Water-Related Variables for Predicting Yield of Apple under Deficit Irrigation


Abstract
:
1. Introduction
2. Materials and Methods
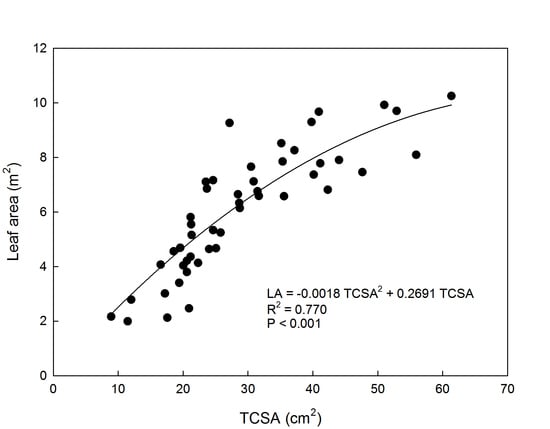
3. Results and Discussion
Funding
Conflicts of Interest
References
- Hamon, R.W. Evapotranspiration and water yield predictions. In Evapotranspiration and its role in water resources management. Conf. Proc. Am. Soc. Agric. Eng. 1966, December, 8–9. [Google Scholar]
- Shockley, D.G. Evapotranspiration and farm irrigation planning and management. In Evapotranspiration and its role in water resources management. Conf. Proc. Am. Soc. Agric. Eng. 1966, December, 3–5. [Google Scholar]
- Hansen, V.E. Evapotranspiration and water resources management. Conf. Proc. Am. Soc. Agric. Eng. 1966, December, 12–13. [Google Scholar]
- Kang, S.; Gu, B.; Du, T.; Zhang, J. Crop coefficient and ratio of transpiration to evapotranspiration of winter wheat and maize in a semi-humid region. Agric. Water Manag. 2003, 59, 239–254. [Google Scholar] [CrossRef]
- Xu, L.K.; Hsiao, T.C. Predicted versus measured photosynthetic water-use efficiency of crop stands under dynamically changing field environments. J. Exp. Bot. 2004, 55, 2395–2411. [Google Scholar] [CrossRef] [Green Version]
- Kijne, J.W.; Barker, R.; Molden, D. Water Productivity in Agriculture: Limits and Opportunities for Improvement; CABI Publishing: Wallingford, UK, 2003; 332p. [Google Scholar]
- Chalmers, D.J.; Mitchell, P.D.; Vanheek, L. Control of peach tree growth and productivity by regulated water supply, tree density and summer pruning. J. Am. Soc. Hortic. Sci. 1981, 106, 307–397. [Google Scholar]
- Chalmers, D.J.; Burge, G.; Jerie, P.H.; Mitchell, P.D. The mechanism of regulation of Bartlett pear fruit and vegetative growth by irrigation withholding and regulated deficit irrigation. J. Am. Soc. Hortic. Sci. 1986, 111, 904–907. [Google Scholar]
- English, M.J. Deficit irrigation: Analytical framework. J. Irrig. Drain. Eng. 1990, 116, 399–412. [Google Scholar] [CrossRef]
- Mitchell, P.D.; Jerie, P.H.; Chalmers, D.J. Effects of regulated water deficits on pear tree growth, flowering, fruit growth and yields. J. Am. Soc. Hortic. Sci. 1984, 109, 604–606. [Google Scholar]
- Zwart, S.J.; Bastiaanssen, W.G. Review of measured crop water productivity values for irrigated wheat, rice, cotton and maize. Agric. Water Manag. 2004, 69, 115–133. [Google Scholar] [CrossRef]
- Fan, T.; Wang, S.; Xiaoming, T.; Luo, J.; Stewart, B.A.; Gao, Y. Grain yield and water use in a long-term fertilization trial in Northwest China. Agric. Water Manag. 2005, 76, 36–52. [Google Scholar] [CrossRef]
- Fereres, E.; Goldhamer, D.A.; Parsons, L.R. Irrigation water management of horticultural crops. HortScience 2003, 38, 1036–1042. [Google Scholar]
- Johnson, R.S.; Handley, D.F. Using water stress to control vegetative growth and productivity of temperate fruit trees. HortScience 2000, 35, 1048–1050. [Google Scholar]
- Dry, P.R.; Loveys, B.R.; Botting, D.G.; Düring, H. Effects of partial root-zone drying on grapevine vigour, yield, composition of fruit and use of water. In Proceedings of the Ninth Australian Wine Industry Technical Conference, Adelaide, South Australia, 16–19 July 1995; pp. 128–131. [Google Scholar]
- Lo Bianco, R. Responses of apple to partial root-zone drying. A review. In Irrigation Management, Technologies, and Environmental Impacts; Ali, M.H., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2013; Chapter 3; pp. 71–86. [Google Scholar]
- Dodd, I.C.; Egea, G.; Davies, W.J. Abscisic acid signalling when soil moisture is heterogeneous: Decreased photoperiod sap flow from drying roots limits ABA export to the shoots. Plant Cell Environ. 2008, 31, 1263–1274. [Google Scholar] [CrossRef] [PubMed]
- Gowing, D.J.G.; Davies, W.J.; Jones, H.G. A positive root-sourced signal as an indicator of soil drying in apple, Malus × domestica Borkh. J. Exp. Bot. 1990, 41, 1535–1540. [Google Scholar] [CrossRef]
- Dry, P.R.; Loveys, B.R.; Düring, H. Partial drying of the root-zone of grape. I. Transient changes in shoot growth and gas exchange. Vitis 2000, 39, 3–7. [Google Scholar]
- Cowan, I.R.; Farquhar, G.D. Stomatal function in relation to leaf metabolism and environment. Symp. Soc. Exp. Biol. 1977, 31, 471. [Google Scholar] [PubMed]
- Davies, W.J.; Zhang, J. Root signals and the regulation of growth and development of plants in drying soil. Ann. Rev. Plant Biol. 1991, 42, 55–76. [Google Scholar] [CrossRef]
- Doorenbos, J.; Kassam, A.H. Yield response to water. In Irrigation and Drainage Paper No. 33; FAO: Rome, Italy, 1986. [Google Scholar]
- Howel, T.A.; Musick, J.T. Relationship of dry matter production of field crops to water consumption. In Crop Water Requirements; Perrier, A., Riou, C., Eds.; INRA: Paris, France, 1985; pp. 247–269. [Google Scholar]
- Ouda, S.A.; Khalil, F.A.; Tantawy, M.M. Predicting the impact of water stress on the yield of different maize hybrids. Res. J. Agric. Biol. Sci. 2006, 2, 369–374. [Google Scholar]
- Lo Bianco, R.; Francaviglia, D. Comparative responses of ‘Gala’ and ‘Fuji’ apple trees to deficit irrigation: Placement versus volume effects. Plant Soil 2012, 357, 41–58. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop evapotranspiration—Guidelines for computing crop water requirements. In Irrigation and Drainage Paper 56; FAO: Rome, Italy, 1998. [Google Scholar]
- García-Tejero, I.; Romero-Vicente, R.; Jiménez-Bocanegra, J.A.; Martínez-García, G.; Durán-Zuazo, V.H.; Muriel-Fernández, J.L. Response of citrus trees to deficit irrigation during different phenological periods in relation to yield, fruit quality, and water productivity. Agric. Water Manag. 2010, 97, 689–699. [Google Scholar] [CrossRef]
- Myers, B.J. Water stress integral—A link between short term stress and long term growth. Tree Physiol. 1988, 4, 315–323. [Google Scholar] [CrossRef]
- Lo Bianco, R.; Policarpo, M.; Scariano, L. Effect of rootstock vigor and in-row spacing on stem and root growth, conformation, and dry matter distribution of young apple trees. J. Hortic. Sci. Biotechnol. 2003, 78, 828–836. [Google Scholar] [CrossRef]
- Westwood, M.N.; Roberts, A.N. The relationship between trunk cross-sectional area and weight of apple trees. J. Am. Soc. Hortic. Sci. 1970, 95, 28–30. [Google Scholar]
- Marsal, J.; Mata, M.; Del Campo, J.; Arbones, A.; Vallverdú, X.; Girona, J.; Olivo, N. Evaluation of partial root-zone drying for potential field use as a deficit irrigation technique in commercial vineyards according to two different pipeline layouts. Irrig. Sci. 2008, 26, 347–356. [Google Scholar] [CrossRef]
- Mossad, A.; Scalisi, A.; Lo Bianco, R. Growth and water relations of field-grown ‘Valencia’ orange trees under long-term partial rootzone drying. Irrig. Sci. 2018, 36, 9–24. [Google Scholar] [CrossRef]
- Naor, A.; Naschitz, S.; Peres, M.; Gal, Y. Responses of apple fruit size to tree water status and crop load. Tree Physiol. 2008, 28, 1255–1261. [Google Scholar] [CrossRef] [Green Version]
- Davies, F.S.; Lakso, A.N. Diurnal and seasonal changes in leaf water potential components and elastic properties in response to water stress in apple trees. Physiol. Plant. 1979, 46, 109–114. [Google Scholar] [CrossRef]
- Olien, W.C.; Lakso, A.N. Effect of rootstock on apple (Malus domestica) tree water relations. Physiol. Plant. 1986, 67, 421–430. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| CI | PRD | CDI | |||||
|---|---|---|---|---|---|---|---|
| Yield (kg/tree) | 36.3 | ab z | 39.4 | a | 33.5 | b | * y |
| Yield Efficiency (kg cm−2) | 0.729 | ab | 0.853 | a | 0.607 | b | ** |
| GRtrunk (cm2 cm−2) | 0.051 | a | 0.052 | a | 0.037 | b | * |
| GRfruit (mm mm−1) | 0.541 | 0.545 | 0.501 | ns | |||
| Leaf Area (m2) | 9.02 | 8.70 | 9.39 | ns | |||
| Vapor Pressure Deficit (kPa) | 158 | 159 | 159 | ns | |||
| LWSDint x | 10.1 | a | 10.5 | ab | 11.1 | b | * |
| Gsint w (mol m−2 s−1) | 16.0 | a | 15.5 | a | 12.5 | b | ** |
| Tr v (mol m−2 s−1) | 25.6 | a | 24.9 | a | 20.0 | b | ** |
| GRtrunk/Tr | 0.022 | 0.023 | 0.019 | ns | |||
| GRfruit/Tr | 0.025 | 0.025 | 0.028 | ns | |||
| Yield/Trtree | 0.177 | b | 0.210 | a | 0.191 | ab | * |
| Trtree u (mol s−1/tree) | 229 | 216 | 188 | ns | |||
| Gstree u (mol s−1/tree) | 143 | 136 | 118 | ns | |||
- z
- Mean separation within rows by Tukey’s multiple comparison test at P < 0.05
- y
- Level of statistical significance for irrigation factor from analysis of variance: ns, P > 0.05; *, P < 0.05; **, P < 0.01; ***, P < 0.001.
- x
- LWSDint is leaf water saturation deficit integrated across the irrigation period.
- w
- Gsint is stomatal conductance integrated across the irrigation period.
- v
- Tr is transpiration integrated across the irrigation period.
- u
- Seasonal integrals of total conductance (GStree) and transpiration (Trtree) per tree.
| CI | PRD | CDI | |||||
|---|---|---|---|---|---|---|---|
| Yield (kg/tree) | 19.3 | 18.5 | 17.2 | ns z | |||
| YieldEff (kg cm−2) | 0.600 | 0.659 | 0.537 | ns | |||
| GRtrunk (cm2 cm−2) | 0.082 | a y | 0.071 | a | 0.053 | b | *** |
| GRfruit (mm mm−1) | 0.545 | 0.568 | 0.563 | ns | |||
| Leaf Area (m2) | 6.69 | 6.31 | 6.95 | ns | |||
| Vapor Pressure Deficit (kPa) | 169 | 168 | 170 | ns | |||
| LWSDint x | 7.53 | a | 8.28 | ab | 8.93 | b | * |
| Gsint w (mol m−2 s−1) | 13.9 | 12.9 | 12.4 | ns | |||
| Tr v (mol m−2 s−1) | 23.7 | 21.0 | 22.0 | ns | |||
| GRtrunk/Tr | 0.044 | a | 0.044 | a | 0.029 | b | * |
| Grfruit/Tr | 0.028 | 0.033 | 0.028 | ns | |||
| Yield/Trtree | 0.141 | ab | 0.161 | a | 0.124 | b | * |
| Trtree u (mol s−1/tree) | 154 | 131 | 150 | ns | |||
| Gstree u (mol s−1/tree) | 90.5 | 77.4 | 88.5 | ns | |||
- z
- Level of statistical significance for irrigation factor from analysis of variance: ns, P > 0.05; *, P < 0.05; **, P < 0.01; ***, P < 0.001.
- y
- Mean separation within rows by Tukey’s multiple comparison test at P < 0.05
- x
- LWSDint is leaf water saturation deficit integrated across the irrigation period.
- w
- Gsint is stomatal conductance integrated across the irrigation period.
- v
- Tr is transpiration integrated across the irrigation period.
- u
- Seasonal integrals of total conductance (GStree) and transpiration (Trtree) per tree.
| Yield = 28.5 + (2774 × GRtrunk/Tr) − (295 × GRfruit/Tr) + (0.070 × Gstree) | |||||
|---|---|---|---|---|---|
| N = 65 | R = 0.554 | R2 = 0.306 | SE of Estimate = 8.85 | P < 0.001 | |
| Parameters | Coefficient | SE | t | P | % of SSreg |
| Constant | 28.5 | 6.74 | 4.23 | <0.001 | - |
| GRtrunk/Tr z | 2774 | 973 | 2.85 | 0.006 | 3.1 |
| GRfruit/Tr y | −295 | 118 | −2.51 | 0.015 | 77.9 |
| Gstree x | 0.070 | 0.031 | 2.26 | 0.027 | 19.0 |
- z
- Trunk growth/transpiration.
- y
- Fruit growth/transpiration.
- x
- Seasonal integral of total conductance (Gs) per tree.
| Yield = −0.019 + (1.35 × LWSDint) + (0.085 × Gstree) | |||||
|---|---|---|---|---|---|
| N = 68 | R = 0.410 | R2 = 0.168 | SE of Estimate = 6.33 | P = 0.003 | |
| Parameters | Coefficient | SE | t | P | % of SSreg |
| Constant | −0.019 | 5.50 | −0.003 | 0.997 | - |
| LWSDint z | 1.35 | 0.52 | 2.59 | 0.012 | 20.5 |
| Gstree y | 0.085 | 0.026 | 3.23 | 0.002 | 79.5 |
- z
- LWSDint is leaf water saturation deficit integrated across the irrigation period.
- y
- Seasonal integral of total conductance (Gs) per tree.
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lo Bianco, R. Water-Related Variables for Predicting Yield of Apple under Deficit Irrigation. Horticulturae 2019, 5, 8. https://doi.org/10.3390/horticulturae5010008
Lo Bianco R. Water-Related Variables for Predicting Yield of Apple under Deficit Irrigation. Horticulturae. 2019; 5(1):8. https://doi.org/10.3390/horticulturae5010008
Chicago/Turabian StyleLo Bianco, Riccardo. 2019. "Water-Related Variables for Predicting Yield of Apple under Deficit Irrigation" Horticulturae 5, no. 1: 8. https://doi.org/10.3390/horticulturae5010008
APA StyleLo Bianco, R. (2019). Water-Related Variables for Predicting Yield of Apple under Deficit Irrigation. Horticulturae, 5(1), 8. https://doi.org/10.3390/horticulturae5010008