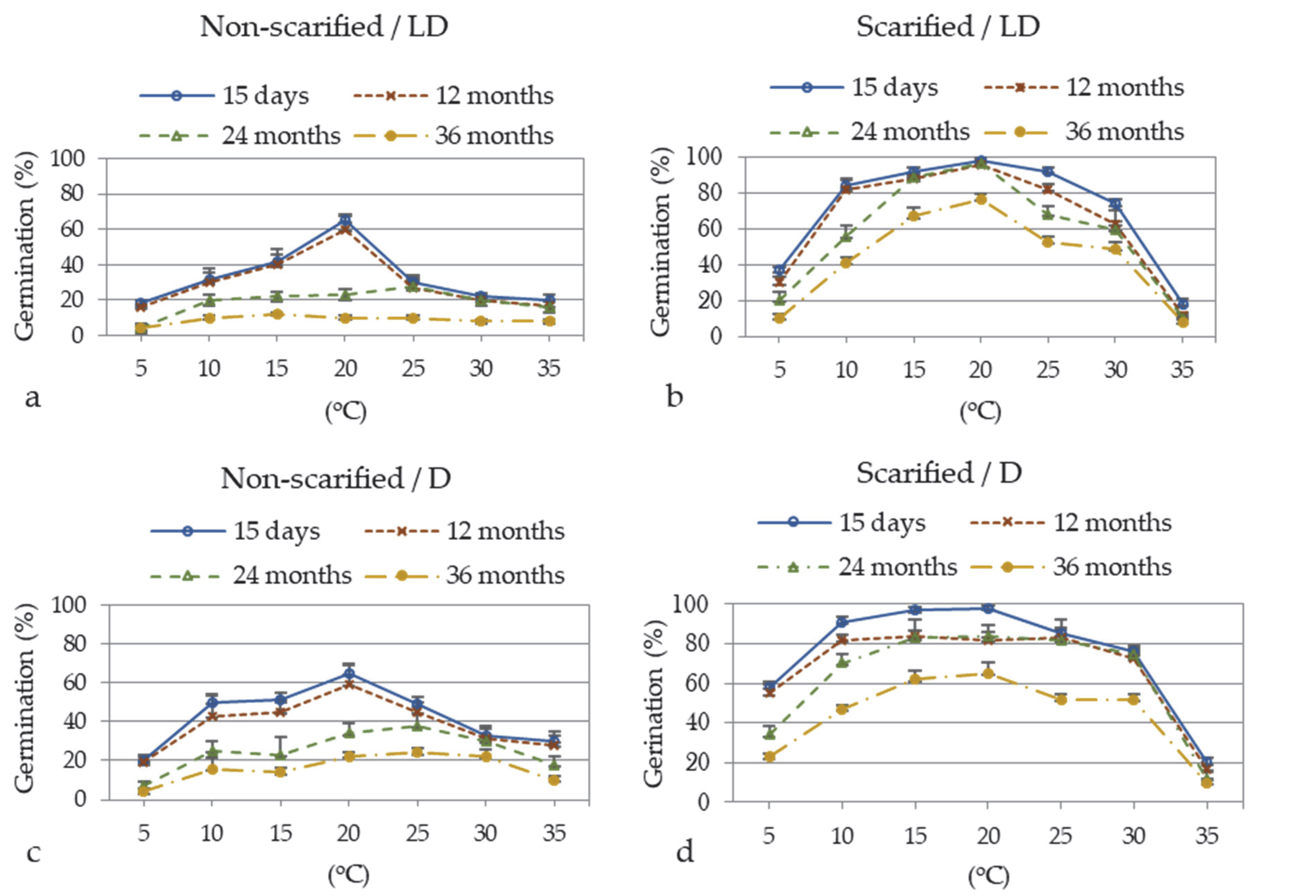
3.2. Seed Germination
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Arianoutsou, M. Landscape changes in Mediterranean Ecosystems of Greece: Implications for Fire and Biodiversity issues. J. Mediterr. Ecol. 2001, 2, 165–178. [Google Scholar]
- Biondi, E.; Vagge, I.; Mossa, L. On the phytosociological importance of Anthyllis barba-jovis L. Coll. Phytosoc. 1997, 27, 95–104. [Google Scholar]
- Paradis, G. Observations sur l’espèce rare et protégée Anthyllis barba-jovis L. (Fabaceae) en Corse: Description de ses stations et phytosociologie. J. Bot. Soc. Bot. 1997, 4, 33–44. [Google Scholar]
- Pignatti, S. Flora d’Italia; Edagricole: Bologna, Italy, 1992; Volume 1. [Google Scholar]
- Motta, F. Nel Mondo Della Natura; Enciclopedia di Scienze Naturali: Milano, Italy, 1992; Volume 1. [Google Scholar]
- Tutin, G.T.; Heywood, V.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb., D.A. Flora Europaea; Cambridge University Press: Cambridge, UK, 1972; Volume 4. [Google Scholar]
- Trinajstic, I. Anthyllis barba-jovis L. Crvena Knjiga Biljnih Vrsta Republike Hrvatske, Ministarstvo Graditeljstva i Zastite Okolisa; Zavod za Zastitu Prirode: Zagreb, Croatia, 1994; pp. 21–23. [Google Scholar]
- Danton, P.; Baffray, M. Inventaire des Plantes Protégées en France; Nathan & A.F.C.E.V.: Paris, France; Mulhouse, France, 1995; p. 294. [Google Scholar]
- Ballesteros, D.; Meloni, F.; Bacchetta, G. (Eds) Manual for the Propagation of Selected Mediterranean Native Plant Species. Ecoplantmed ENPI CBC-MED 2015, 127. Available online: http://www.ecoplantmed.eu/en/publications/propagation_manual (accessed on 28 September 2022).
- Conti., F.; Manzi, A.; Pedrotti, F. Liste Rosse Regionali delle Piante d’Italia; WWF-S.B.I.; Università di Camerino: Camerino, Italy, 1997. [Google Scholar]
- Papafotiou, M.; Bertsouklis, K.F.; Martini, A.N.; Vlachou, G.; Akoumianaki-Ioannidou, A.; Kanellou, E.; Kartsonas, E.D. Evaluation of the establishment of native Mediterranean plant species suggested for landscape enhancement in archaeological sites of Greece. Acta Hortic. 2017, 1189, 177–180. [Google Scholar] [CrossRef]
- Morbidoni, M.; Estrelles, E.; Soriano, P.; Martínez-Solís, I.; Biondi, E. Effects of environmental factors on seed germination of Anthyllis barba-jovis L. Plant Biosyst. 2008, 142, 275–286. [Google Scholar] [CrossRef]
- Benelli, G.; Benvenuti, S.; Scaramozzino, P.L.; Canale, A. Food for honeybees? Pollinators and seed set of Anthyllis barba-jovis L. (Fabaceae) in arid coastal areas of the Mediterranean basin. Saudi J. Biol. Sci. 2017, 24, 1056–1060. [Google Scholar] [CrossRef]
- Pistelli, L.; Noccioli, C.; Bertoli, A.; Scapecchi, G.; Potenza, D. Chemical composition and volatile constituents of Anthyllis barba-jovis. Nat. Prod. Res. 2007, 21, 418–425. [Google Scholar] [CrossRef]
- Bewley, J.D.; Black, M. Some Ecophysiological Aspects of Germination. In Seeds-Physiology of Development and Germination, 2nd ed.; Bewley, J.D., Black, M., Eds.; Plenum Press: New York, NY, USA, 1994; pp. 273–292. [Google Scholar]
- Tomer, R.P.; Kumari, P. Hard seed studies in black gram (Vigna mungo L.). Seed Sci. Technol. 1991, 19, 51–56. [Google Scholar]
- Baskin, J.M.; Baskin, C.C.; Li, X. Taxonomy, anatomy and evolution of physical dormancy in seeds. Plant Spec. Biol. 2000, 15, 139–152. [Google Scholar] [CrossRef]
- Van Staden, J.; Manning, J.C.; Kelly, K.M. Legume seeds-The structure: Function equation. In Advances in Legume Research; Stirton, C.H.L., Zarucchi, J.L., Eds.; Monographs in Systematic Botany from the Missouri Botanical Garden no. 29.; Missouri Botanical Garden: St. Louis, MO, USA, 1989; pp. 417–450. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Academic Press: San Diego, CA, USA, 1998; p. 37. [Google Scholar]
- Wang, Y.R.; Hanson, J.; Mariam, Y.W. Effect of sulfuric acid pretreatment on breaking hard seed dormancy in diverse accessions of five wild Vigna species. Seed Sci. Technol. 2007, 35, 550–559. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Breaking Seed Dormancy during Dry Storage: A Useful Tool or Major Problem for Successful Restoration via Direct Seeding? Plants 2020, 9, 636. [Google Scholar] [CrossRef] [PubMed]
- Gutterman, Y. Strategies of seed dispersal and germination in plants inhabiting deserts. Bot. Rev. 1994, 60, 373–425. [Google Scholar] [CrossRef]
- Ellner, S. ESS germination fractions in randomly varying environments. I. Logistic-type models. Theor. Popul. Biol. 1985, 28, 50–79. [Google Scholar] [CrossRef]
- Moore, R.P.E. Handbook on Tetrazolium Testing, 1st ed.; The International Seed Testing Association (ISTA): Bassersdorf, Switzerland, 1985; p. 99. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- International Seed Testing Association. International rules for seed testing. Seed Sci. Technol. 1999, 27, 333. [Google Scholar]
- Thanos, C.A.; Kadis, C.C.; Skarou, F. Ecophysiology of germination in the aromatic plants thyme, savory and oregano (Labiatae). Seed Sci. Res. 1995, 5, 161–170. [Google Scholar] [CrossRef]
- Kadis, C.; Georghiou, K. Seed dispersal and germination behavior of three threatened endemic labiates of Cyprus. Plant Spec. Biol. 2010, 25, 77–84. [Google Scholar] [CrossRef]
- Bertsouklis, K.; Papafotiou, M. Seed germination of Arbutus unedo, A. andrachne and their natural hybrid A. andrachnoides in relation to temperature and period of storage. HortScience 2013, 48, 347–351. [Google Scholar] [CrossRef]
- Ibanez, A.N.; Passera, C.B. Factors affecting the germination of albaida (Anthyllis cytisoides L.), a forage legume of the Mediterranean coast. J. Arid Environ. 1997, 35, 225–231. [Google Scholar] [CrossRef]
- Papafotiou, M.; Stragas, J. Seed germination and in vitro propagation of Dianthus fruticosus. L. Acta Hortic. 2009, 813, 481–484. [Google Scholar] [CrossRef]
- Bertsouklis, K.F.; Papafotiou, M. Studies on propagation of Globularia alypum L. Acta Hortic. 2010, 885, 73–77. [Google Scholar] [CrossRef]
- Papafotiou, M.; Kalantzis, A. Seed germination and in vitro propagation of Sideritis athoa. Acta Hortic. 2009, 813, 471–476. [Google Scholar] [CrossRef]
- Thanos, C.A.; Doussi, M.A. Ecophysiology of seed germination in endemic Labiates of Crete. Israel J. Plant Sci. 1995, 43, 227–237. [Google Scholar] [CrossRef]
- Papafotiou, M.; Martini, A.N. In Vitro Seed and Clonal Propagation of the Mediterranean Aromatic and Medicinal Plant Teucrium capitatum. HortScience 2016, 51, 403–411. [Google Scholar] [CrossRef]
- Vlachou, G.; Papafotiou, Μ.; Bertsouklis, K. Studies on seed germination and micropropagation of Clinopodium nepeta a medicinal and aromatic plant. HortScience 2019, 54, 1558–1564. [Google Scholar] [CrossRef]
- Vlachou, G.; Papafotiou, Μ.; Bertsouklis, K. Seed germination, micropropagation from adult and juvenile origin explants and address of hyperhydricity of the Cretan endemic herb Calamintha cretica. Not. Bot. Horti Agrobot. Cluj Napoca 2020, 48, 1504–1518. [Google Scholar] [CrossRef]
- Travlos, I.S.; Economou, G.; Karamanos, A.I. Germination and emergence of the hard seed coated Tylosema esculentum (Burch) A. Schreib in response to different pre-sowing seed treatments. J. Arid Environ. 2007, 68, 501–507. [Google Scholar] [CrossRef]
- Talei, D.; Valdiani, A.; Abdullah, M.; Hassan, S.A. A rapid and effective method for dormancy breakage and germination of King of Bitters (Andrographis paniculata Nees.) seeds. Maydica 2012, 57, 98–105. [Google Scholar]
- Rodrigues-Junior, A.G.; Baskin, C.C.; Baskin, J.M.; Garcia, Q.S. Sensitivity cycling in physically dormant seeds of the Neotropical tree Senna multijuga (Fabaceae). Plant Biol. J. 2008, 20, 698–706. [Google Scholar] [CrossRef]
- Soltani, E.; Baskin, J.; Baskin, C.; Benakashani, F. A meta-analysis of the effects of treatments used to break dormancy in seeds of the megagenus Astragalus (Fabaceae). Seed Sci. Res. 2020, 30, 224–233. [Google Scholar] [CrossRef]
- Khuat, Q.V.; Kalashnikova, E.A.; Kirakosyan, R.N.; Nguyen, H.T.; Baranova, E.N.; Khaliluev, M.R. Improvement of In Vitro Seed Germination and Micropropagation of Amomum tsao-ko (Zingiberaceae Lindl.). Horticulturae 2022, 8, 640. [Google Scholar] [CrossRef]
- Bonner, F.T.; Franklin, T.; Karrfalt, R.P.; Robert, P. (Eds.) The Woody Plant Seed Manual. In Agricultural Handbook; No 727; U.S. Department of Agriculture, Food Service: Washington, DC, USA, 2008; p. 1223. [Google Scholar]
- Missanjo, E.; Maya, C.; Kapira, D.; Banda, H.; Kamanga-Thole, G. Effect of seed size and pretreatment methods on germination of Albizia lebbeck. Int. Sch Res. Not. 2013, 2013, 969026. [Google Scholar] [CrossRef]
- Bouteiller, X.P.; Porté, A.J.; Mariette, S.; Monty, A. Using automated sanding to homogeneously break seed dormancy in black locust (Robinia pseudoacacia L., Fabaceae). Seed Sci. Res. 2017, 27, 243–250. [Google Scholar] [CrossRef]
- Guàrdia, R.; Gallart, F.; Ninot, J.M. Soil seed bank and seedling dynamics in badlands of the Upper Llobregat basin (Pyrenees). Catena 2000, 40, 189–202. [Google Scholar] [CrossRef]
- Acherkouk, M.; Aberkani, K.; Sabre, I.A.; Maatougui, A.; Amhamdi, H.; Halou, B. Effect of seeds pretreatment and storage on improvement of the germination and emergence of Anthyllis cytisoides L. Afr. J. Agric. Res. 2017, 12, 2642–2650. [Google Scholar] [CrossRef]
- Giusti, L.; Grau, A. Inhibidores de la germinación en Atriplex cordobensis Gand. et Stucker (Chenopodiaceae). Lilloa 1983, 36, 143–149. [Google Scholar]
- Adams, C.R.; Early, M.P. Principles of Horticulture, 4th ed.; Elsevier Butterworth-Heinemann Publication: Burlington, MA, USA, 2004; p. 70. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Principal Components | ||
| 1 | 2 | 3 |
| % Contribution of variability | ||
| 40.91 | 21.59 | 16.67 |
| Related variables | ||
| T50 | AGE | PHOT |
| GT | TEMP | |
| GERM | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertsouklis, K.; Vlachou, G.; Trigka, M.; Papafotiou, M. In Vitro Studies on Seed Germination of the Mediterranean Species Anthyllis barba-jovis to Facilitate Its Introduction into the Floriculture Industry. Horticulturae 2022, 8, 889. https://doi.org/10.3390/horticulturae8100889
Bertsouklis K, Vlachou G, Trigka M, Papafotiou M. In Vitro Studies on Seed Germination of the Mediterranean Species Anthyllis barba-jovis to Facilitate Its Introduction into the Floriculture Industry. Horticulturae. 2022; 8(10):889. https://doi.org/10.3390/horticulturae8100889
Chicago/Turabian StyleBertsouklis, Konstantinos, Georgia Vlachou, Marina Trigka, and Maria Papafotiou. 2022. "In Vitro Studies on Seed Germination of the Mediterranean Species Anthyllis barba-jovis to Facilitate Its Introduction into the Floriculture Industry" Horticulturae 8, no. 10: 889. https://doi.org/10.3390/horticulturae8100889
APA StyleBertsouklis, K., Vlachou, G., Trigka, M., & Papafotiou, M. (2022). In Vitro Studies on Seed Germination of the Mediterranean Species Anthyllis barba-jovis to Facilitate Its Introduction into the Floriculture Industry. Horticulturae, 8(10), 889. https://doi.org/10.3390/horticulturae8100889