Mechanism of the Alternaria alternata Pathogenicity in ‘Fortune’ Mandarin

Abstract
:1. Introduction
2. Materials and Methods
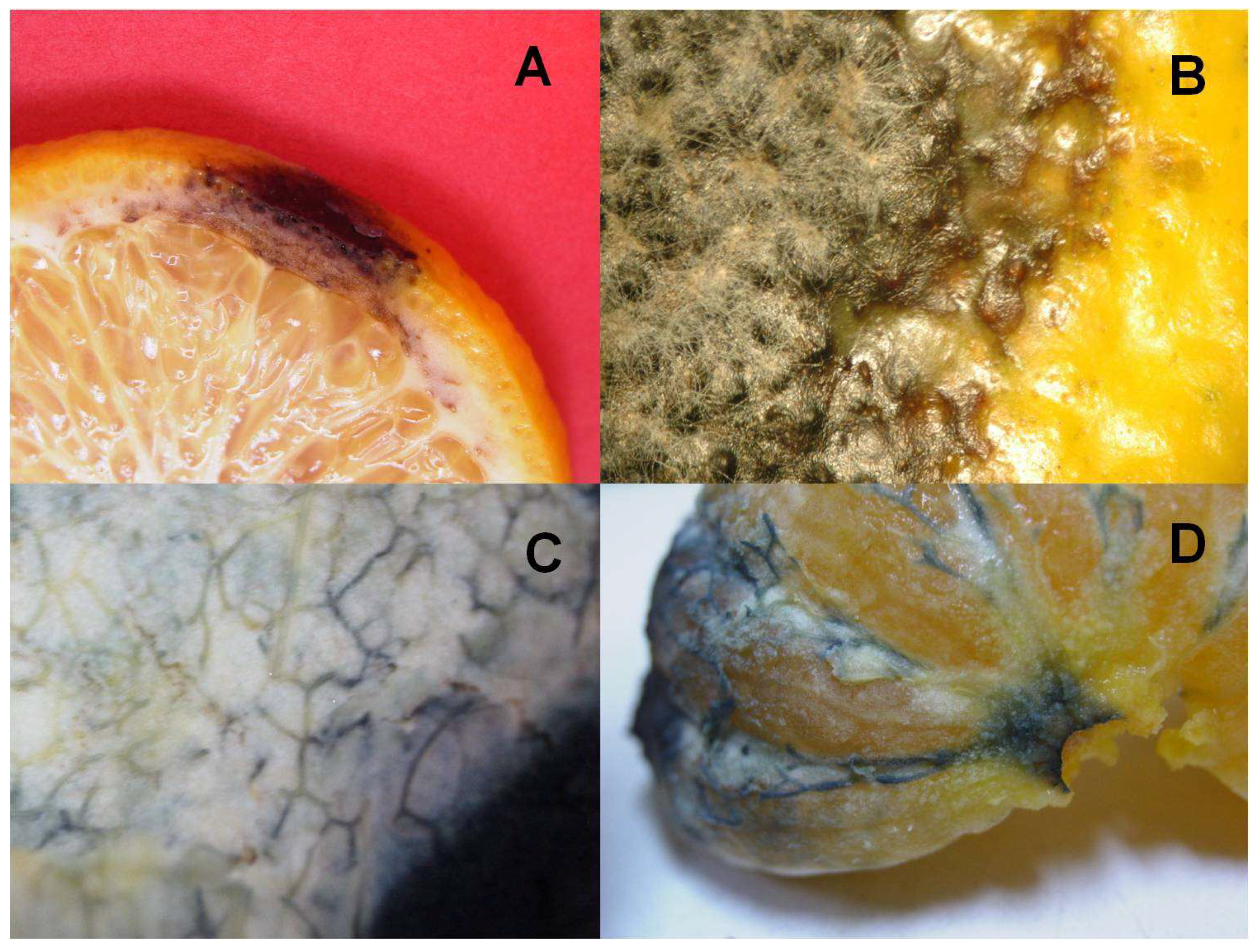
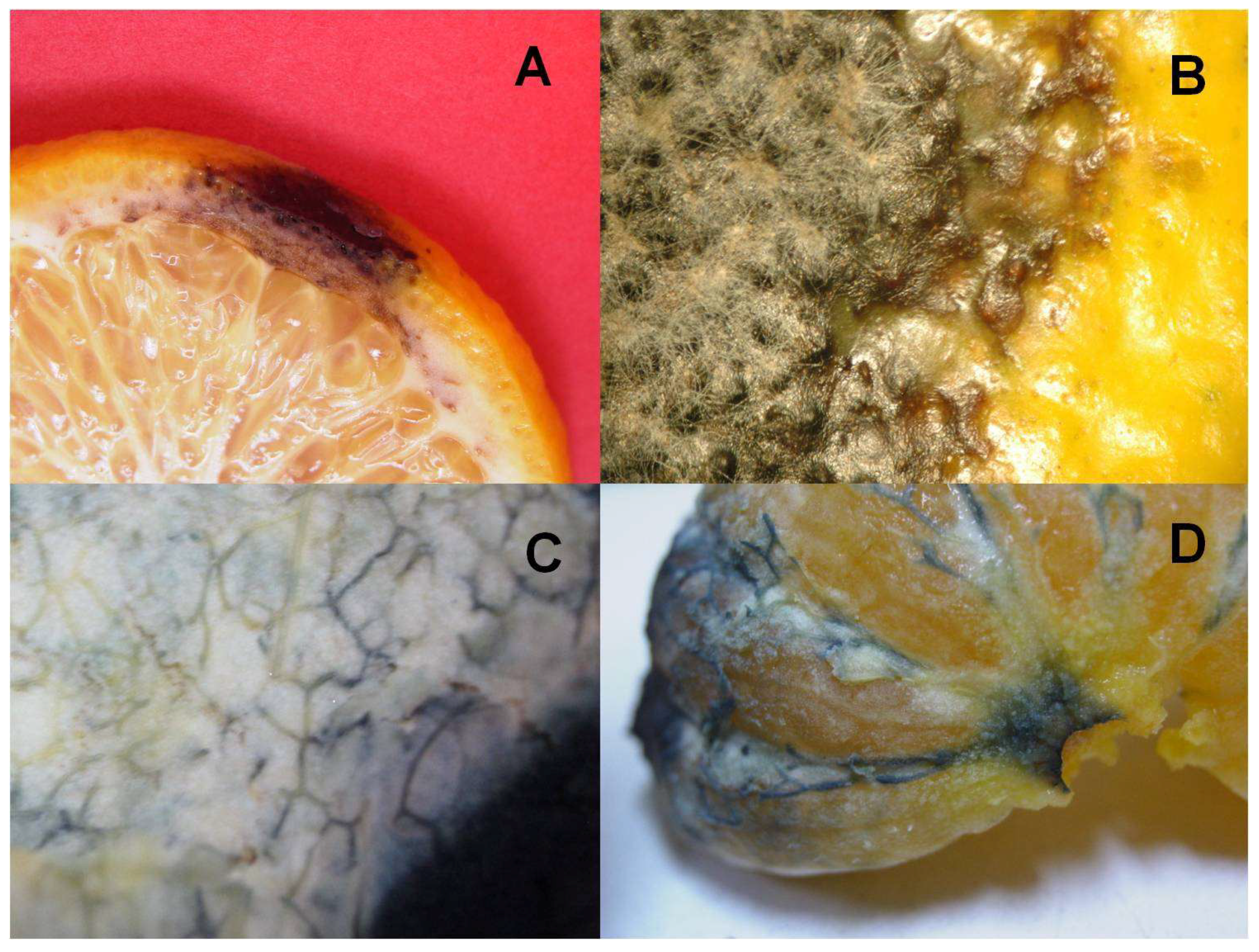
2.1. Selection of Alternaria alternata Isolates, Culture of the Fungus, and Pathogenicity Studies
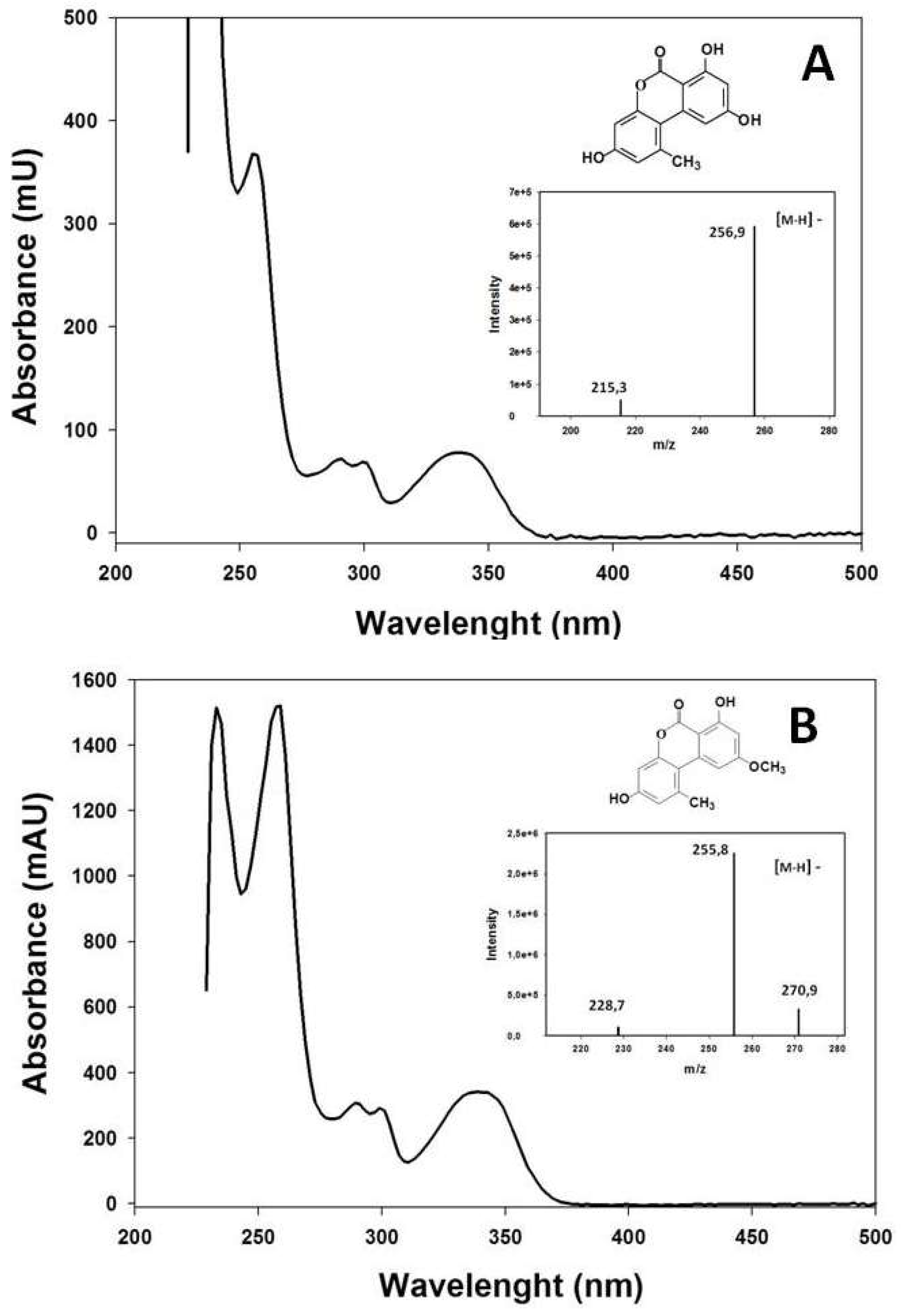
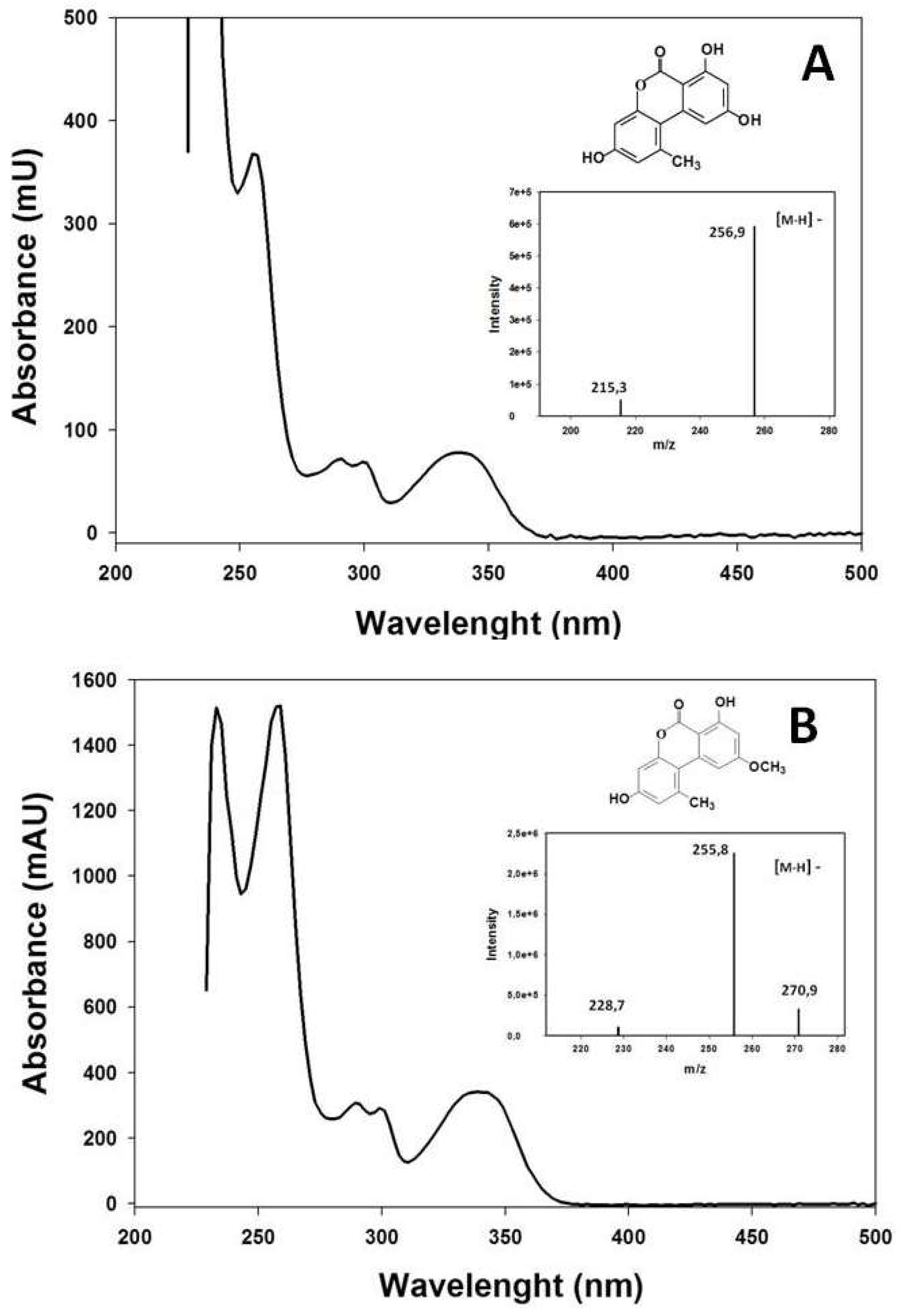
2.2. Extraction and Measurement of A. alternata Mycotoxins
2.3. Extraction and Measurement of Laccase Activity in the Mycelium of A. alternata
2.4. Histological Localization of the Laccase Acitvity
2.5. Statistical Analysis
3. Results
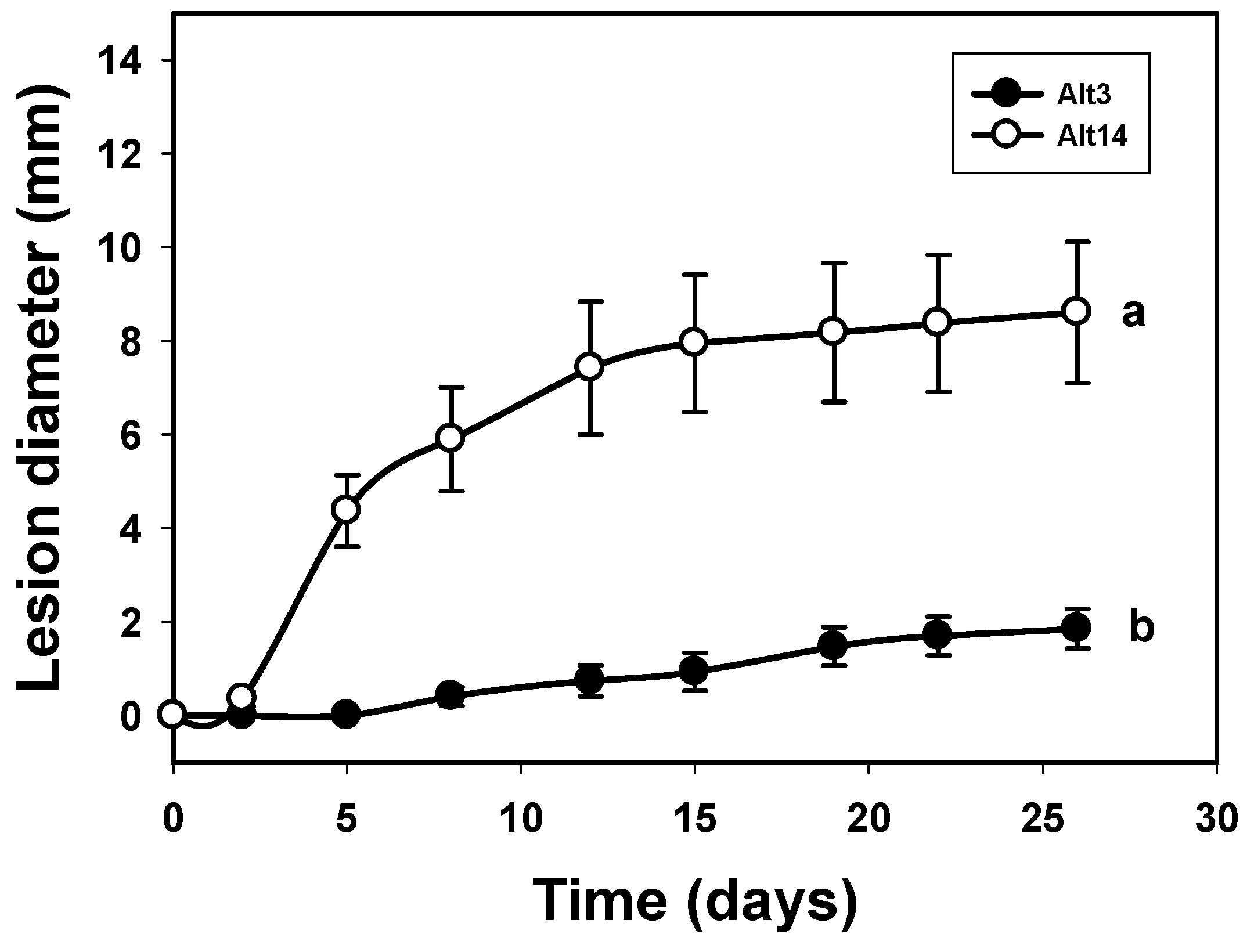
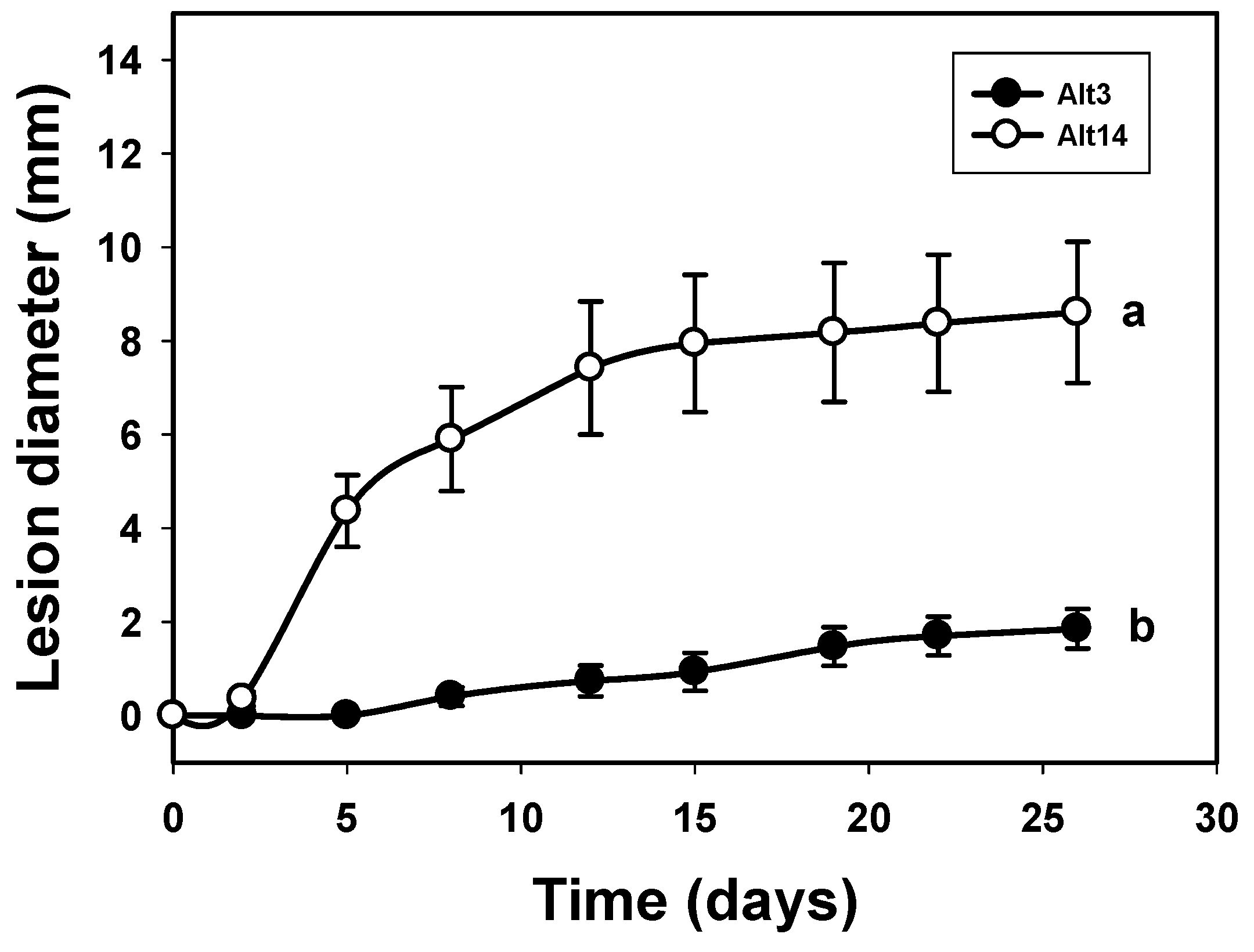
3.1. Study of the Pathogenicity and the Mycotoxins Content of the A. alternata Isolates
3.2. Correlation between the Laccase Activity of A. alternata Isolates and Their Pathogenicity
3.3. Location of Laccase Activity in the Tissues of ‘Fortune’ Fruits Inoculated with A. alternata
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Pegg, K.G. Studies of a strain of Alternaria citri Pierce, the casual organism of brown spot of Emperor mandarin. Queensland J. Agric. Anim. Sci. 1966, 23, 15–18. [Google Scholar]
- Gardner, J.M.; Kono, Y.; Chandler, J.L. Bioassay and host-selectivity of Alternaria citri toxins affecting rouge lemon and mandarins. Physiol. Mol. Plant Pathol. 1986, 29, 293–304. [Google Scholar] [CrossRef]
- Kohmoto, K.; Akimitsu, K.; Otani, H. Correlation of resistance and susceptibility of citrus to Alternaria alternata with sensitivity to host-specific toxins. Phytopathology 1991, 81, 719–722. [Google Scholar] [CrossRef]
- Reis, R.F.; De Almedia, T.F.; Stuchi, E.S.; De Goes, A. Susceptibility of citrus species to Alternaria alternata, the causal agent to the Alternaria brown spot. Sci. Hortic. 2007, 113, 336–342. [Google Scholar] [CrossRef]
- Nemsa, I.; Hernández, M.A.; Lacasa, A.; Porras, I.; García-Lidón, A.; Cifuentes, D.; Bouzid, S.; Ortuño, A.; Del Río, J.A. Pathogenicity of Alternaria alternata on fruits and leaves of ‘Fortune’ mandarin (Citrus clementina × Citrus tangerina). Can. J. Plant Pathol. 2012, 34, 195–202. [Google Scholar] [CrossRef]
- Ortuño, A.; Nemsa, I.; Álvarez, N.; Lacasa, A.; Porras, I.; García-Lidón, A.; Del Río, J.A. Correlation of ethylene synthesis in Citrus fruits and their susceptibility to Alternaria alternata pv. citri. Physiol. Mol. Plant Pathol. 2008, 72, 162–166. [Google Scholar] [CrossRef]
- Ortuño, A.; Del Río, J.A. Role of Citrus phenolic compounds in the resistance mechanism against pathogenic fungi. In Tree and Forestry Science and Biotechnology; Tennant, P., Benkeblia, N., Eds.; Global Science Books, Ltd.: London, UK, 2009; Volume 3, pp. 49–53. ISBN 978-4-903313-25-2. [Google Scholar]
- Díaz, L.; Del Río, J.A.; Pérez-Gilabert, M.; Ortuño, A. Involvement of an extracelular fungus laccase in the flavonoid metabolism in Citrus fruits inoculated with Alternaria alternata. Plant Physiol. Biochem. 2015, 89, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Logrieco, A.; Visconti, A. Mandarin fruit rot caused by Alternaria alternata and associated mycotoxins. Plant Dis. 1990, 74, 415–417. [Google Scholar] [CrossRef]
- Ostry, V. Alternaria mycotoxins: An overview of chemical characterization, producers, toxicity, analysis and occurrence in foodstuffs. World Mycotoxin J. 2008, 1, 175–188. [Google Scholar] [CrossRef]
- Kohmoto, K.; Scheffer, R.P.; Whiteside, J.O. Host-selective toxins from Alternaria citri. Phytopathology 1979, 69, 667–671. [Google Scholar] [CrossRef]
- Gardner, J.M.; Kono, Y.; Tatum, J.H.; Suzuki, Y.; Takeuchi, S. Structure of major component of ACRL toxins, host-specific phytotoxic compound produced by Alternaria citri. Agric. Biol. Chem. 1985, 49, 1235–1238. [Google Scholar]
- Kono, Y.; Gardner, J.M.; Takeuchi, S. Structure of the host-selective toxins produced by a pathotype of Alternaria citri causing brown spot disease of mandarins. Agric. Biol. Chem. 1986, 50, 801–804. [Google Scholar] [CrossRef]
- Nakatsuka, S.; Goto, T.; Kohmoto, K.; Nishimura, S. Natural Products and Biological Activities; Imura, H., Ed.; University of Tokyo Press: Tokyo, Japan, 1986; pp. 11–18. ISBN 978-4-13-068118-6. [Google Scholar]
- Visconti, A.; Bottalico, A.; Solfrizzo, M. Activity of Alternaria alternata metabolites on tomato leaves and Geotrichum candidum. In Phytotoxins and Plant Patogénesis; Graniti, A., Durban, R., Ballio, A., Eds.; Springer: Berlin, Germany, 1989; p. 457. ISBN 978-3-642-73178-5. [Google Scholar]
- Akimitsu, K.; Peever, T.L.; Timmer, L.W. Molecular, ecological and evolutionary approaches to understanding Alternaria diseases of citrus. Mol. Plant Pathol. 2003, 4, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, G.; Giardina, P.; Bianco, C.; Fontanella, B.; Sannia, G. Cooper Induction of laccase isoenzymes in the ligninolytic fungus Pleurotus ostreatus. Appl. Environ. Microb. 2000, 66, 920–924. [Google Scholar] [CrossRef]
- Niku-Paavola, M.L.; Karhunen, E.; Salola, P.; Raunio, V. Ligninolytic enzymes of the White-rot fungus Phlebiaradiata. Biochem. J. 1988, 254, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Stinson, E.E.; Osman, S.F.; Heisler, E.G.; Siciliano, J.; Bills, D.D. Mycotoxin production in whole tomatoes, apples, oranges, and lemons. J. Agric. Food Chem. 1981, 29, 790–792. [Google Scholar] [CrossRef] [PubMed]
- Magnani, R.F.; De Souza, G.D.; Rodrigues-Filho, E. Analysis of Alternariol and Alternariol Monomethyl Ether on Flavedo and Albedo Tissues of Tangerines (Citrus reticulata) with Symptoms of Alternatia Brown Spot. J. Agric. Food Chem. 2007, 55, 4980–4986. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. alternata Isolate | Mycotoxin (µmol/g DW) | Laccase Activity (mU/mL) | |
|---|---|---|---|
| AME | AOH | ||
| Alt3 | 32 ± 2 b | 3 ± 0.2 b | 105 ± 21 b |
| Alt14 | 58 ± 3 a | 28 ± 4 a | 215 ± 25 a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz, L.; Del Río, J.A.; Ortuño, A. Mechanism of the Alternaria alternata Pathogenicity in ‘Fortune’ Mandarin. Horticulturae 2018, 4, 54. https://doi.org/10.3390/horticulturae4040054
Díaz L, Del Río JA, Ortuño A. Mechanism of the Alternaria alternata Pathogenicity in ‘Fortune’ Mandarin. Horticulturae. 2018; 4(4):54. https://doi.org/10.3390/horticulturae4040054
Chicago/Turabian StyleDíaz, Licinio, José Antonio Del Río, and Ana Ortuño. 2018. "Mechanism of the Alternaria alternata Pathogenicity in ‘Fortune’ Mandarin" Horticulturae 4, no. 4: 54. https://doi.org/10.3390/horticulturae4040054
APA StyleDíaz, L., Del Río, J. A., & Ortuño, A. (2018). Mechanism of the Alternaria alternata Pathogenicity in ‘Fortune’ Mandarin. Horticulturae, 4(4), 54. https://doi.org/10.3390/horticulturae4040054