
Analysis of Compounds with Oenological Interest in Somatic Variants of Grapevines

 ,
,  ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Chemical Compounds
2.3. Genetic Analysis
2.4. Sample Processing
2.5. Ultrasound-Assisted Extraction
2.6. Analytical Measurements
2.6.1. Sugars
2.6.2. Organic Acids
2.6.3. Phenolic Compounds (Excluding Anthocyanins)
2.6.4. Identification of Anthocyanins
2.6.5. Analysis of Anthocyanins
2.7. Data Analysis
3. Results and Discussion
3.1. Genetic Analysis
3.2. Sugar Evolution during Ripening Period
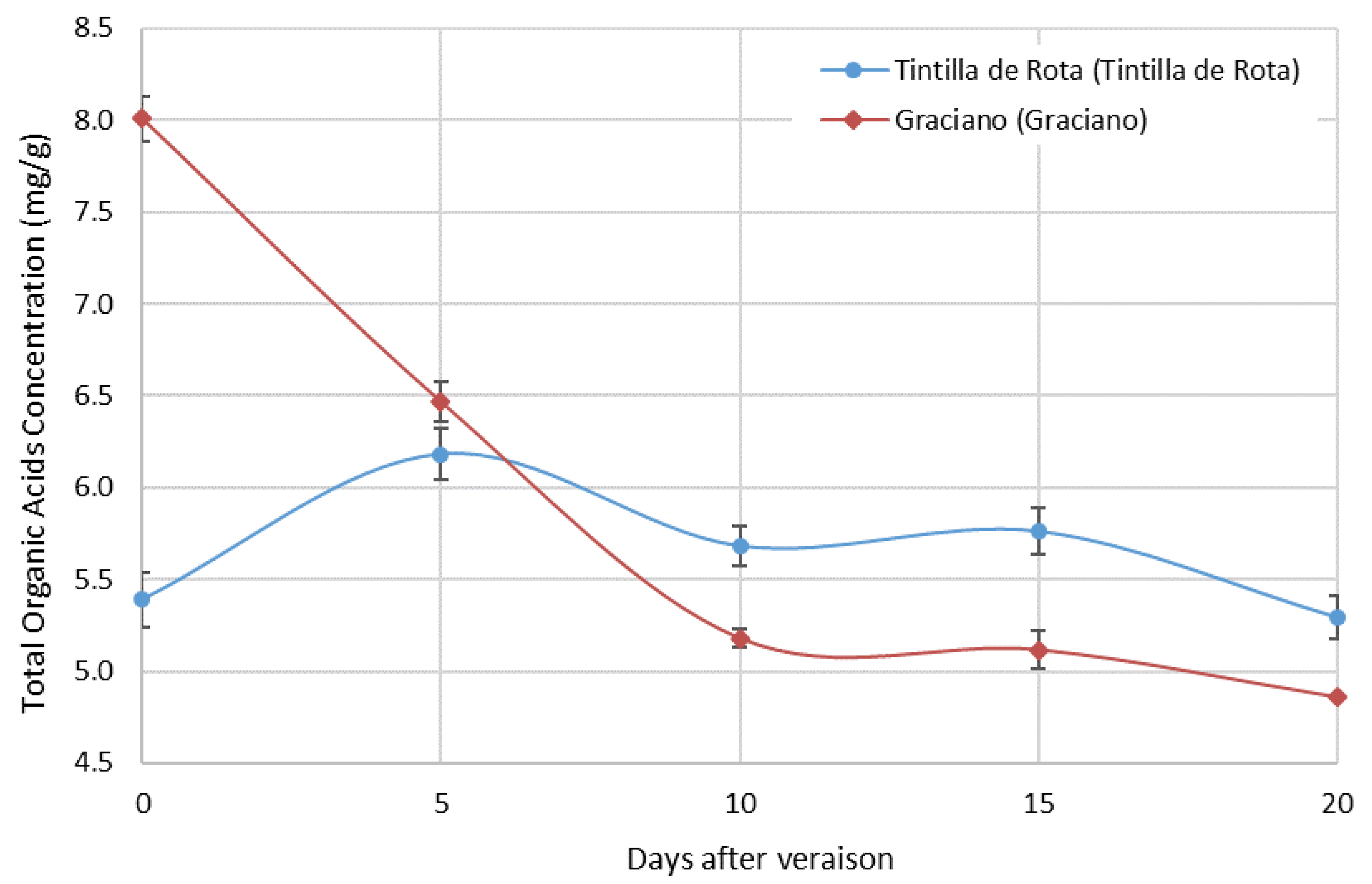
3.3. Organic Acid Content during Grape Ripening
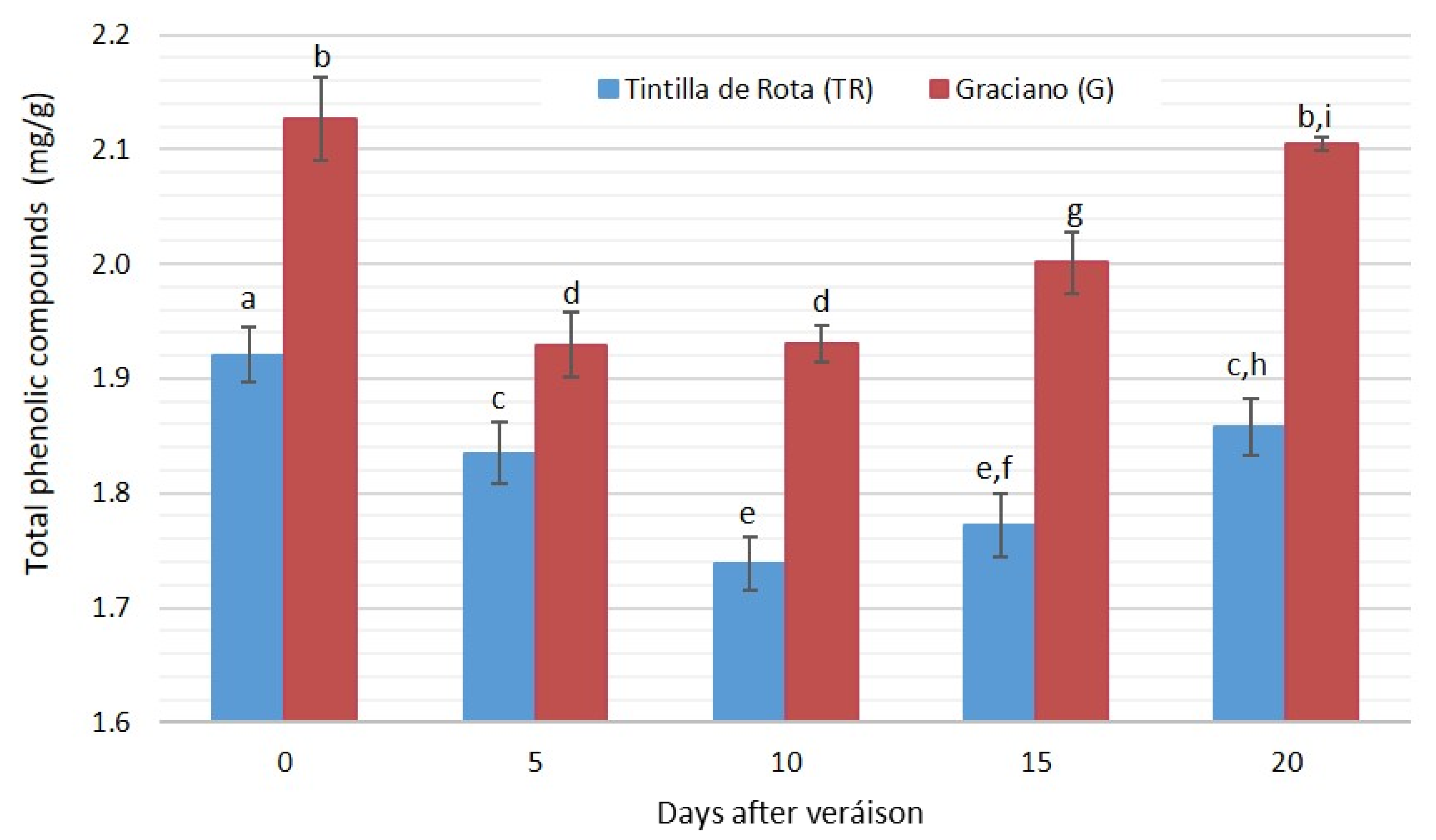
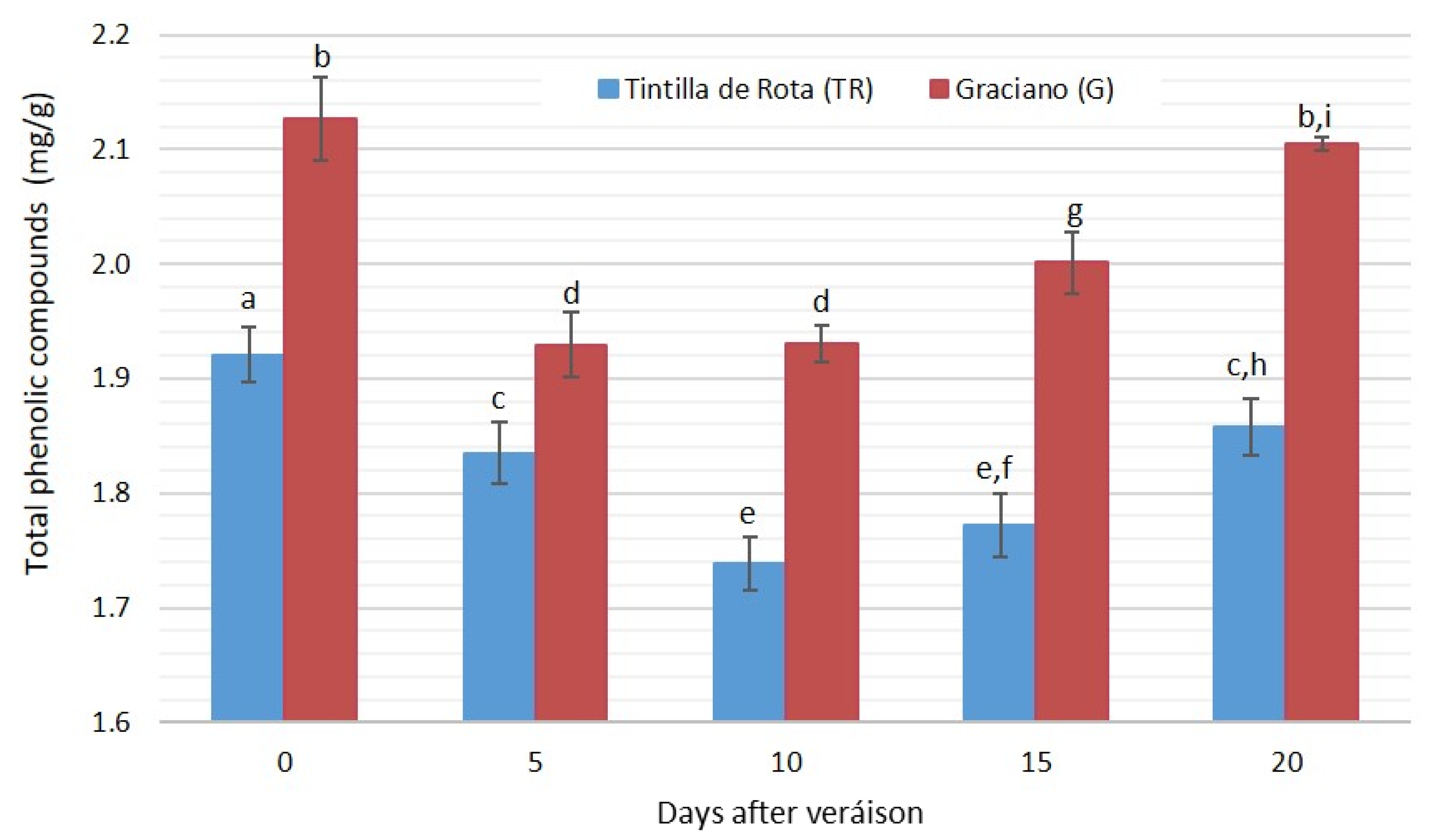
3.4. Phenolic Compound Evolution during Grape Ripening (Excluding Anthocyanins)
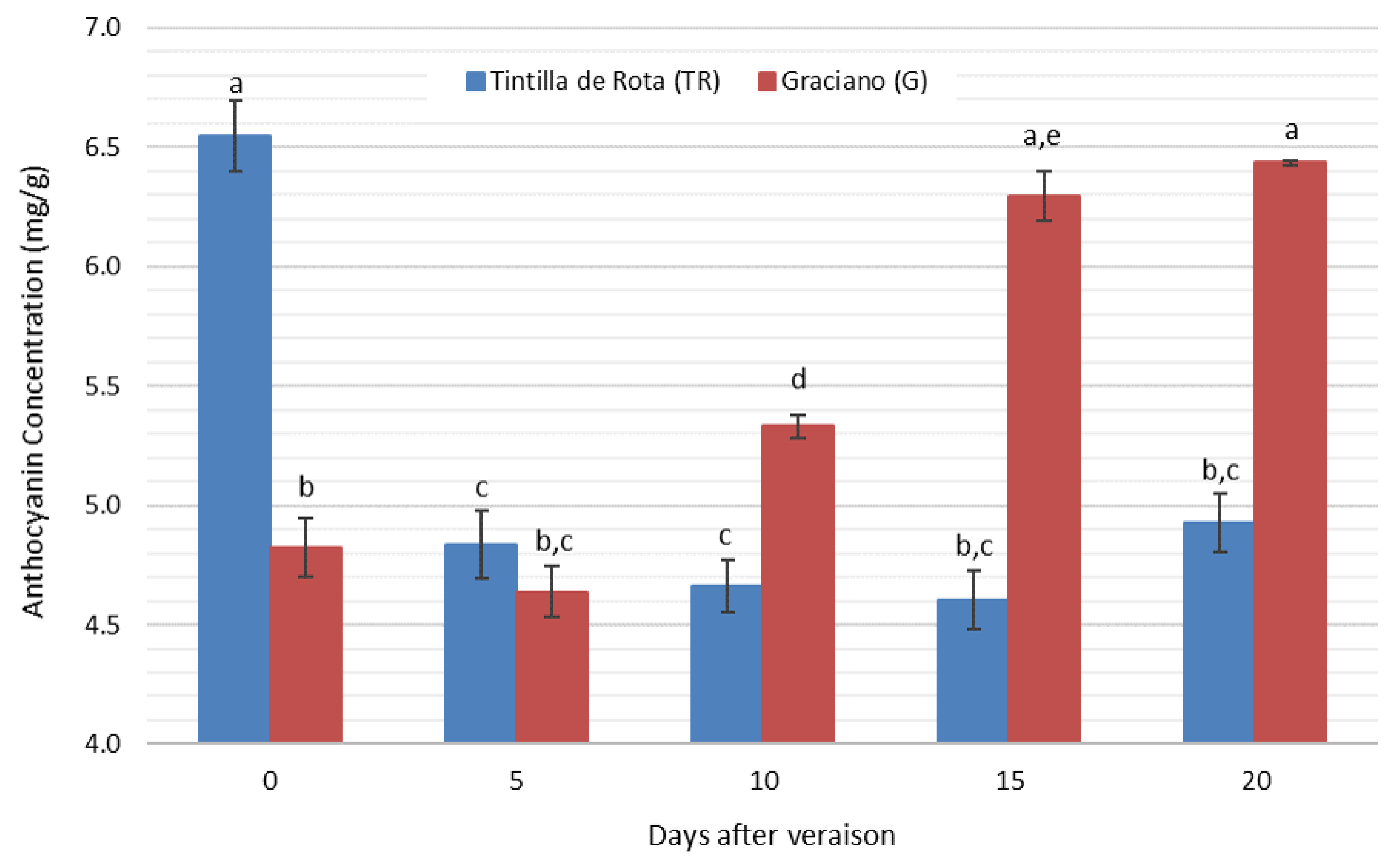
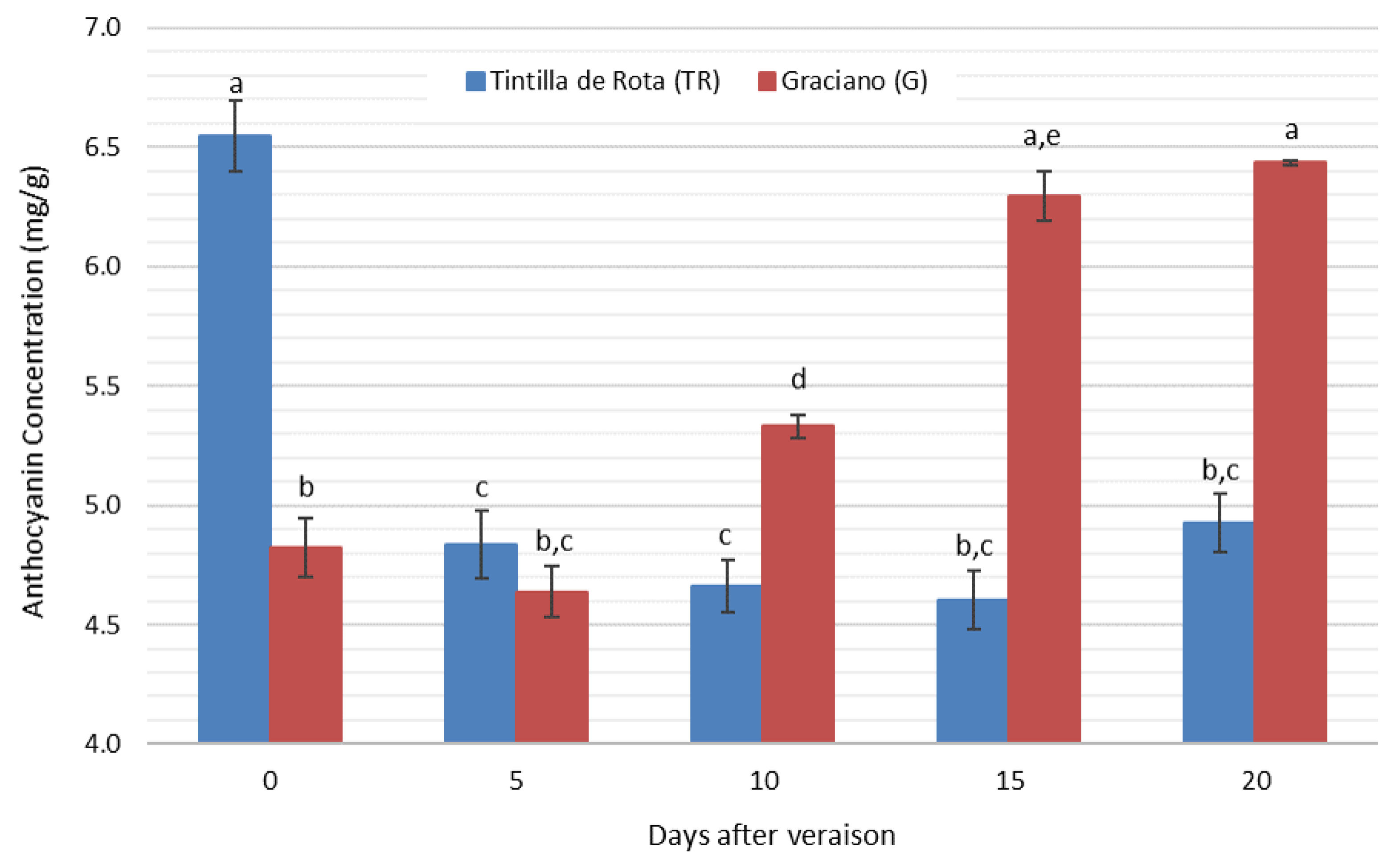
3.5. Anthocyanin Concentration Evolution during Grape Ripening
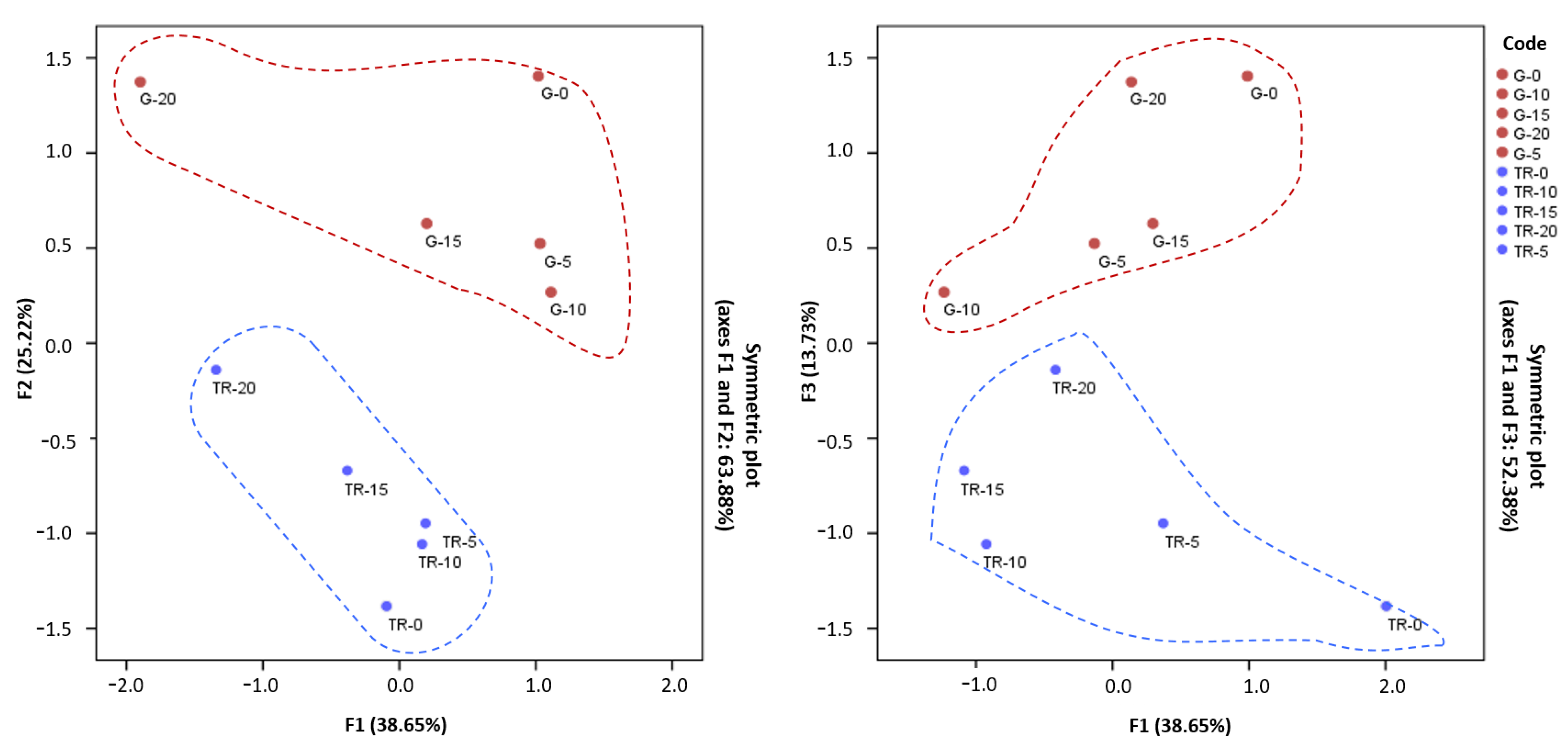
3.6. Principal Component Analysis (PCA)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Sancho-Galán, P.; Amores-Arrocha, A.; Palacios, V.; Jiménez-Cantizano, A. Effect of Grape Over-Ripening and Its Skin Presence on White Wine Alcoholic Fermentation in a Warm Climate Zone. Foods 2021, 10, 1583. [Google Scholar] [CrossRef] [PubMed]
- Riaz, S.; De Lorenzis, G.; Velasco, D.; Koehmsted, A.; Maghradze, D.; Bobokashvili, Z.; Musayev, M.; Zdunic, G.; Laucou, V.; Walker, M.A.; et al. Genetic diversity analysis of cultivated and wild grapevine (Vitis vinifera L.) accessions around the Mediterranean basin and Central Asia. BMC Plant Biol. 2018, 18, 137. [Google Scholar] [CrossRef]
- Jiménez-Cantizano, A.; García de Lujan, A.; Arroyo-García, R. Molecular characterization of table grape varieties preserved in the Rancho de la Merced Grapevine Germplasm Bank (Spain). Vitis 2018, 57, 93–101. [Google Scholar]
- Distribution of the World’s Grapevine Varieties. Available online: https://www.oiv.int/public/medias/5888/en-distribution-of-the-worlds-grapevine-varieties.pdf (accessed on 11 November 2021).
- Moncada, X.; Pelsy, F.; Merdinoglu, D.; Hinrichsen, P. Genetic diversity and geographical dispersal in grapevine clones revealed by microsatellite markers. Genome 2006, 49, 1459–1472. [Google Scholar] [CrossRef]
- Carbonell-Bejerano, P.; Royo, C.; Mauri, N.; Ibáñez, J.; Martínez-Zapater, J.M. Somatic variation and cultivar innovation in grapevine. In Advances in Grape and Wine Biotechnology, 1st ed.; Morata, A., Loira, I., Eds.; Intechopen: London, UK, 2016; p. 8. [Google Scholar]
- This, P.; Martínez-Zapater, J.M.; Péros, J.P.; Lacombe, T. Natural Variation in Vitis. In Genetics, Genomics and Breeding of Grapes, 1st ed.; Adam-Blondon, A.F., Martínez-Zapater, J.M., Kole, C., Eds.; Science Publishers: New York, NY, USA, 2011; p. 30. [Google Scholar]
- Bourisquot, J.M.; This, P. Essai de définition du cépage. Prog. Agric. Vitic. 1999, 116, 359–361. [Google Scholar]
- Graça, A.; Manso, J.; Sandeman, G. A systematic approach to organize grapevine varieties’ naming at the international level while retaining their cultural heritage and allegiance across different countries and wine regions. In Proceedings of the 37th OIV World Congress of Vine and Wine, Mendoza, Argentina, 9–14 November 2014. [Google Scholar]
- Niculcea, M.; Martínez-Lapuente, L.; Guadalupe, Z.; Sanchez-Díaz, M.; Ayestarán, B.; Antolín, C. Characterization of phenolic composition of Vitis vinifera L. ‘Tempranillo’and ‘Graciano’ subjected to deficit irrigation during berry development. Vitis 2015, 54, 9–16. [Google Scholar]
- Dominguez, N.; García-Escudero, E.; Romero, I.; Benito, A.; Martín, I. Leaf blade and petiole nutritional evolution and variability throughout the crop season for Vitis vinifera L. cv. Graciano. Span. J. Agric. Res. 2015, 13, e0801. [Google Scholar] [CrossRef] [Green Version]
- Roxas Clemente, S. Ensayo Sobre las Variedades de vid que Vegetan en Andalucía, 1st ed.; Imprenta de Villapando: Madrid, Spain, 1807; pp. 111–113. [Google Scholar]
- Vitis International Variety Catalogue. Available online: www.vivc.de (accessed on 11 November 2021).
- Jiménez-Cantizano, A. Caracterización Molecular del Banco de Germoplasma de vid del Rancho de la Merced. Ph.D. Thesis, Universidad de Cadiz, Cadiz, Spain, 2014. [Google Scholar]
- Puertas, B.; Lara, M.; Serrano, M.J.; Varcárcel, M.; Cruz, S.; Jiménez Cantizano, A.; Garcia de Luján, A. Caracterización vitícola y enológica de las variedades tintas Graciano, Monastrell, Tempranillo y Tintilla de Rota, cultiva-das en zona cálida. In Proceedings of the Congreso Nacional de Enologos, Santiago de Compostela, Spain, 26–28 April 2002. [Google Scholar]
- Organisation Internationale de la Vigne et du Vin (OIV). OIV Descriptor List for Grape Varieties and Vitis Species, 2nd ed.; OIV: Paris, France, 2009. [Google Scholar]
- This, P.; Jung, A.; Boccacci, P.; Borrego, J.; Botta, R.; Costantini, L.; Crespan, M.; Dangl, G.S.; Eisenheld, C.; Ferreira-Monteiro, F.; et al. Development of a standard set of microsatellite reference alleles for identification of grape cultivars. Theor. Appl. Genet. 2004, 109, 1448–1458. [Google Scholar] [CrossRef] [PubMed]
- Zinelabidine, L.H.; Cunha, J.; Eiras-Dias, J.E.; Cabello, F.; Martínez-Zapater, J.M.; Ibáñez-Marcos, J. Pedigree analysis of the Spanish grapevine cultivar ‘Hebén’. Vitis 2015, 54, 81–86. [Google Scholar]
- Moreno-Vigara, J.J.; Peinado-Amores, R.A. Química Enológica, 1st ed.; Ediciones Mundi-Prensa; AMV Ediciones: Madrid, Spain, 2010. [Google Scholar]
- Ferreira, V.; Fernandes, F.; Pinto-Carnide, O.; Valentão, P.; Falco, V.; Martín, J.P.; Ortiz, J.M.; Arroyo-García, R.; Andrade, P.B.; Castro, I. Identification of Vitis vinifera L. grape berry skin color mutants and polyphenolic profile. Food Chem. 2016, 194, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Jiménez Cantizano, A.; Amores Arrocha, A.; Gutiérrez Escobar, R.; Palacios, V. Identification and relationship of the autochthonous ‘Romé’ and ‘Rome Tinto’ grapevine cultivars. Span. J. Agric. Res. 2018, 16, e07SC02. [Google Scholar] [CrossRef]
- Santesteban, L.G.; Miranda, C.; Royo, J.B. Vegetative growth, reproductive development and vineyard balance. In Methodologies and Results in Grapevine Research, 1st ed.; Delrot, S., Medrano, H., Or, E., Bavaresco, L., Grando, S., Eds.; Springer: New York, NY, USA, 2010; pp. 45–56. [Google Scholar]
- Sancho-Galán, P.; Amores-Arrocha, A.; Jiménez-Cantazino, A.; Ferreiro-González, M.; Palacios, V.; Barbero, G.F. Ultrasound-assisted extraction of anthocyanins and total phenolic compounds in Vitis vinifera L. Tempranillo winemaking lees. Vitis 2019, 58, 39–47. [Google Scholar]
- Amores-Arrocha, A. Aplicación del Polen de abeja como Activador en el Proceso de Fermentación Alcohólica. Ph.D. Thesis, Facultad de Ciencias, Universidad de Cádiz, Cádiz, Spain, July 2018. [Google Scholar]
- Sancho-Galán, P.; Amores-Arrocha, A.; Jiménez-Cantizano, A.; Palacios, V. Use of Multiflora Bee Pollen as a Flor Velum Yeast Growth Activator in Biological Aging Wines. Molecules 2019, 24, 1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Peredo, A.V.G.; Vázquez-Espinosa, M.; Piñeiro, Z.; Espada-Bellido, E.; Ferreiro-González, M.; Barbero, G.F.; Palma, M. Development of a rapid and accurate UHPLC-PDA-FL, method for the quantification of phenolic compounds in grapes. Food Chem. 2021, 334, 127569. [Google Scholar] [CrossRef]
- Pereira, D.T.V.; Tarone, A.G.; Cazarin, C.B.B.; Barbero, G.F.; Martínez, J. Pressurized liquid extraction of bioactive compound from grape marc. J. Food Eng. 2019, 240, 105–113. [Google Scholar] [CrossRef]
- Ribéreau-Gayon, P.; Dubourdieu, D.; Donèche, B.; Lonvaud, A. Handbook of Enology, 2nd ed.; John Wiley & Sons Ltd.: Chichester, UK, 2006. [Google Scholar]
- Kliewer, W.M. The Glucose-Fructose Ratio of Vitis Vinifera Grapes. Am. J. Enol. Vitic. 1967, 18, 33–41. [Google Scholar]
- Kuhn, N.; Guan, L.; Dai, Z.W.; Wu, B.; Lauvergeat, V.; Gomès, E.; Li, S.H.; Godoy, F.; Arce-Johnson, P.; Delrot, S. Berry ripening: Recently heard through the grapevine. J. Exp. Bot. 2014, 65, 4543–4559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Z.; Sang, M.; Fan, P.; Wu, B.; Wang, L.; Duan, W.; Li, S. Changes of Polyphenols, Sugars, and Organic Acid in 5 Vitis Genotypes during Berry Ripening. J. Food Sci. 2011, 76, 1231–1238. [Google Scholar] [CrossRef]
- Chivite, J.; Raventós, M.; Castro, E. Gestión de pH en El Vino de Calidad, 1st ed.; Fundación Para la Cultura Del Vino: Madrid, Spain, 2005; pp. 9–15. [Google Scholar]
- Gil, P.L.O. Efectos del Deshojado Precoz, Durante Cuatro Años, Sobre las Características de la Producción en las Variedades Tempranillo, Mazuelo y Graciano (Vitis vinífera L.). Ph.D. Thesis, Universidad de La Rioja, Logroño, Spain, 2010. [Google Scholar]
- Tardaguilloa, J.; de Toda, F.M.; Poni, S.; Diago, M.P. Impact of early leaf removal on yield and fruit and wine composition of Vitis vinifera L. Graciano and Carignan. Am. J. Enol. Vitic. 2010, 61, 372–381. [Google Scholar]
- Gutiérrez-Gamboa, G.; Portu, J.; López, R.; Santamaría, P.; Garde-Cerdán, T. Elicitor and nitrogen applications to Garnacha, Graciano and Tempranillo vines: Effect on grape amino acid composition. J. Sci. Food Agric. 2018, 98, 2341–2349. [Google Scholar] [CrossRef]
- Dai, Z.W.; Ollat, N.; Gomès, E.; Decroocq, S.; Tandonnet, J.P.; Bordenave, L.; Pieri, P.; Hilbert, G.; Kapel, C.; Van Leeuwen, C.; et al. Ecophysiological, genetic, and molecular causes of variation in grape berry weight and composition: A review. Am. J. Enol. Vitic. 2011, 62, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo-Togores, J. Tratado de Enología, 2nd ed.; Ediciones Mundi-Prensa: Madrid, Spain, 2011. [Google Scholar]
- Kliewer, W.M. Concentration of Tartrates, Malates, Glucose and Fructose in the Fruits of the Genus Vitis. Am. J. Enol. Vitic. 1967, 18, 91–96. [Google Scholar]
- Conde, C.; Silva, P.; Fontes, N.; Dias, A.C.P.; Tavares, R.M.; Sousa, M.J.; Agasse, A.; Delrot, S.; Gerós, H. Biochemical Changes throughout Grape Berry Development and Fruit and Wine Quality C. Food 2007, 1, 1–22. [Google Scholar]
- Ruffner, P.H.; Possner, D.; Brem, S.; Rast, D. The physiological role of malic enzyme in grape ripening. Planta 1984, 160, 444–448. [Google Scholar] [CrossRef]
- Possner, D.; Ruffner, H.P.; Rast, D.M. Isolation and Biochemical Characterization of Grape Malic Enzyme. Planta 1981, 151, 549–554. [Google Scholar] [CrossRef]
- Texeira, A.; Erias-Dias, J.; Castellarin, S.D.; Gerós, H. Berry Phenolics of Grapevine under Challenging Environments. Int. J. Mol. Sci. 2013, 14, 18711–18739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mira de Orduña, R. Climate change associated effects on grape and wine quality and production. Food Res. Int. 2010, 43, 1844–1855. [Google Scholar] [CrossRef]
- Cincotta, F.; Verzera, A.; Prestia, O.; Tripodi, G.; Lechhab, W.; Sparacio, A.; Condurso, C. Influence of leaf removal on grape, wine and aroma compounds of Vitis vinifera L. cv. Merlot under Mediterranean climate. Eur. Food Res. Technol. 2021, 1–11. [Google Scholar] [CrossRef]
- Barceló, A.R.; Calderón, A.A.; Zapata, J.M.; Muñoz, R. The histochemical localization of anthocyanins in seeded and seedless grapes (Vitis vinifera). Sci. Hortic. 1994, 57, 265–268. [Google Scholar] [CrossRef]
- Buttrose, M.S.; Hale, C.R.; Kliewer, W.M. Effect of Temperature on the Composition of «Cabernet Sauvignon» Berries. Am. J. Enol. Vitic. 1971, 22, 71–75. [Google Scholar]
- Yamane, T.; Jeong, S.T.; Goto-Yamamoto, N.; Koshita, Y.; Kobayashi, S. Effects of Temperature on Anthocyanin Biosynthesis in Grape Berry Skins. Am. J. Enol. Vitic. 2006, 57, 54–59. [Google Scholar]
- He, F.; Mu, L.; Yan, G.L.; Liang, N.N.; Pan, Q.H.; Wang, J.; Reevez, M.J.; Duan, C.Q. Biosynthesis of anthocyanins and their regulation in colored grapes. Molecules 2010, 15, 9057–9091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Núñez, V.; Monagas, M.; Gomez-Cordovés, M.C.; Bartolomé, B. Vitis vinifera L. cv. Graciano grapes characterized by its anthocyanin profile. Postharvest Biol. Technol. 2004, 31, 69–79. [Google Scholar] [CrossRef]
- Arozarena, I.; Ayestarán, B.; Cantalejo, M.; Navarro, M.; Vera, M.; Abril, I.; Casp, A. Anthocyanin composition of Tempranillo, Garnacha and Cabernet Sauvignon grapes from high-and low-quality vineyards over two years. Eur. Food Res. Technol. 2002, 214, 303–309. [Google Scholar] [CrossRef]
- Heinonen, M. Antioxidant activity and antimicrobial effect of berry phenolics–a Finnish perspective. Mol. Nutr. Food Res. 2007, 51, 684–691. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Parameter | Day 0 | Day 5 | Day 10 | Day 15 | Day 20 |
|---|---|---|---|---|---|---|
| Tintilla de Rota | Glucose | 38.50 ± 0.36 a | 38.97 ± 1.17 a | 38.55 ± 0.48 a | 39.14 ± 0.42 a | 38.05 ± 1.84 a |
| Fructose | 36.10 ± 1.40 a | 37.95 ± 1.42 a | 37.40 ± 0.76 a | 38.30 ± 1.04 a | 37.68 ± 4.29 a | |
| Total | 74.59 ± 1.75 a | 76.92 ± 2.54 a | 75.95 ± 1.05 a | 77.44 ± 1.24 a | 75.72 ± 6.12 a | |
| Graciano | Glucose | 38.46 ± 0.43 a | 37.96 ± 1.05 a | 38.37 ± 0.81 a | 38.04 ± 0.77 a | 37.96 ± 1.72 a |
| Fructose | 36.33 ± 0.56 a | 37.44 ± 1.25 a | 40.13 ± 0.44 a | 39.72 ± 1.69 a | 40.18 ± 2.25 a | |
| Total | 74.79 ± 0.85 a | 75.40 ± 2.31 a | 78.49 ± 0.96 a | 77.77 ± 2.42 a | 78.14 ± 3.95 a |
| Parameter | Accessions | Day 0 | Day 5 | Day 10 | Day 15 | Day 20 |
|---|---|---|---|---|---|---|
| Caftaric | Tintilla de Rota | 0.010 ± 0.003 a | 0.026 ± 0.001 a | 0.021 ± 0.002 a | 0.012 ± 0.000 a | 0.077 ± 0.009 b |
| Graciano | 0.140 ± 0.002 a | 0.045 ± 0.003 b | 0.058 ± 0.001 c | 0.081 ± 0.004 a | 0.135 ± 0.009 a | |
| Catechin | Tintilla de Rota | 0.022 ± 0.008 a | 0.009 ± 0.016 a | 0.000 ± 0.000 a | 0.010 ± 0.010 a | 0.008 ± 0.014 a |
| Graciano | nd | nd | nd | nd | 0.040 ± 0.056 | |
| Epicatechin | Tintilla de Rota | 0.173 ± 0.044 a | 0.140 ± 0.058 a | 0.079 ± 0.027 b | 0.099 ± 0.057 c | 0.121 ± 0.020 a |
| Graciano | 0.181 ± 0.049 a | 0.146 ± 0.061 a | 0.096 ± 0.060 b | 0.118 ± 0.046 b | 0.152 ± 0.033 a | |
| Epicatechin gallate | Tintilla de Rota | 0.047 ± 0.003 a | 0.027 ± 0.004 a | 0.023 ± 0.008 a | 0.019 ± 0.015 a | 0.032 ± 0.003 a |
| Graciano | 0.036 ± 0.000 a | 0.025 ± 0.011 a | 0.018 ± 0.003 a | 0.045 ± 0.019 a | 0.029 ± 0.041 a | |
| Q-3-gal | Tintilla de Rota | 0.033 ± 0.004 a | 0.026 ± 0.002 a | 0.013 ± 0.001 a | 0.014 ± 0.002 a | 0.006 ± 0.001 a |
| Graciano | 0.092 ± 0.006 a | 0.068 ± 0.012 a | 0.072 ± 0.004 a | 0.063 ± 0.004 a | 0.043 ± 0.000 b | |
| Rutin | Tintilla de Rota | 0.017 ± 0.003 a | 0.018 ± 0.002 a | 0.008 ± 0.002 a | 0.013 ± 0.001 a | 0.004 ± 0.001 a |
| Graciano | 0.049 ± 0.002 a | 0.026 ± 0.002 a | 0.033 ± 0.001 a | 0.024 ± 0.001 a | 0.029 ± 0.004 a | |
| Q-3-rahm | Tintilla de Rota | 0.013 ± 0.001 a | 0.010 ± 0.000 a | 0.016 ± 0.001 a | 0.018 ± 0.000 a | 0.021 ± 0.001 a |
| Graciano | 0.011 ± 0.002 a | 0.007 ± 0.002 a | 0.019 ± 0.001 a | 0.020 ± 0.001 a | 0.019 ± 0.001 a | |
| Kaempferol | Tintilla de Rota | 1.469 ± 0.013 a | 1.451 ± 0.001 a | 1.449 ± 0.001 a | 1.454 ± 0.003 a | 1.452 ± 0.004 a |
| Graciano | 1.471 ± 0.000 a | 1.467 ± 0.001 a | 1.480 ± 0.004 a | 1.490 ± 0.011 a | 1.480 ± 0.003 a | |
| Quercetin | Tintilla de Rota | 0.137 ± 0.010 a | 0.129 ± 0.004 a | 0.129 ± 0.003 a | 0.132 ± 0.003 a | 0.136 ± 0.006 a |
| Graciano | 0.146 ± 0.001 a | 0.144 ± 0.004 a | 0.155 ± 0.002 a | 0.159 ± 0.011 a | 0.177 ± 0.004 a | |
| Del3Glu | Tintilla de Rota | 0.310 ± 0.027 a | 0.204 ± 0.020 b | 0.176 ± 0.016 b | 0.161 ± 0.019 b | 0.177 ± 0.011 b |
| Graciano | 0.206 ± 0.009 a,b | 0.179 ± 0.011 a | 0.193 ± 0.007 a,b | 0.249 ± 0.009 b,c | 0.215 ± 0.032 a,b | |
| Cy3Glu | Tintilla de Rota | 0.034 ± 0.006 a | 0.025 ± 0.003 a | 0.023 ± 0.002 a | 0.025 ± 0.001 a | 0.039 ± 0.005 a |
| Graciano | 0.016 ± 0.001 a | 0.017 ± 0.002 a | 0.017 ± 0.001 a | 0.024 ± 0.001 a | 0.060 ± 0.009 a | |
| Pet3Glu | Tintilla de Rota | 0.363 ± 0.025 a | 0.249 ± 0.029 b | 0.227 ± 0.012 b | 0.208 ± 0.020 b | 0.220 ± 0.010 b |
| Graciano | 0.253 ± 0.009 a,b | 0.224 ± 0.013 a | 0.253 ± 0.001 a,b | 0.300 ± 0.011 b,c | 0.262 ± 0.038 a,b | |
| Peo3Glu | Tintilla de Rota | 0.468 ± 0.028 a | 0.466 ± 0.054 a | 0.511 ± 0.040 a,b | 0.560 ± 0.049 b | 0.715 ± 0.031 c |
| Graciano | 0.293 ± 0.013a | 0.363 ± 0.016 b,c | 0.383 ± 0.031 c | 0.503 ± 0.007 d | 0.936 ± 0.015 e | |
| Mal3Glu | Tintilla de Rota | 4.494 ± 0.030 a | 3.236 ± 0.026 b,e | 3.132 ± 0.014 c | 3.055 ± 0.030 d | 3.255 ± 0.138 e |
| Graciano | 3.532 ± 0.142 a | 3.315 ± 0.169 b | 3.884 ± 0.056 c,e | 4.571 ± 0.092 d | 3.836 ± 0.051 e | |
| Del36AcG | Tintilla de Rota | 0.032 ± 0.003 a | 0.022 ± 0.002 a | 0.018 ± 0.001 a | 0.017 ± 0.002 a | 0.018 ± 0.002 a |
| Graciano | 0.011 ± 0.000 a | 0.012 ± 0.000 a | 0.013 ± 0.001 a | 0.015 ± 0.001 a | 0.017 ± 0.002 a | |
| Cy36AcG | Tintilla de Rota | 0.020 ± 0.005 a | 0.009 ± 0.001 a | 0.009 ± 0.000 a | 0.011 ± 0.002 a | 0.010 ± 0.000 a |
| Graciano | 0.006 ± 0.000 a | 0.007 ± 0.001 a | 0.006 ± 0.001 a | 0.011 ± 0.002 a | 0.011 ± 0.002 a | |
| Pet36AcG | Tintilla de Rota | 0.036 ± 0.003 a | 0.023 ± 0.002 a | 0.020 ± 0.002 a | 0.017 ± 0.002 a | 0.016 ± 0.000 a |
| Graciano | 0.015 ± 0.000 a | 0.015 ± 0.001 a | 0.014 ± 0.001 a | 0.015 ± 0.001 a | 0.017 ± 0.002 a | |
| Peo36AcG | Tintilla de Rota | 0.720 ± 0.046 a | 0.532 ± 0.034 b,c,d | 0.489 ± 0.030 c,e | 0.494 ± 0.006 d,e | 0.470 ± 0.019 e,f |
| Graciano | 0.416 ± 0.033 a | 0.430 ± 0.019 a,c,e | 0.483 ± 0.007 c,d | 0.519 ± 0.012 d | 0.480 ± 0.007 d,e | |
| Mal36AcG | Tintilla de Rota | 0.008 ± 0.013 a | 0.001 ± 0.000 a | 0.001 ± 0.000 a | 0.001 ± 0.000 a | 0.001 ± 0.000 a |
| Graciano | 0.009 ± 0.007 a | 0.010 ± 0.006 a | 0.010 ± 0.011 a | 0.010 ± 0.001 a | 0.007 ± 0.005 a | |
| Cy36CuG | Tintilla de Rota | 0.019 ± 0.017 a | 0.021 ± 0.004 a | 0.022 ± 0.019 a | 0.016 ± 0.023 a | 0.001 ± 0.000 a |
| Graciano | 0.001 ± 0.000 a | 0.011 ± 0.002 a | 0.023 ± 0.004 a | 0.004 ± 0.003 a | 0.015 ± 0.002 a | |
| Pet36CuG | Tintilla de Rota | 0.015 ± 0.020 a | 0.014 ± 0.002 a | 0.009 ± 0.007 a | 0.009 ± 0.012 a | 0.001 ± 0.000 a |
| Graciano | 0.028 ± 0.006 a | 0.020 ± 0.007 a | 0.018 ± 0.008 a | 0.024 ± 0.003 a | 0.018 ± 0.005 a | |
| Peo36CuG | Tintilla de Rota | 0.027 ± 0.044 a | 0.035 ± 0.007 a | 0.025 ± 0.021 a | 0.030 ± 0.005 a | 0.001 ± 0.000 a |
| Graciano | 0.039 ± 0.007 a | 0.036 ± 0.012 a | 0.032 ± 0.014 a | 0.049 ± 0.002 a | 0.072 ± 0.030 a | |
| T.non-anthocyanin phenols | Tintilla de Rota | 1.921 ± 0.043 a | 1.835 ± 0.071 a | 1.739 ± 0.028 a | 1.772 ± 0.078 a | 1.858 ± 0.015 a |
| Graciano | 2.127 ± 0.060 a | 1.929 ± 0.087 a | 1.931 ± 0.058 a | 2.001 ± 0.067 a | 2.105 ± 0.006 a | |
| T. Anthocyanin | Tintilla de Rota | 6.546 ± 0.264 a | 4.836 ± 0.185 b | 4.663 ± 0.165 b | 4.604 ± 0.171 b | 4.926 ± 0.218 b |
| Graciano | 4.826 ± 0.230 a | 4.638 ± 0.260 a | 5.330 ± 0.144 b | 6.296 ± 0.144 c | 5.944 ± 0.200 c |
| Compounds | F1 | F2 | F3 |
|---|---|---|---|
| Caftaric | 0.896 | −0.119 | 0.153 |
| Catechin | 0.045 | −0.802 | 0.276 |
| Epicatechin | 0.366 | 0.184 | 0.582 |
| Epicatechin gallate | 0.110 | −0.007 | 0.795 |
| Quercetin 3-galactoside | 0.698 | 0.676 | 0.183 |
| Rutin | 0.713 | 0.0537 | 0.207 |
| Quercetin 3-rhamnoside | −0.024 | −0.458 | −0.655 |
| Kaempferol | −0.055 | 0.294 | −0.557 |
| Quercetin | 0.636 | 0.120 | −0.561 |
| Delphinidin 3-O-glycoside | −0.265 | 0.308 | 0.905 |
| Cyanidin 3-O-glycoside | 0.079 | −0.971 | −0.006 |
| Petunidin 3-O-glycoside | −0.338 | 0.527 | 0.746 |
| Peonidin 3-O-glycoside | 0.014 | −0.910 | −0.364 |
| Malvidin 3-O-glycoside | 0.372 | 0.841 | 0.130 |
| Delphinidin 3-O-(6″-acetyl)-glucoside | −0.890 | −0.262 | 0.291 |
| Cyanidin 3-O-(6″-acetyl)-glucoside | −0.700 | −0.454 | 0.369 |
| Petunidin 3-O-(6″-acetyl)-glucoside | −0.843 | -0.028 | 0.423 |
| Peonidin 3-O-(6″-acetyl)-glucoside | −0.942 | 0.059 | 0.040 |
| Malvidin 3-O-(6″-acetyl)-glucoside | 0.615 | 0.564 | 0.275 |
| Cyanidin 3-O-(6″-p-coumaryl)-glucoside | −0.574 | 0.155 | −0.370 |
| Petunidin 3-O-(6″-p-coumaroyl)-glucoside | 0.623 | 0.646 | 0.301 |
| Peonidin 3-O-(6″-p-coumaroyl)-glucoside | 0.585 | 0.023 | 0.076 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez-Cantizano, A.; Sancho-Galán, P.; Barbero, G.F.; Palacios, V.; Amores-Arrocha, A. Analysis of Compounds with Oenological Interest in Somatic Variants of Grapevines. Horticulturae 2022, 8, 22. https://doi.org/10.3390/horticulturae8010022
Jiménez-Cantizano A, Sancho-Galán P, Barbero GF, Palacios V, Amores-Arrocha A. Analysis of Compounds with Oenological Interest in Somatic Variants of Grapevines. Horticulturae. 2022; 8(1):22. https://doi.org/10.3390/horticulturae8010022
Chicago/Turabian StyleJiménez-Cantizano, Ana, Pau Sancho-Galán, Gerardo F. Barbero, Víctor Palacios, and Antonio Amores-Arrocha. 2022. "Analysis of Compounds with Oenological Interest in Somatic Variants of Grapevines" Horticulturae 8, no. 1: 22. https://doi.org/10.3390/horticulturae8010022
APA StyleJiménez-Cantizano, A., Sancho-Galán, P., Barbero, G. F., Palacios, V., & Amores-Arrocha, A. (2022). Analysis of Compounds with Oenological Interest in Somatic Variants of Grapevines. Horticulturae, 8(1), 22. https://doi.org/10.3390/horticulturae8010022