

Optimizing Carbon Dioxide Enrichment to Balance Yield, Functional Food Quality, and Economic Feasibility in Plant-Factory-Cultivated Kale

 , , ,
, , ,  , ,
, ,  and
and 
Abstract
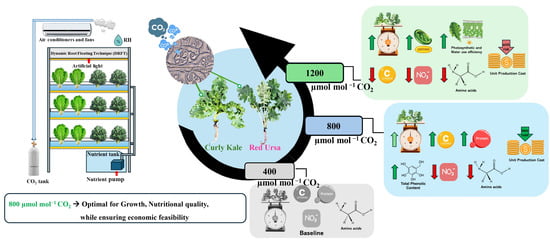
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Growth and Yield Measurements
2.3. Physiological Responses
2.4. Photosynthetic Pigment Content Evaluation
2.5. Soluble Protein, Phytonutrients, and Nitrate Analysis
2.6. Amino Profile Evaluation
2.7. Economic Evaluation
2.8. Statistical Analysis
3. Results
3.1. Effect of CO2 Concentrations on Growth and Yield of Kale
3.2. Effect of CO2 Concentrations on the Yield of Kale Plants
3.3. Effect of CO2 Concentrations on Photosynthetic Efficiency of Kale Plants
3.4. Effect of CO2 Concentrations on Spectral Indices of Kale Plants
3.5. Effect of CO2 Concentrations on Photosynthetic Pigment Accumulation
3.6. Effect of CO2 Concentration on Soluble Protein, Phytonutrients, and Nitrate Content
3.7. Effect of CO2 Concentration on the Amino Profile of Kale Plants
3.8. Heat Map Analysis
3.9. Economic Analysis
4. Discussion
4.1. Elevated CO2 Concentrations Enhance the Growth and Yield of Kale Plants
4.2. Elevated CO2 Concentrations Enhance Carbon Assimilation and Pigment Accumulation
4.3. Elevated CO2 Concentrations Alter Some Spectral Indices
4.4. Elevated CO2 Concentrations Have Both Positive and Negative Effects on Nutritional Values
4.5. Elevated CO2 Concentrations Dramatically Reduced Amino Acids in Kales
4.6. Economic Feasibility and the Nutritional Integrity of Kale as a Superfood
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cheng, F.; Wu, J.; Cai, C.; Fu, L.; Liang, J.; Borm, T.; Zhuang, M.; Zhang, Y.; Zhang, F.; Bonnema, G.; et al. Genome resequencing and comparative variome analysis in a Brassica rapa and Brassica oleracea collection. Sci. Data 2016, 3, 160119. [Google Scholar] [CrossRef] [PubMed]
- Šamec, D.; Urlić, B.; Salopek-Sondi, B. Kale (Brassica oleracea var. acephala) as a superfood: Review of the scientific evidence behind the statement. Crit. Rev. Food Sci. Nutr. 2019, 59, 2411–2422. [Google Scholar] [CrossRef]
- He, R.; Zhang, Y.; Song, S.; Su, W.; Hao, Y.; Liu, H. UV-A and FR irradiation improves growth and nutritional properties of lettuce grown in an artificial light plant factory. Food Chem. 2021, 345, 128727. [Google Scholar] [CrossRef] [PubMed]
- Khalid, W.; Iqra; Afzal, F.; Rahim, M.A.; Abdul Rehman, A.; Faiz ul Rasul, H.; Arshad, M.S.; Ambreen, S.; Zubair, M.; Safdar, S.; et al. Industrial applications of kale (Brassica oleracea var. sabellica) as a functional ingredient: A review. Int. J. Food Prop. 2023, 26, 489–501. [Google Scholar] [CrossRef]
- Ayaz, F.A.; Glew, R.H.; Millson, M.; Huang, H.S.; Chuang, L.T.; Sanz, C.; Hayirlioglu-Ayaz, S. Nutrient contents of kale (Brassica oleraceae L. var. acephala DC.). Food Chem. 2006, 96, 572–579. [Google Scholar] [CrossRef]
- National Institutes of Health Office of Dietary Supplements. Dietary Supplement Label Database. 2026. Available online: https://dsld.od.nih.gov/.
- Łukaszyk, A.; Kwiecień, I.; Kanik, A.; Blicharska, E.; Tatarczak-Michalewska, M.; Białowąs, W.; Czarnek, K.; Szopa, A. Nutritional, Therapeutic, and Functional Food Perspectives of Kale (Brassica oleracea var. acephala): An Integrative Review. Molecules 2025, 30, 4214. [Google Scholar] [CrossRef]
- Kozai, T. Smart Plant Factory: The Next Generation Indoor Vertical Farms; Springer: Singapore, 2018. [Google Scholar]
- Mandriota, L.; Blanco, I.; Scarascia-Mugnozza, G. Plant Factory with Artificial Lighting: Innovation Technology for Sustainable Agriculture Production. In Biosystems Engineering Towards the Green Deal; Springer: Cham, Switzerland, 2023; Volume 337, pp. 1163–1172. [Google Scholar]
- Terlaje, T.; Tuquero, J. Growing Kale Brassica oleracea. Nativ. Plants Guam 2018, FS016-19, 1–5. [Google Scholar]
- Liu, K.; Gao, M.; Jiang, H.; Ou, S.; Li, X.; He, R.; Li, Y.; Liu, H. Light Intensity and Photoperiod Affect Growth and Nutritional Quality of Brassica Microgreens. Molecules 2022, 27, 883. [Google Scholar] [CrossRef]
- Huang, L.C. Consumer attitude, concerns, and brand acceptance for the vegetables cultivated with sustainable plant factory production systems. Sustainability 2019, 11, 4862. [Google Scholar] [CrossRef]
- Myers, S.S.; Zanobetti, A.; Kloog, I.; Huybers, P.; Leakey, A.D.B.; Bloom, A.J.; Carlisle, E.; Dietterich, L.H.; Fitzgerald, G.; Hasegawa, T.; et al. Rising CO2 threatens human nutrition. Nature 2014, 510, 139–142. [Google Scholar] [CrossRef]
- Chen, D.; Mei, Y.; Liu, Q.; Wu, Y.; Yang, Z. Carbon dioxide enrichment promoted the growth, yield, and light-use efficiency of lettuce in a plant factory with artificial lighting. Agron. J. 2021, 113, 5196–5206. [Google Scholar] [CrossRef]
- Lupitu, A.; Moisa, C.; Gavrila, S.; Copolovici, D.M.; Copolovici, L.; Dochia, M.; Chambre, D.; Ciutin, V. The Influence of Elevated CO2 on Volatile Emissions, Photosynthetic Characteristics, and Pigment Content in Brassicaceae Plants Species and Varieties. Plants 2022, 11, 973. [Google Scholar] [CrossRef] [PubMed]
- Dou, H.; Niu, G.; Gu, M.; Masabni, J.G. Effects of light quality on growth and phytonutrient accumulation of herbs under controlled environments. Horticulturae 2017, 3, 36. [Google Scholar] [CrossRef]
- Tomimatsu, H.; Tang, Y. Effects of high CO2 levels on dynamic photosynthesis: Carbon gain, mechanisms, and environmental interactions. J. Plant Res. 2016, 129, 365–377. [Google Scholar] [CrossRef]
- Mosaleeyanon, K. Current Situation, Direction, Policy Support, and Challenges of Plant Factories with Artificial lighting (PFAL) in Thailand. FFTC J. Agric. Policy 2022, 3, 46–56. [Google Scholar] [CrossRef]
- Kozai, T.; Niu, G. Plant Factory as a Resource-Efficient Closed Plant Production System; Elsevier Inc.: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Royal Thai Government Gazette. 2022. Available online: https://www.ratchakitcha.soc.go.th/DATA/PDF/2564/E/031/T_0024.PDF (accessed on 21 March 2026). (In Thai)
- Renna, M.; Stellacci, A.M.; Corbo, F.; Santamaria, P. The use of a nutrient quality score is effective to assess the overall nutritional value of three brassica microgreens. Foods 2020, 9, 1226. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition, and Allergies (NDA). Scientific Opinion on Dietary Reference Values for vitamin A. EFSA J. 2015, 13, 4028. [CrossRef]
- European Commission. Regulation Statement on Possible Public Health Risks for Infants and Young Children from the Presence of Nitrates in Leafy Vegetables; European Commission: Brussels, Belgium, 2011. [Google Scholar]
- Chowdhury, M.; Kiraga, S.; Islam, M.N.; Ali, M.; Reza, M.N.; Lee, W.H.; Chung, S.O. Effects of temperature, relative humidity, and carbon dioxide concentration on growth and glucosinolate content of kale grown in a plant factory. Foods 2021, 10, 1524. [Google Scholar] [CrossRef]
- Hori, H. Gravel Culture of Vegetable and Ornamental Crops; OCLC No. 673083780; Youkendo Co., Ltd.: Tokyo, Japan, 1966; pp. 60–80. [Google Scholar]
- Chrysargyris, A.; Petropoulos, S.A.; Fernandes, Â.; Barros, L.; Tzortzakis, N.; Ferreira, I.C.F.R. Effect of phosphorus application rate on Mentha spicata L. grown in deep flow technique (DFT). Food Chem. 2019, 276, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Chrysargyris, A.; Nikolaidou, E.; Stamatakis, A.; Tzortzakis, N. Vegetative, physiological, nutritional and antioxidant behavior of spearmint (Mentha spicata L.) in response to different nitrogen supply in hydroponics. J. Appl. Res. Med. Aromat. Plants 2017, 6, 52–61. [Google Scholar] [CrossRef]
- Sonjaroon, W.; Tepkaew, T.; Kupia, M.; Tongkok, P.; Boonkorkaew, P.; Thussagunpanit, J. Pre-Harvest UV-A Supplementation in Plant Factory with Artificial Lighting Improves Growth, Photosynthesis, and Phytonutrients in Kale. Horticulturae 2024, 10, 701. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll Fluorescence—A Practical Guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. BBA—Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ağlar, E.; Saraçoğlu, O. Role of the foliar fertilization treatments on quality attributes of sweet cherry fruits (Prunus avium). Akad. Ziraat Derg. 2018, 7, 131–136. [Google Scholar] [CrossRef]
- Parks, S.E.; Irving, D.E.; Milham, P.J. A critical evaluation of on-farm rapid tests for measuring nitrate in leafy vegetables. Sci. Hortic. 2012, 134, 1–6. [Google Scholar] [CrossRef]
- Tepe, B.; Sokmen, A. Screening of the antioxidative properties and total phenolic contents of three endemic Tanacetum subspecies from Turkish flora. Bioresour. Technol. 2007, 98, 3076–3079. [Google Scholar] [CrossRef] [PubMed]
- Chrysargyris, A.; Papakyriakou, E.; Petropoulos, S.A.; Tzortzakis, N. The combined and single effect of salinity and copper stress on growth and quality of Mentha spicata plants. J. Hazard. Mater. 2019, 368, 584–593. [Google Scholar] [CrossRef]
- Dong, J.; Gruda, N.; Lam, S.K.; Li, X.; Duan, Z. Effects of elevated CO2 on nutritional quality of vegetables: A review. Front. Plant Sci. 2018, 9, 924. [Google Scholar] [CrossRef]
- Hou, L.; Shang, M.; Chen, Y.; Zhang, J.; Xu, X.; Song, H.; Zheng, S.; Li, M.; Xing, G. Physiological and molecular mechanisms of elevated CO2 in promoting the growth of pak choi (Brassica rapa ssp. chinensis). Sci. Hortic. 2021, 288, 110318. [Google Scholar] [CrossRef]
- Wheeler, R.M.; Spencer, L.S.E.; Bhuiyan, R.H.; Mickens, M.A.; Bunchek, J.M.; van Santen, E.; Massa, G.D.; Romeyn, M.W. Effects of elevated and super-elevated carbon dioxide on salad crops for space. J. Plant Interact. 2024, 19, 2292219. [Google Scholar] [CrossRef]
- Ferrio, J.P.; Pou, A.; Florez-Sarasa, I.; Gessler, A.; Kodama, N.; Flexas, J.; Ribas-Carbó, M. The Péclet effect on leaf water enrichment correlates with leaf hydraulic conductance and mesophyll conductance for CO2. Plant Cell Environ. 2012, 35, 611–625. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, F.; Hao, L.; Yu, J.; Guo, L.; Zhou, H.; Ma, C.; Zhang, X.; Xu, M. Elevated CO2 concentration induces photosynthetic down-regulation with changes in leaf structure, non-structural carbohydrates and nitrogen content of soybean. BMC Plant Biol. 2019, 19, 255. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Sinha, P.G.; Bhatnagar, A.K. Effect of elevated [CO2] on cell structure and function in seed plants. Clim. Chang. Environ. Sustain. 2014, 2, 69. [Google Scholar] [CrossRef]
- Dwivedi, N. The interactive effect of the elevated CO2 and moisture stress on the photosynthesis in Brassica leaves at different canopy positions. Agronomy 2012, 1, 273–276. [Google Scholar]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.; von Caemmerer, S.; Berry, J.A. A Biochemical Model of Photosynthetic CO2 Assimilation in Leaves of C3 Species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef]
- Xu, Z.; Jiang, Y.; Jia, B.; Zhou, G. Elevated-CO2 response of stomata and its dependence on environmental factors. Front. Plant Sci. 2016, 7, 657. [Google Scholar] [CrossRef]
- Franks, P.J.; Beerling, D.J. Maximum leaf conductance driven by CO2 effects on stomatal size and density over geologic time. Proc. Natl. Acad. Sci. USA 2009, 106, 10343–10347. [Google Scholar] [CrossRef]
- Mishra, R.S.; Abrol, Y.P.; Uprety, D.C. Interactive Effects of Elevated CO2 and Moisture Stress on the Photosynthesis, Water Relation and Growth of Brassica Species. J. Agron. Crop Sci. 1999, 182, 223–229. [Google Scholar] [CrossRef]
- Wang, M.; Xie, B.; Fu, Y.; Dong, C.; Hui, L.; Guanghui, L.; Liu, H. Effects of different elevated CO2 concentrations on chlorophyll contents, gas exchange, water use efficiency, and PSII activity on C3 and C4 cereal crops in a closed artificial ecosystem. Photosynth. Res. 2015, 126, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Lamichaney, A.; Tewari, K.; Basu, P.S.; Katiyar, P.K.; Singh, N.P. Effect of elevated carbon-dioxide on plant growth, physiology, yield and seed quality of chickpea (Cicer arietinum L.) in Indo-Gangetic plains. Physiol. Mol. Biol. Plants 2021, 27, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Yu, X.; Wen, J.; Jensen, N.B.; Dos Santos, T.M.; Wu, Z.; Rosenqvist, E.; Ottosen, C.O. Interactive effects of elevated CO2 concentration and combined heat and drought stress on tomato photosynthesis. BMC Plant Biol. 2020, 20, 260. [Google Scholar] [CrossRef] [PubMed]
- Greer, D.H. Leaf temperature and CO2 effects on photosynthetic CO2 assimilation and chlorophyll a fluorescence light responses during mid-ripening of Vitis vinifera cv. Shiraz grapevines grown in outdoor conditions. Funct. Plant Biol. 2022, 49, 659–671. [Google Scholar] [CrossRef]
- Gjindali, A.; Johnson, G.N. Photosynthetic acclimation to changing environments. Biochem. Soc. Trans. 2023, 51, 473–486. [Google Scholar] [CrossRef]
- Agüera, E.; De la Haba, P. Leaf senescence in response to elevated atmospheric CO2 concentration and low nitrogen supply. Biol. Plant. 2018, 62, 401–408. [Google Scholar] [CrossRef]
- Swapnil, P.; Meena, M.; Singh, S.K.; Dhuldhaj, U.P.; Harish; Marwal, A. Vital roles of carotenoids in plants and humans to deteriorate stress with its structure, biosynthesis, metabolic engineering and functional aspects. Curr. Plant Biol. 2021, 26, 100203. [Google Scholar] [CrossRef]
- Kunyawut, C.; Paopo, I.; Umpuch, C. Enhanced carotenoid accumulation in Chloroccocum humicola under controlled CO2 and light conditions. J. Genet. Eng. Biotechnol. 2025, 23, 100619. [Google Scholar] [CrossRef]
- Katsoulas, N.; Elvanidi, A.; Bartzanas, T.; Ferentinos, K.P.; Kittas, C. Sensing crop reflectance for water stress detection in greenhouses. Acta Hortic. 2018, 1197, 117–126. [Google Scholar] [CrossRef]
- Jiang, H.; Liu, Z.; Wang, J.; Yang, P.; Zhang, R.; Zhang, X.; Zheng, P. Combining Chlorophyll Fluorescence and Vegetation Reflectance Indices to Estimate Non-Photochemical Quenching (NPQ) of Rice at the Leaf Scale. Remote Sens. 2023, 15, 4222. [Google Scholar] [CrossRef]
- Gamon, J.A.; Peñuelas, J.; Field, C.B. A narrow-waveband spectral index that tracks diurnal changes in photosynthetic efficiency. Remote Sens. Environ. 1992, 41, 35–44. [Google Scholar] [CrossRef]
- Jiao, Q.; Liu, L.; Liu, J.; Zhang, H.; Zhang, B. Atmospherically resistant vegetation water indices using the 970-nm water absorption feature. J. Appl. Remote Sens. 2020, 14, 034504. [Google Scholar] [CrossRef]
- Muthusamy, M.; Hwang, J.E.; Kim, S.H.; Kim, J.A.; Jeong, M.J.; Park, H.C.; Lee, S.I. Elevated carbon dioxide significantly improves ascorbic acid content, antioxidative properties and restricted biomass production in cruciferous vegetable seedlings. Plant Biotechnol. Rep. 2019, 13, 293–304. [Google Scholar] [CrossRef]
- Mamatha, H.; Srinivasa Rao, N.K.; Laxman, R.H.; Shivashankara, K.S.; Bhatt, R.M.; Pavithra, K.C. Impact of elevated CO2 on growth, physiology, yield, and quality of tomato (Lycopersicon esculentum Mill) cv. Arka Ashish. Photosynthetica 2014, 52, 519–528. [Google Scholar] [CrossRef]
- Smirnoff, N.; Wheeler, G.L. The ascorbate biosynthesis pathway in plants is known, but there is a way to go with understanding control and functions. J. Exp. Bot. 2024, 75, 2604–2630. [Google Scholar] [CrossRef]
- Singh, A.; Agrawal, M. Effects of ambient and elevated CO2 on growth, chlorophyll fluorescence, photosynthetic pigments, antioxidants, and secondary metabolites of Catharanthus roseus (L.) G Don. grown under three different soil N levels. Environ. Sci. Pollut. Res. 2015, 22, 3936–3946. [Google Scholar] [CrossRef]
- Salesse-Smith, C.E.; Wang, Y.; Long, S.P. Increasing Rubisco as a simple means to enhance photosynthesis and productivity now without lowering nitrogen use efficiency. New Phytol. 2025, 245, 951–965. [Google Scholar] [CrossRef]
- Ekele, J.U.; Obaje, J.O.; Zajitschek, S.R.K.; Webster, R.J.; Perez de Heredia, F.; Lane, K.E.; Fadel, A.; Symonds, R.C. Effects of Elevated CO2 on Yield and Nutritional Quality of Kale and Spinach: A Meta-Analysis. Biology 2026, 15, 152. [Google Scholar] [CrossRef]
- Goufo, P.; Pereira, J.; Moutinho-Pereira, J.; Correia, C.M.; Figueiredo, N.; Carranca, C.; Rosa, E.A.S.; Trindade, H. Rice (Oryza sativa L.) phenolic compounds under elevated carbon dioxide (CO2) concentration. Environ. Exp. Bot. 2014, 99, 28–37. [Google Scholar] [CrossRef]
- Takatani, N.; Ito, T.; Kiba, T.; Mori, M.; Miyamoto, T.; Maeda, S.I.; Omata, T. Effects of high CO2 on growth and metabolism of Arabidopsis seedlings during growth with a constantly limited supply of nitrogen. Plant Cell Physiol. 2014, 55, 281–292. [Google Scholar] [CrossRef]
- Bian, Z.; Wang, Y.; Zhang, X.; Li, T.; Grundy, S.; Yang, Q.; Cheng, R. A Review of Environment Effects on Nitrate Accumulation in Leafy Vegetables Grown in Controlled Environments. Foods 2020, 9, 732. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Tzortzakis, N. Optimising fertigation of hydroponically grown sowthistle (Sonchus oleraceus L.): The impact of the nitrogen source and supply concentration. Agric. Water Manag. 2023, 289, 108528. [Google Scholar] [CrossRef]
- Pal, M.; Rao, L.S.; Jain, V.; Srivastava, A.C.; Pandey, R.; Raj, A.; Singh, K.P. Effects of elevated CO2 and nitrogen on wheat growth and photosynthesis. Biol. Plant. 2005, 49, 467–470. [Google Scholar] [CrossRef]
- Du, S.; Zhang, R.; Zhang, P.; Liu, H.; Yan, M.; Chen, N.; Xie, H.; Ke, S. Elevated CO2-induced production of nitric oxide (NO) by NO synthase differentially affects nitrate reductase activity in Arabidopsis plants under different nitrate supplies. J. Exp. Bot. 2016, 67, 893–904. [Google Scholar] [CrossRef] [PubMed]
- Rosa-Téllez, S.; Alcántara-Enguídanos, A.; Martínez-Seidel, F.; Casatejada-Anchel, R.; Saeheng, S.; Bailes, C.L.; Erban, A.; Barbosa-Medeiros, D.; Alepúz, P.; Matus, J.T.; et al. The serine–glycine–one-carbon metabolic network orchestrates changes in nitrogen and sulfur metabolism and shapes plant development. Plant Cell 2024, 36, 404–426. [Google Scholar] [CrossRef]
- Zhang, Z.; Wu, Z. CO2 enhances low-nitrogen adaption by promoting amino acid metabolism in Brassica napus. Plant Physiol. Biochem. 2023, 201, 107864. [Google Scholar] [CrossRef] [PubMed]
- Stepansky, A.; Leustek, T. Histidine biosynthesis in plants. Amino Acids 2006, 30, 127–142. [Google Scholar] [CrossRef]
- Liao, H.S.; Chung, Y.H.; Hsieh, M.H. Glutamate: A multifunctional amino acid in plants. Plant Sci. 2022, 318, 111238. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | CO2 (µmol mol−1) | Shoot Fresh Weight (g) | Root Fresh Weight (g) | Shoot Dry Weight (g) | Root Dry Weight (g) | Fresh:Dry Weight Ratio | Stem Diameter (mm) | Leaf Greenness Index (SPAD Unit) |
|---|---|---|---|---|---|---|---|---|
| 400 | 67.25 ± 8.56 b | 15.21 ± 4.68 b | 4.70 ± 0.59 c | 0.87 ± 0.01 b | 14.75 ± 0.86 a | 7.75 ± 0.77 b | 42.55 ± 1.80 a | |
| Curly Kale | 800 | 111.00 ± 12.43 b | 11.12 ± 1.75 b | 8.08 ± 0.75 b | 0.55 ± 0.08 c | 14.15 ± 0.97 a | 15.09 ± 1.01 a | 44.73 ± 8.18 a |
| 1200 | 176.87 ± 49.07 a | 22.05 ± 0.12 a | 15.19 ± 1.43 a | 1.10 ± 0.00 a | 12.23 ± 2.89 a | 8.00 ± 2.42 b | 41.45 ± 8.01 a | |
| F-test | * | *** | *** | *** | ns | *** | ns | |
| Red Ursa | 400 | 111.28 ± 19.67 a | 10.95 ± 3.10 b | 7.78 ± 1.37 a | 0.54 ± 0.15 b | 14.65 ± 0.05 b | 8.25 ± 0.89 b | 46.17 ± 1.50 a |
| 800 | 113.25 ± 7.41 a | 11.00 ± 0.81 b | 7.92 ± 0.51 a | 0.55 ± 0.04 b | 14.65 ± 0.01 b | 15.81 ± 3.97 a | 40.22 ± 3.25 a | |
| 1200 | 113.75 ± 31.71 a | 16.57 ± 3.73 a | 7.96 ± 2.07 a | 0.82 ± 0.18 a | 14.83 ± 0.06 a | 14.5 ± 1.42 a | 33.32 ± 5.77 b | |
| F-test | ns | * | ns | * | *** | * | * |
| Cultivar | CO2 (µmol mol−1) | Total Chl (mg g−1 FW) | Chl a (mg g−1 FW) | Chl b (mg g−1 FW) | Chl a/Chl b Ratio | Total Car (mg g−1 FW) | Total Car/Total Chl Ratio |
|---|---|---|---|---|---|---|---|
| Curly Kale | 400 | 8.24 ± 0.83 b | 5.82 ± 0.54 b | 2.42 ± 0.31 a | 2.42 ± 0.12 b | 0.66 ± 0.14 b | 0.08 ± 0.17 b |
| 800 | 9.17 ± 0.72 b | 7.11 ± 1.66 ab | 2.81 ± 0.53 a | 2.52 ± 0.15 ab | 1.35 ± 0.23 a | 0.15 ± 0.32 a | |
| 1200 | 11.53 ± 1.52 a | 8.46 ± 1.19 a | 3.07 ± 0.35 a | 2.75 ± 0.12 a | 1.49 ± 0.13 a | 0.13 ± 0.09 a | |
| F-test | ** | * | ns | * | *** | ** | |
| Red Ursa | 400 | 8.38 ± 0.69 b | 6.15 ± 0.46 a | 2.23 ± 0.23 b | 2.77 ± 0.09 a | 0.98 ± 0.11 b | 0.12 ± 0.16 b |
| 800 | 9.98 ± 0.99 b | 8.63 ± 1.19 a | 3.39 ± 0.46 a | 2.55 ± 0.02 b | 1.83 ± 0.25 a | 0.18 ± 0.25 a | |
| 1200 | 11.00 ± 0.73 a | 7.00 ± 2.02 a | 2.64 ± 0.76 ab | 2.67 ± 0.15 ab | 2.53 ± 0.51 a | 0.23 ± 0.70 a | |
| F-test | ** | ns | * | * | *** | ** |
| Amino Acid (mg/100 g DW) | Curly Kale | Red Ursa | ||||||
|---|---|---|---|---|---|---|---|---|
| 400 µmol mol−1 | 800 µmol mol−1 | 1200 µmol mol−1 | F-Test | 400 µmol mol−1 | 800 µmol mol−1 | 1200 µmol mol−1 | F-Test | |
| Aspartic acid | 2807 ± 40.82 a | 2148 ± 54.31 b | 2108 ± 55.43 b | *** | 2540 ± 38.38 a | 2436 ± 40.82 b | 1924 ± 32.66 c | *** |
| Glutamic acid | 3721.33 ± 30.64 a | 3042 ± 52.26 b | 2964 ± 40.82 b | *** | 3395 ± 48.99 b | 3694 ± 57.15 a | 2853 ± 53.89 c | *** |
| Serine | 1274 ± 124.16 a | 878.77 ± 53.49 b | 837.04 ± 57.81 b | *** | 1004 ± 41.64 a | 1032 ± 33.48 a | 733.05 ± 41.64 b | *** |
| Histidine | 566.75 ± 14.70 a | 538.29 ± 40.56 a | 462.15 ± 23.14 b | ** | 560.77 ± 28.58 a | 532.82 ± 14.70 a | 471.38 ± 13.88 b | *** |
| Glycine | 1513 ± 28.58 a | 1433.16 ± 41.31 a | 1186.75 ± 132.24 b | ** | 1469 ± 40.01 a | 1364 ± 124.16 ab | 1227 ± 28.58 b | ** |
| Threonine | 1123 ± 33.48 a | 742.605 ± 66.13 b | 680.56 ± 74.73 b | *** | 938.72 ± 14.70 a | 914.81 ± 28.58 a | 590.18 ± 13.80 b | *** |
| Arginine | 1536 ± 40.01 b | 1709 ± 42.25 a | 1486 ± 28.58 b | *** | 1565 ± 124.16 b | 2073 ± 41.64 a | 1570 ± 33.48 b | *** |
| Alanine | 1621 ± 41.64 a | 1335.50 ± 31.40 b | 1322.50 ± 35.91 b | *** | 1607 ± 40.82 a | 1594 ± 40.01 a | 1468 ± 124.16 a | ns |
| Tyrosine | 591.77 ± 12.25 a | 277.68 ± 18.14 b | 264.31 ± 19.28 b | *** | 386.15 ± 14.70 a | 295.25 ± 13.94 b | 177.95 ± 22.86 c | *** |
| Cystine | nd | nd | nd | - | nd | nd | nd | - |
| Valine | 1451 ± 28.58 a | 1210.75 ± 27.55 b | 1186.25 ± 48.31 b | *** | 1412 ± 33.48 a | 1344 ± 30.12 b | 1256 ± 8.98 c | *** |
| Methionine | 283.06 ± 12.25 a | 142.93 ± 14.70 b | 128.28 ± 13.88 b | *** | 183.76 ± 12.75 a | 134.44 ± 22.86 b | 121.49 ± 14.70 b | ** |
| Phenylalanine | 1447 ± 24.04 a | 1185.75 ± 49.36 b | 1159.25 ± 42.70 b | *** | 1521 ± 124.16 a | 1382 ± 115.95 a | 1300 ± 120.64 a | ns |
| Isoleucine | 1047 ± 13.88 a | 846.30 ± 46.46 b | 814.08 ± 35.41 b | *** | 1013 ± 28.58 a | 931.65 ± 13.88 b | 892.51 ± 28.58 b | *** |
| Leucine | 2295 ± 40.01 a | 1733 ± 91.24 b | 1651 ± 96.37 b | *** | 2217 ± 33.48 a | 1875 ± 28.58 b | 1821 ± 40.01 b | *** |
| Lysine | 1790 ± 33.48 a | 1551.25 ± 129.70 b | 1438.75 ± 47.15 b | ** | 1734 ± 41.64 a | 1653 ± 40.36 b | 1498 ± 33.48 c | *** |
| Tryptophan | 141.74 ± 14.70 a | 168.16 ± 13.88 a | 144.27 ± 22.86 a | ns | 135.27 ± 12.25 a | 143.46 ± 10.44 a | 144.25 ± 13.80 a | ns |
| Proline | 1287 ± 221.86 a | 1103.65 ± 31.73 b | 1060.56 ± 139.82 b | ** | 1263 ± 40.01 a | 1120.50 ± 54.49 b | 1083.50 ± 43.49 b | ** |
| Total essential amino acid (EA) | 10,145 ± 192.57 a | 8119 ± 414.79 b | 7665 ± 385.46 b | *** | 9716 ± 323.22 a | 8911 ± 312.76 b | 8095 ± 285.44 c | *** |
| Total non-essential amino acid (NEA) | 14,351 ± 327.15 a | 11,928 ± 219.08 b | 11,229 ± 484.62 b | *** | 13,229 ± 380.53 a | 13,609 ± 385.54 a | 11,037 ± 354.19 b | *** |
| Total amino acid (TA) | 24,496 ± 516.21 a | 20,047 ± 632.47 b | 18,894 ± 852.41 b | *** | 22,945 ± 703.69 a | 22,520 ± 697.41 a | 19,131 ± 638.66 b | *** |
| EA/TA ratio | 0.414 ± 0.002 a | 0.405 ± 0.008 a | 0.406 ± 0.005 a | ns | 0.423 ± 0.001 a | 0.396 ± 0.004 a | 0.423 ± 0.005 a | ns |
| NEA/TA ratio | 0.586 ± 0.002 a | 0.595 ± 0.008 a | 0.594 ± 0.005 a | ns | 0.576 ± 0.001 b | 0.604 ± 0.001 a | 0.577 ± 0.001 b | *** |
| Description | Curly Kale | Red Ursa | ||||
|---|---|---|---|---|---|---|
| 400 µmol mol−1 | 800 µmol mol−1 | 1200 µmol mol−1 | 400 µmol mol−1 | 800 µmol mol−1 | 1200 µmol mol−1 | |
| Seeds (960 seeds) | 9.30 | 9.30 | 9.30 | 46.50 | 46.50 | 46.50 |
| Growing media (960 sets) | 29.76 | 29.76 | 29.76 | 29.76 | 29.76 | 29.76 |
| Nutrient solution | 89.28 | 89.28 | 89.28 | 89.28 | 89.28 | 89.28 |
| Electricity charges | 496.00 | 496.00 | 496.00 | 496.00 | 496.00 | 496.00 |
| Water charges | 62.00 | 62.00 | 62.00 | 62.00 | 62.00 | 62.00 |
| CO2 | 77.50 | 155.00 | 387.50 | 77.50 | 155.00 | 387.50 |
| Planting and maintenance labor | 930.00 | 930.00 | 930.00 | 930.00 | 930.00 | 930.00 |
| Depreciation and maintenance | 124.00 | 124.00 | 124.00 | 124.00 | 124.00 | 124.00 |
| Total Cost per planting cycle (USD) | 1817.84 | 1895.34 | 2127.84 | 1855.04 | 1932.54 | 2165.04 |
| Production cost per plant (USD plant−1) | 1.89 | 1.97 | 2.22 | 1.93 | 2.01 | 2.26 |
| Shoot fresh weight (g plant−1) | 67.25 | 111.00 | 176.87 | 111.28 | 113.25 | 113.75 |
| Unit production cost (USD kg−1) | 28.16 | 17.79 | 12.53 | 17.36 | 17.78 | 19.83 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Kupia, M.; Sonjaroon, W.; Matmarurat, G.; Shigyo, M.; Boonkorkaew, P.; Tzortzakis, N.; Thussagunpanit, J. Optimizing Carbon Dioxide Enrichment to Balance Yield, Functional Food Quality, and Economic Feasibility in Plant-Factory-Cultivated Kale. Horticulturae 2026, 12, 621. https://doi.org/10.3390/horticulturae12050621
Kupia M, Sonjaroon W, Matmarurat G, Shigyo M, Boonkorkaew P, Tzortzakis N, Thussagunpanit J. Optimizing Carbon Dioxide Enrichment to Balance Yield, Functional Food Quality, and Economic Feasibility in Plant-Factory-Cultivated Kale. Horticulturae. 2026; 12(5):621. https://doi.org/10.3390/horticulturae12050621
Chicago/Turabian StyleKupia, Manop, Weerasin Sonjaroon, Gadewara Matmarurat, Masayoshi Shigyo, Patchareeya Boonkorkaew, Nikolaos Tzortzakis, and Jutiporn Thussagunpanit. 2026. "Optimizing Carbon Dioxide Enrichment to Balance Yield, Functional Food Quality, and Economic Feasibility in Plant-Factory-Cultivated Kale" Horticulturae 12, no. 5: 621. https://doi.org/10.3390/horticulturae12050621
APA StyleKupia, M., Sonjaroon, W., Matmarurat, G., Shigyo, M., Boonkorkaew, P., Tzortzakis, N., & Thussagunpanit, J. (2026). Optimizing Carbon Dioxide Enrichment to Balance Yield, Functional Food Quality, and Economic Feasibility in Plant-Factory-Cultivated Kale. Horticulturae, 12(5), 621. https://doi.org/10.3390/horticulturae12050621