Propagation from Basal Epicormic Meristems Remediates an Aging-Related Disorder in Almond Clones

Abstract
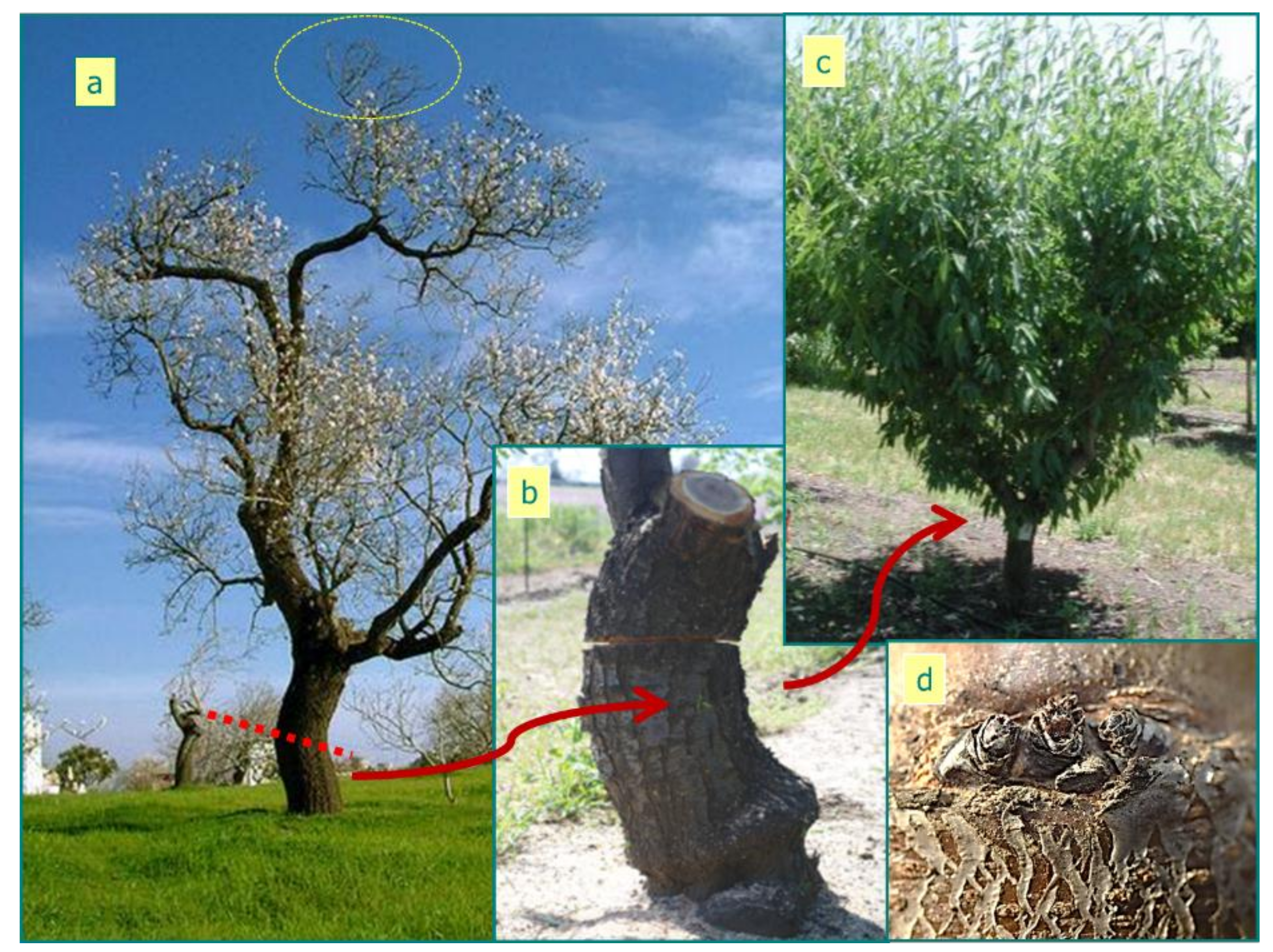
:1. Introduction
2. Materials and Methods
3. Results
3.1. Carmel
3.2. Nonpareil
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Janick, J. The Origins of Fruits, Fruit Growing, and Fruit Breeding. Plant Breed. Rev. 2005, 25, 255–320. [Google Scholar]
- Kester, D.E. The clone in Horticulture. HortScience 1983, 18, 831–837. [Google Scholar]
- Skowcroft, W.R. Somaclonal Variation, the Myth of Clonal Uniformity. In Genetic Flux in Plants; Hohn, B., Dennis, E.S., Eds.; Springer: New York, NY, USA, 1985; pp. 217–245. [Google Scholar]
- Skirvin, R.M.; McPheeters, K.D.; Norton, M. Sources and frequency of somaclonal variation. HortScience 1994, 29, 1232–1237. [Google Scholar] [CrossRef]
- D’Aquila, P.; Rose, G.; Bellizzi, D.; Passarino, G. Epigenetics and aging. Maturitas 2013, 74, 130–136. [Google Scholar] [CrossRef]
- Fraga, M.F.; Rodriguez, R.; Canal, M.J. Genomic DNA methylation-demethylation during aging and reinvigoration of Pinus radiata. Tree Physiol. 2002, 22, 813–816. [Google Scholar] [CrossRef] [PubMed]
- Kester Dale, E.; Asay, R.N. Variability in noninfectious bud-failure of ‘Nonpareil’ almond. Location and environment. J. Am. Soc. Hortic. Sci. 1978, 103, 377–382. [Google Scholar]
- Kester, D.E.; Jones, R.W. Noninfectious bud-failure from breeding programs of almond (Prunus amygdalus Batsch). J. Am Soc. Hortic. Sci. 1970, 74, 214–219. [Google Scholar]
- Fenton, C.A.L.; Kester, D.E.; Kuniyuki, A.H. A model for the development of noninfectious bud failure in vegetatively propagated populations of almond cultivars. Hortscience 1986, 21, 738. [Google Scholar]
- Wilson, E.E.; Schein, R.D. The nature and development of noninfectious bud-failure in almond. Hilgardia 1956, 24, 519–542. [Google Scholar] [CrossRef]
- Almond Board of California. 2018. Available online: http: //www.almonds.com/sites/default/files/Almanac%202018.pdf (accessed on 6 January 2019).
- Kester, D.E.; Shackel, K.A.; Micke, W.C.; Gradziel, T.M.; Viveros, M. Variability in potential and expression of noninfectious bud-failure among nursery propagules of ‘Carmel’ almond. Acta Hortic. 1998, 470, 268–272. [Google Scholar] [CrossRef]
- Fenton, C.A.L.; Kuniyuki, A.H.; Kester, D.E. Search for a viroid etiology for noninfectious bud failure in almond. HortScience 1988, 23, 1050–1053. [Google Scholar]
- Kester, D.E.; Gradziel, T.M. Genetic Disorders. In Almond Production Manual; Micke, W.C., Ed.; University of California: Oakland, CA, USA, 1996; pp. 76–87. [Google Scholar]
- Kester, D.E. Noninfectious Bud-Failure in Almond. In Virus Diseases and Disorders of Stone Fruits in North America; Fulton, R.W., Ed.; Agricultural Research Service, U.S. Department of Agriculture: Washington, DC, USA, 1976; pp. 278–283. [Google Scholar]
- Kester, D.E. Noninfectious bud-failure, a nontransmissable inherited disorder in almond. I. Pattern of phenotypic inheritance. Proc. Am. Soc. Hortic. Sci. 1968, 92, 7–15. [Google Scholar]
- Kester, D.E. Noninfectious bud-failure, a nontransmissable inherited disorder in almond II. Progeny tests for bud-failure. Proc. Am. Soc. Hortic. Sci. 1968, 92, 16–28. [Google Scholar]
- Kester, D.E.; Shackel, K.A.; Micke, W.C.; Viveros, M.; Gradziel, T.M. Noninfectious bud failure in ‘Carmel’ almond, I. Pattern of development in vegetative progeny trees. J. Am. Soc. Hortic. Sci. 2004, 129, 244–249. [Google Scholar] [CrossRef]
- Negrón, C.; Contador, L.; Lampinen, B.D.; Metcalf, S.G.; Guédon, Y.; Costes, E.; DeJong, T.M. Differences in proleptic and epicormic shoot structures in relation to water deficit and growth rate in almond trees (Prunus dulcis). Ann Bot. 2014, 113, 545–554. [Google Scholar] [CrossRef]
- Fontaine, F.; Kiefer, E.; Clemen, T.C.; Burrus, M.; Druelle, J.L. Ontogeny of the proventitious epicormic buds in Quercus petraea. II. From 6 to 40 years of the tree’s life. Trees Struct. Funct. 1999, 14, 83–90. [Google Scholar]
- Meier, A.R.; Saunders, M.R.; Michler, C.H. Epicormic buds in trees: A review of bud establishment, development and dormancy release. Tree Physiol. 2012, 32, 565–584. [Google Scholar] [CrossRef]
- Preece, J.E. Stock Plant Physiological Factors Affecting Growth and Morphogenesis. In Plant Propagation by Tissue Culture, 3rd ed.; George, E.F., Hall, M.A., de Klerk, G.J., Eds.; Springer: Dordrecht, The Netherlands, 2008; Volume 1, p. 507. [Google Scholar]
- Fishel, D.W.; Zaczek, J.J.; Preece, J.E. Positional influence on rooting of shoots forced from the main bole in swamp white oak and northern red oak. Can. J. For. Res. 2003, 33, 705–711. [Google Scholar] [CrossRef]
- Foundation Plant Services. 2019. Available online: http://fps.ucdavis.edu/index.cfm (accessed on 6 January 2019).
- Fresnedo-Ramírez, J.; Chan, H.M.; Parfitt, D.E.; Crisosto, C.H.; Gradziel, T.M. Genome-wide DNA-(de)methylation is associated with Noninfectious Bud-failure exhibition in Almond (Prunus dulcis [Mill.] D.A.Webb). Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Kester, D.E. Noninfectious bud-failure, a nontransmissable inherited disorder in almonds, III. Variability in BF potential within plants. J. Am. Soc. Hortic. Sci. 1970, 95, 162–165. [Google Scholar]
- Gradziel, T.M. Almond origin and domestication. In Horticultural Reviews; Janick, J., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 23–82. [Google Scholar]
- Kester, D.E. Inheritance of noninfectious bud-failure in almond. Proc. Am. Soc. Hortic. Sci. 1961, 77, 278–285. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Source | FPS ID 1 | Origin |
|---|---|---|
| E1 | na | Delta-mother tree propagated from original Carmel seedling tree |
| E2 | 3-56-1-90 | Delta source tree 2 propagated basal epicormic bud from Delta tree |
| E3 | 3-56-8-92 | Delta source tree 13 propagated basal epicormic bud from Delta tree |
| E4 | 3-56-2-90 | Delta source tree 7 propagated basal epicormic bud from Delta tree |
| C1 | na | Standard commercial nursery source established in 1988 |
| C2 | na | Standard commercial nursery source established in 1986 |
| C3 | na | Standard commercial nursery source established in 1985 |
| C4 | na | Standard commercial nursery source established in 1981 |
| Source | FPS ID 1 | Origin |
|---|---|---|
| E1 | 3-8-6-72 | Propagated from a basal epicormic bud of a 108-year-old relic tree |
| E2 | 3-8-17-92 | Propagated from a basal epicormic bud of a 100+ year-old relic tree |
| E3 | 3-8-2-70 | Propagated from a basal epicormic bud of a 100+ year-old heat-treated relic tree |
| E4 | na | Commercial nursery propagation source originating from 100+ year-old relic tree |
| C1 | na | Standard commercial nursery source established in 1988 |
| C2 | na | Standard commercial nursery source established in 1986 |
| C3 | na | Standard commercial nursery source established in 1984 |
| C4 | na | Standard commercial nursery source established in 1977 |
| E1 | E2 | E3 | E4 | C1 | C2 | C3 | C4 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | Bf Trees | Per-cent | Bf Trees | Per-cent | Bf Trees | Per-cent | Bf Trees | Per-cent | Bf Trees | Per-cent | Bf Trees | Per-cent | Bf Trees | Per-cent | Bf Trees | Per-cent |
| 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 31 | 10.7 | 68 | 36.0 |
| 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 36 | 12.4 | 147 | 77.8 |
| 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 60 | 14.7 | 53 | 22.5 | 102 | 35.1 | 183 | 96.8 |
| 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 71 | 17.4 | 66 | 28.0 | 114 | 39.2 | 189 | 100 |
| 6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 114 | 28.0 | 102 | 43.2 | 156 | 53.6 | 189 | 100 |
| 7 | 0 | 0 | 0 | 0 | 0 | 0 | 152 | 51.2 | 145 | 35.6 | 181 | 76.7 | 199 | 68.4 | 189 | 100 |
| 8 | 1 | 3.2 | 11 | 5.1 | 25 | 10.7 | 221 | 74.4 | 166 | 40.8 | 218 | 92.4 | 235 | 80.8 | 189 | 100 |
| 9 | 2 | 6.5 | 15 | 6.9 | 31 | 13.2 | 238 | 80.1 | 197 | 48.4 | 229 | 97.0 | 286 | 98.3 | 189 | 100 |
| 10 | 4 | 12.9 | 17 | 7.8 | 34 | 14.5 | 264 | 88.9 | 213 | 52.3 | 236 | 100 | 291 | 100 | 189 | 100 |
| Total | 31 | 217 | 234 | 297 | 407 | 236 | 291 | 189 | ||||||||
| E1 | E2 | E3 | E4 | C1 | C2 | C3 | C4 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | BF Trees | Per-cent | BF Trees | Per-cent | BF Trees | Per-cent | BF Trees | Per-cent | BF Trees | Per-cent | BF Trees | Per-cent | BF Trees | Per-cent | BF Trees | Per-cent |
| 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1.7 |
| 7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 3.3 |
| 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1.7 | 3 | 5.0 |
| 9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1.7 | 1 | 1.7 | 2 | 3.3 | 7 | 11.7 |
| 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1.7 | 2 | 3.3 | 3 | 5.0 | 13 | 21.7 |
| 11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 3.3 | 4 | 6.7 | 6 | 10.0 | 22 | 36.7 |
| 12 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 5.0 | 7 | 11.7 | 10 | 16.7 | 28 | 46.7 |
| 13 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 6.7 | 10 | 16.7 | 14 | 23.3 | 40 | 66.7 |
| 14 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 9 | 15.0 | 15 | 25.0 | 17 | 28.3 | 44 | 73.3 |
| 15 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 11 | 18.3 | 18 | 30.0 | 20 | 33.3 | 48 | 80.0 |
| 16 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 14 | 23.3 | 21 | 35.0 | 26 | 43.3 | 50 | 83.3 |
| 17 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 20 | 33.3 | 23 | 38.3 | 28 | 46.7 | 52 | 86.7 |
| 18 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 5 | 21 | 35.0 | 25 | 41.7 | 34 | 56.7 | 53 | 88.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gradziel, T.; Lampinen, B.; Preece, J.E. Propagation from Basal Epicormic Meristems Remediates an Aging-Related Disorder in Almond Clones. Horticulturae 2019, 5, 28. https://doi.org/10.3390/horticulturae5020028
Gradziel T, Lampinen B, Preece JE. Propagation from Basal Epicormic Meristems Remediates an Aging-Related Disorder in Almond Clones. Horticulturae. 2019; 5(2):28. https://doi.org/10.3390/horticulturae5020028
Chicago/Turabian StyleGradziel, Thomas, Bruce Lampinen, and John E. Preece. 2019. "Propagation from Basal Epicormic Meristems Remediates an Aging-Related Disorder in Almond Clones" Horticulturae 5, no. 2: 28. https://doi.org/10.3390/horticulturae5020028
APA StyleGradziel, T., Lampinen, B., & Preece, J. E. (2019). Propagation from Basal Epicormic Meristems Remediates an Aging-Related Disorder in Almond Clones. Horticulturae, 5(2), 28. https://doi.org/10.3390/horticulturae5020028