
Foliar Treatments with Urea and Nano-Urea Modify the Nitrogen Profile of Monastrell Grapes and Wines

 ,
,  ,
,  , ,
, ,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents and Standards
2.2. Plant Material, Experimental Design in Field and Treatments
2.3. Vinifications
2.4. Physicochemical Parameters of Grapes and Wines
2.5. Analysis of Nitrogen Compounds in Grapes by HPLC
2.6. Analysis of Nitrogen Compounds in Wines by HPLC
2.7. Statistical Analysis
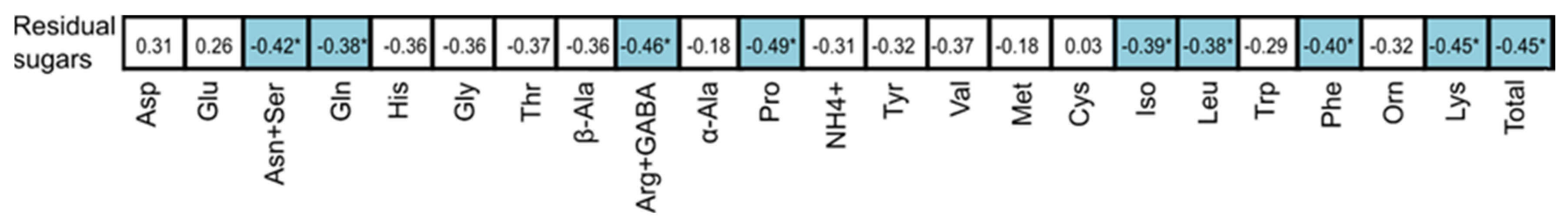
3. Results and Discussion
3.1. Physicochemical Parameters of Grapes and Wines
3.2. Nitrogen Composition of Grapes
3.3. Nitrogen Composition of Wines
3.4. Multivariate Discriminant Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bellido López, L.; Ramos Mompó, C.; Betrán Aso, J.; Pomares García, F. Abonado del viñedo. In Guía Práctica de la Fertilización Racional de los Cultivos en España. Parte II; Ministerio de Medio Ambiente Y Medio Rural Y Marino: Madrid, Spain, 2010; pp. 213–219. [Google Scholar]
- Thomidis, T.; Zioziou, E.; Koundouras, S.; Karagiannidis, C.; Navrozidis, I.; Nikolaou, N. Effects of nitrogen and irrigation on the quality of grapes and the susceptibility to botrytis bunch rot. Sci. Hortic. 2016, 212, 60–68. [Google Scholar] [CrossRef]
- Guilpart, N.; Metay, A.; Gary, C. Grapevine bud fertility and number of berries per bunch are determined by water and nitrogen stress around flowering in the previous year. Eur. J. Agron. 2014, 54, 9–20. [Google Scholar] [CrossRef]
- Schreiner, R.P.; Lee, J.; Skinkis, P.A. N, P, and K supply to Pinot Noir grapevines: Impact on vine nutrient status, growth, physiology, and yield. Am. J. Enol. Vitic. 2013, 64, 26–38. [Google Scholar] [CrossRef]
- Verdenal, T.; Dienes-Nagy, Á.; Spangenberg, J.E.; Zufferey, V.; Spring, J.L.; Viret, O.; Marin-Carbonne, J.; van Leeuwen, C. Understanding and managing nitrogen nutrition in grapevine: A Review. Oeno One 2021, 55, 1–43. [Google Scholar] [CrossRef]
- Bell, S.J.; Henschke, P.A. Implications of nitrogen nutrition for grapes, fermentation and wine. Aust. J. Grape Wine Res. 2005, 11, 242–295. [Google Scholar] [CrossRef]
- Hernández-Orte, P.; Cacho, J.F.; Ferreira, V. Relationship between varietal amino acid profile of grapes and wine aromatic composition. Experiments with model solutions and chemometric study. J. Agric. Food Chem. 2002, 50, 2891–2899. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Gamboa, G.; Alañón-Sánchez, N.; Mateluna-Cuadra, R.; Verdugo-Vásquez, N. An overview about the impacts of agricultural practices on grape nitrogen composition: Current research approaches. Food Res. Int. 2020, 136, 109477. [Google Scholar] [CrossRef]
- Gil-Muñoz, R.; Giménez-Bañón, M.J.; Moreno-Olivares, J.D.; Paladines-Quezada, D.F.; Bleda-Sánchez, J.A.; Fernández-Fernández, J.I.; Parra-Torrejón, B.; Ramírez-Rodríguez, G.B.; Delgado-López, J.M. Effect of methyl jasmonate doped nanoparticles on nitrogen composition of Monastrell grapes and wines. Biomolecules 2021, 11, 1631. [Google Scholar] [CrossRef]
- Costantini, A.; Vaudano, E.; Pulcini, L.; Carafa, T.; Garcia-Moruno, E. An Overview on biogenic amines in wine. Beverages 2019, 5, 19. [Google Scholar] [CrossRef]
- Hannam, K.D.; Neilsen, G.H.; Neilsen, D.; Midwood, A.J.; Millard, P.; Zhang, Z.; Thornton, B.; Steinke, D. Amino acid composition of grape (Vitis vinifera L.) juice in response to applications of urea to the soil or foliage. Am. J. Enol. Vitic. 2016, 67, 47–55. [Google Scholar] [CrossRef]
- Lasa, B.; Menendez, S.; Sagastizabal, K.; Cervantes, M.E.C.; Irigoyen, I.; Muro, J.; Aparicio-Tejo, P.M.; Ariz, I. Foliar application of urea to “Sauvignon Blanc” and “Merlot” vines: Doses and time of application. Plant Growth Regul. 2012, 67, 73–81. [Google Scholar] [CrossRef]
- Hannam, K.D.; Neilsen, G.H.; Neilsen, D.; Rabie, W.S.; Midwood, A.J.; Millard, P. Late-season foliar urea applications can increase berry yeast-assimilable nitrogen in winegrapes (Vitis vinifera L.). Am. J. Enol. Vitic. 2014, 65, 89–95. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Diez-Zamudio, F.; Stefanello, L.O.; Tassinari, A.; Brunetto, G. Application of foliar urea to grapevines: Productivity and flavour components of grapes. Aust. J. Grape Wine Res. 2022, 28, 27–40. [Google Scholar] [CrossRef]
- Pérez-de-Luque, A.; Hermosín, M.C. Nanotechnology and its use in agriculture. In Bio-Nanotechnology; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 383–398. ISBN 9781118451915. [Google Scholar]
- Wang, X.; Xie, H.; Wang, P.; Yin, H. Nanoparticles in plants: Uptake, transport and physiological activity in leaf and root. Materials 2023, 16, 3097. [Google Scholar] [CrossRef]
- Sabir, A.; Yazar, K.; Sabir, F.; Kara, Z.; Yazici, M.A.; Goksu, N. Vine growth, yield, berry quality attributes and leaf nutrient content of grapevines as influenced by seaweed extract (Ascophyllum nodosum) and nanosize fertilizer pulverizations. Sci. Hortic. 2014, 175, 1–8. [Google Scholar] [CrossRef]
- Pérez-Álvarez, E.P.; Ramírez-Rodríguez, G.B.; Carmona, F.J.; Martínez-Vidaurre, J.M.; Masciocchi, N.; Guagliardi, A.; Garde-Cerdán, T.; Delgado-López, J.M. Towards a more sustainable viticulture: Foliar application of N-Doped calcium phosphate nanoparticles on Tempranillo grapes. J. Sci. Food Agric. 2020, 101, 1307–1313. [Google Scholar] [CrossRef]
- Ramírez-Rodríguez, G.B.; Dal Sasso, G.; Carmona, F.J.; Miguel-Rojas, C.; Pérez-De-Luque, A.; Masciocchi, N.; Guagliardi, A.; Delgado-López, J.M. Engineering biomimetic calcium phosphate nanoparticles: A green synthesis of slow-release multinutrient (NPK) nanofertilizers. ACS Appl. Bio Mater. 2020, 3, 1344–1353. [Google Scholar] [CrossRef]
- Carmona, F.J.; Dal Sasso, G.; Ramírez-Rodríguez, G.B.; Pii, Y.; Delgado-López, J.M.; Guagliardi, A.; Masciocchi, N. Urea-functionalized amorphous calcium phosphate nanofertilizers: Optimizing the synthetic strategy towards environmental sustainability and manufacturing costs. Sci. Rep. 2021, 11, 3419. [Google Scholar] [CrossRef]
- Gómez-Alonso, S.; Hermosín-Gutiérrez, I.; García-Romero, E. Simultaneous HPLC analysis of biogenic amines, amino acids, and ammonium ion as aminoenone derivatives in wine and beer samples. J. Agric. Food Chem. 2007, 55, 608–613. [Google Scholar] [CrossRef]
- Garde-Cerdán, T.; Gutiérrez-Gamboa, G.; Portu, J.; Fernández-Fernández, J.I.; Gil-Muñoz, R. Impact of phenylalanine and urea applications to Tempranillo and Monastrell vineyards on grape amino acid content during two consecutive vintages. Food Res. Int. 2017, 102, 451–457. [Google Scholar] [CrossRef]
- Portu, J.; López-Alfaro, I.; Gómez-Alonso, S.; López, R.; Garde-Cerdán, T. Changes on grape phenolic composition induced by grapevine foliar applications of phenylalanine and urea. Food Chem. 2015, 180, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Portu, J.; González-Arenzana, L.; Hermosín-Gutiérrez, I.; Santamaría, P.; Garde-Cerdán, T. Phenylalanine and urea foliar applications to grapevine: Effect on wine phenolic content. Food Chem. 2015, 180, 55–63. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Garde-Cerdán, T.; Portu, J.; Moreno-Simunovic, Y.; Martínez-Gil, A.M. Foliar nitrogen application in Cabernet Sauvignon vines: Effects on wine flavonoid and amino acid content. Food Res. Int. 2017, 96, 46–53. [Google Scholar] [CrossRef]
- Lang, C.P.; Merkt, N.; Klaiber, I.; Pfannstiel, J.; Zörb, C. Different forms of nitrogen application affect metabolite patterns in grapevine leaves and the sensory of wine. Plant Physiol. Biochem. 2019, 143, 308–319. [Google Scholar] [CrossRef]
- Kemsawasd, V.; Viana, T.; Ardö, Y.; Arneborg, N. Influence of nitrogen sources on growth and fermentation performance of different wine yeast species during alcoholic fermentation. Appl. Microbiol. Biotechnol. 2015, 99, 10191–10207. [Google Scholar] [CrossRef] [PubMed]
- Fairbairn, S.; McKinnon, A.; Musarurwa, H.T.; Ferreira, A.C.; Bauer, F.F. The impact of single amino acids on growth and volatile aroma production by Saccharomyces cerevisiae strains. Front. Microbiol. 2017, 8, 2554. [Google Scholar] [CrossRef]
- Gobert, A.; Tourdot-Maréchal, R.; Sparrow, C.; Morge, C.; Alexandre, H. Influence of nitrogen status in wine alcoholic fermentation. Food Microbiol. 2019, 83, 71–85. [Google Scholar] [CrossRef]
- Garde-Cerdán, T.; Lorenzo, C.; Lara, J.F.; Pardo, F.; Ancín-Azplicueta, C.; Salinas, M.R. Study of the evolution of nitrogen compounds during grape ripening. Application to differentiate grape varieties and cultivated systems. J. Agric. Food Chem. 2009, 57, 2410–2419. [Google Scholar] [CrossRef] [PubMed]
- Garde-Cerdán, T.; Santamaría, P.; Rubio-Bretón, P.; González-Arenzana, L.; López-Alfaro, I.; López, R. Foliar application of proline, phenylalanine, and urea to Tempranillo vines: Effect on grape volatile composition and comparison with the use of commercial nitrogen fertilizers. LWT-Food Sci. Technol. 2015, 60, 684–689. [Google Scholar] [CrossRef]
- Murillo-Peña, R.; Garde-Cerdán, T.; Martínez-Vidaurre, J.M. Evaluation of foliar applications of urea at three concentrations on grape amino acids composition. J. Sci. Food Agric. 2023, 103, 4826–4837. [Google Scholar] [CrossRef]
- Arias-Gil, M.; Garde-Cerdán, T.; Ancín-Azpilicueta, C. Influence of addition of ammonium and different amino acid concentrations on nitrogen metabolism in spontaneous must fermentation. Food Chem. 2007, 103, 1312–1318. [Google Scholar] [CrossRef]
- Takagi, H. Proline as a stress protectant in yeast: Physiological functions, metabolic regulations, and biotechnological applications. Appl. Microbiol. Biotechnol. 2008, 81, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Gamboa, G.; Portu, J.; Santamaría, P.; López, R.; Garde-Cerdán, T. Effects on grape amino acid concentration through foliar application of three different elicitors. Food Res. Int. 2017, 99, 688–692. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Time (min) * | 0 | 10 | 19 | 20 | 23 | 24 | 26.5 | 28.5 | 32.5 | 60.5 |
|---|---|---|---|---|---|---|---|---|---|---|
| Phase A (%) | 94 | 86 | 80 | 74 | 70 | 50 | 37 | 0 | 37 | 94 |
| Phase B (%) | 6 | 14 | 20 | 26 | 30 | 50 | 63 | 100 | 63 | 6 |
| Season | Control | Urea | Nano-Urea | p | ||
|---|---|---|---|---|---|---|
| Grapes | 2019 | 8.16 ± 0.84 | 8.17 ± 0.54 | 7.83 ± 0.30 | ns | |
| °Brix/TA | 2020 | 10.83 ± 0.62 | 11.07 ± 0.34 | 10.49 ± 0.84 | ns | |
| 2021 | 12.29 ± 0.93 | 12.13 ± 0.18 | 10.91 ± 1.03 | ns | ||
| Potential alcohol (% vol 20 °C) | 2019 | 13.55 ± 0.52 | 13.20 ± 0.51 | 13.43 ± 0.34 | ns | |
| 2020 | 15.07 ± 0.39 | 14.45 ± 0.66 | 14.55 ± 0.26 | ns | ||
| 2021 | 13.85 ± 0.26 b | 14.33 ± 0.39 a | 13.71 ± 0.41 b | * | ||
| Wines | 2019 | 12.94 ± 0.16 | 12.86 ± 0.07 | 13.15 ± 0.42 | ns | |
| Alcohol | 2020 | 14.72 ± 0.34 a | 13.84 ± 0.29 b | 14.18 ± 0.20 ab | * | |
| (% vol 20 °C) | 2021 | 13.44 ± 0.15 b | 13.90 ± 0.13 a | 13.32 ± 0.05 b | ** | |
| 2019 | 8.50 ± 5.80 a | 2.10 ± 0.36 b | 1.77 ± 0.06 b | * | ||
| Residual sugar | 2020 | 1.83 ± 0.21 a | 1.40 ± 0.10 b | 1.40 ± 0.00 b | * | |
| (g L−1) | 2021 | 2.43 ± 0.49 | 2.17 ± 0.06 | 1.73 ± 0.06 | ns |
| 2019 | 2020 | 2021 | Multifactorial Analysis | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | Urea | Nano-Urea | p | Control | Urea | Nano-Urea | p | Control | Urea | Nano-Urea | p | T (%) | S (%) | TxS (%) | |
| Asp | 27.6 ± 0.3 b | 19.4 ± 0.5 c | 34.6 ± 1.1 a | *** | 10.2 ± 0.5 b | 9.9 ± 0.2 b | 11.6 ± 0.7 a | * | 7.7 ± 0.1 b | 11.0 ± 0.6 a | 7.9 ± 1.0 b | ** | 4 *** | 83 *** | 12 *** |
| Glu | 33.3 ± 0.7 b | 27.6 ± 0.7 c | 44.2 ± 1.1 a | *** | 13.1 ± 0.6 b | 14.4 ± 0.2 b | 17.2 ± 1.0 a | *** | 10.7 ± 0.3 c | 20.5 ± 0.9 a | 12.5 ± 1.1 b | *** | 5 *** | 80 *** | 15 *** |
| Asn+Ser | 42.3 ± 3.5 c | 63.8 ± 2.8 b | 93.7 ± 3.8 a | *** | 91.0 ± 6.0 | 92.5 ± 2.4 | 104.4 ± 7.8 | ns | 55.1 ± 2.2 b | 94.0 ± 1.9 a | 56.8 ± 9.0 b | *** | 22 *** | 39 *** | 36 *** |
| Gln | 53.6 ± 4.9 c | 88.3 ± 4.3 b | 176.7 ± 7.3 a | *** | 198.5 ± 112.9 b | 168.3 ± 6.8 c | 239.2 ± 20.0 a | ** | 68.8 ± 3.2 b | 160.2 ± 2.8 a | 81.3 ± 112.2 b | *** | 15 *** | 54 *** | 29 *** |
| His | 13.1 ± 0.6 c | 19.8 ± 0.5 b | 43.4 ± 1.5 a | *** | 69.7 ± 6.2 b | 57.6 ± 3.2 b | 81.4 ± 8.7 a | * | 17.4 ± 1.0 b | 38.1 ± 0.9 a | 20.3 ± 3.6 b | *** | 7 *** | 77 *** | 14 *** |
| Gly | 6.0 ± 0.3 c | 6.9 ± 0.1 b | 9.5 ± 0.3 a | *** | 12.3 ± 0.6 a | 10.9 ± 0.6 b | 13.0 ± 0.8 a | * | 6.9 ± 0.1 b | 11.0 ± 0.2 a | 7.1 ± 0.6 b | *** | 7 *** | 63 *** | 28 *** |
| Thr | 14.9 ± 1.2 c | 21.7 ± 0.9 b | 34.1 ± 1.1 a | *** | 43.1 ± 3.3 | 43.0 ± 3.6 | 49.7 ± 3.7 | ns | 19.6 ± 1.2 b | 33.2 ± 0.5 a | 16.1 ± 1.7 c | *** | 7 *** | 70 *** | 20 *** |
| β-Ala | 10.5 ± 0.8 c | 15.4 ± 0.6 b | 24.3 ± 0.8 a | *** | 28.9 ± 4.3 | 26.5 ± 0.8 | 34.5 ± 4.5 | ns | 13.9 ± 0.8 b | 23.6 ± 0.3 a | 11.4 ± 1.2 c | *** | 8 *** | 60 *** | 27 *** |
| Arg+GABA | 42.5 ± 3.9 c | 93.8 ± 4.9 b | 202.6 ± 10.7 a | *** | 213.8 ± 20.2 b | 238.9 ± 11.2 b | 280.4 ± 25.2 a | * | 95.6 ± 4.5 c | 268.0 ± 4.9 a | 181.1 ± 34.7 b | *** | 31 *** | 45 *** | 21 *** |
| α-Ala | 21.5 ± 1.1 | 22.1 ± 1.8 | 22.2 ± 0.3 | ns | 23.8 ± 1.0 b | 24.6 ± 1.1 b | 27.0 ± 0.8 a | * | 31.2 ± 0.8 c | 47.9 ± 0.6 b | 53.8 ± 1.6 a | *** | 10 *** | 75 *** | 14 *** |
| Pro | 199.3 ± 8.4 c | 322.8 ± 9.6 b | 386.4 ± 6.0 a | *** | 345.9 ± 14.5 b | 357.2 ± 17.3 b | 393.6 ± 11.4 a | * | 216.2 ± 7.2 b | 360.8 ± 7.8 a | 291.3 ± 62.4 a | ** | 45 *** | 23 *** | 24 *** |
| NH4+ | 14.9 ± 0.8 c | 25.3 ± 0.8 b | 33.0 ± 0.4 a | *** | 21.6 ± 1.1 b | 24.2 ± 1.5 b | 33.2 ± 1.8 a | *** | 9.6 ± 0.3 b | 14.1 ± 0.4 a | 16.8 ± 3.2 a | ** | 40 *** | 51 *** | 7 *** |
| Tyr | 7.0 ± 0.5 c | 8.6 ± 0.2 b | 10.5 ± 0.3 a | *** | 26.1 ± 0.8 b | 25.4 ± 1.0 b | 37.3 ± 1.9 a | *** | 7.23 ± 0.4 c | 16.2 ± 0.6 a | 10.8 ± 0.8 b | *** | 6 *** | 85 *** | 9 *** |
| Val | 8.9 ± 0.4 c | 12.5 ± 0.2 b | 20.7 ± 1.3 a | *** | 33.6 ± 2.1 b | 28.4 ± 0.7 c | 39.4 ± 2.6 a | ** | 15.0 ± 0.3 b | 26.5 ± 0.7 a | 12.7 ± 0.6 c | *** | 4 *** | 72 *** | 22 *** |
| Met | 2.8 ± 0.3 b | 2.9 ± 0.2 b | 4.0 ± 0.6 a | * | 4.8 ± 0.7 b | 4.8 ± 0.4 b | 6.1 ± 0.4 a | * | 1.9 ± 0.1 | 1.9 ± 0.1 | 1.9 ± 0.12 | ns | 7 *** | 85 *** | 3 * |
| Cys | 10.4 ± 1.8 a | 6.5 ± 0.5 b | 7.1 ± 0.9 b | * | 13.2 ± 2.1 a | 8.9 ± 0.9 b | 9.2 ± 1.5 b | * | 12.3 ± 1.0 a | 7.4 ± 0.4 b | 5.8 ± 0.7 c | *** | 65 *** | 16 ** | 5 |
| Iso | 6.5 ± 0.1 c | 8.0 ± 0.0 b | 10.8 ± 0.8 a | *** | 15.1 ± 1.1 ab | 13.2 ± 0.6 b | 17.0 ± 1.1 a | ** | 12.3 ± 0.7 b | 14.0 ± 0.5 a | 8.5 ± 0.7 c | *** | 1 | 65 *** | 31 *** |
| Leu | 8.1 ± 0.4 c | 10.9 ± 0.2 b | 16.1 ± 0.7 a | *** | 24.8 ± 1.7 a | 20.6 ± 0.6 b | 27.8 ± 2.3 a | ** | 14.3 ± 0.9 b | 20.6 ± 0.6 a | 11.3 ± 0.8 c | *** | 3 *** | 70 *** | 25 *** |
| Trp | 20.3 ± 1.4 c | 23.6 ± 1.2 b | 32.2 ± 1.8 a | *** | 59.3 ± 5.8 a | 45.3 ± 2.5 b | 52.8 ± 4.3 ab | * | 29.7 ± 1.0 b | 32.6 ± 0.9 a | 18.5 ± 1.9 c | *** | 1 ns | 81 *** | 16 *** |
| Phe | 5.3 ± 0.2 c | 9.1 ± 0.3 b | 13.4 ± 1.0 a | *** | 17.3 ± 2.7 | 14.7 ± 0.6 | 17.7 ± 1.8 | ns | 9.2 ± 0.6 b | 13.8 ± 0.6 a | 6.4 ± 0.2 c | *** | 4 ** | 57 *** | 34 *** |
| Orn | 2.1 ± 0.1 c | 2.3 ± 0.1 b | 2.8 ± 0.1 a | *** | 2.6 ± 0.1 | 2.7 ± 0.1 | 2.6 ± 0.3 | ns | 2.2 ± 0.6 | 2.1 ± 0.1 | 1.8 ± 0.0 | ns | 1 | 42 *** | 29 ** |
| Lys | 2.7 ± 0.1 c | 3.2 ± 0.3 b | 3.8 ± 0.2 a | *** | 3.4 ± 0.1 | 3.9 ± 0.1 | 4.1 ± 0.8 | ns | 2.6 ± 0.2 b | 3.0 ± 0.2 ab | 3.5 ± 0.3 a | * | 45 *** | 32 *** | 3 |
| Total | 553.6 ± 19.7 c | 814.6 ± 23.9 b | 1226.0 ± 38.8 a | *** | 1271.9 ± 64.1 b | 1235.7 ± 28.8 b | 1499.1 ± 91.6 a | ** | 659.3 ± 23.1 c | 1220.3 ± 23.7 a | 837.7 ± 126.2 b | *** | 24 *** | 47 *** | 26 *** |
| Total-Pro | 354.3 ± 16.6 c | 491.8 ± 14.3 b | 839.6 ± 32.8 a | *** | 926.0 ± 60.0 b | 878.4 ± 32.3 b | 1105.6 ± 82.6 a | ** | 443.1 ± 16.1 c | 859.6 ± 15.9 a | 546.4 ± 66.5 b | *** | 18 *** | 53 *** | 26 *** |
| N 1 | 88.5 ± 3.3 c | 136.2 ± 3.4 b | 210.9 ± 6.2 a | *** | 216.0 ± 11.1 b | 211.4 ± 5.0 b | 261.6 ± 16.2 a | ** | 105.8 ± 3.7 c | 202.7 ± 3.9 a | 141.1 ± 18.4 b | *** | |||
| N-Pro | 64.2 ± 2.8 c | 96.9 ± 2.3 b | 163.9 ± 5.5 a | *** | 174.0 ± 10.6 b | 168.0 ± 5.3 b | 213.7 ± 15.1 a | ** | 79.5 ± 2.9 c | 158.8 ± 2.9 a | 105.7 ± 11.0 b | *** | |||
| 2019 | 2020 | 2021 | Multifactorial Analysis | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | Urea | Nano-Urea | p | Control | Urea | Nano-Urea | p | Control | Urea | Nano-Urea | p | T (%) | S (%) | TxS (%) | |
| Asp | 2.06 ± 0.15 c | 2.43 ± 0.11 b | 3.06 ± 0.11 a | *** | 3.26 ± 0.13 | 3.23 ± 0.51 | 3.99 ± 0.26 | ns | 4.89 ± 0.27 a | 3.78 ± 0.15 b | 2.48 ± 0.08 c | *** | 2 | 37 *** | 56 *** |
| Glu | 4.12 ± 0.64 b | 4.34 ± 0.29 b | 7.68 ± 1.56 a | ** | 9.41 ± 2.83 | 8.48 ± 1.23 | 12.74 ± 1.56 | ns | 2.94 ± 0.21 a | 3.07 ± 0.33 a | 1.91 ± 0.18 b | ** | 7 ** | 75 *** | 9 *** |
| Asn+Ser | 1.26 ± 0.19 b | 1.32 ± 0.18 b | 2.80 ± 0.47 a | ** | 6.44 ± 1.43 b | 6.66 ± 0.64 b | 9.23 ± 1.23 a | * | 7.28 ± 0.45 b | 10.74 ± 0.60 a | 6.91 ± 0.33 b | *** | 4 ** | 80 *** | 13 *** |
| Gln | 2.23 ± 0.17 b | 2.53 ± 0.39 b | 5.57 ± 2.14 a | * | 5.68 ± 0.74 b | 5.74 ± 0.28 b | 7.69 ± 0.59 a | ** | 2.58 ± 0.50 b | 5.98 ± 0.70 a | 2.56 ± 0.17 b | *** | 13 *** | 42 *** | 33 *** |
| His | 3.37 ± 0.11 | 3.52 ± 0.21 | 4.02 ± 0.50 | ns | 5.64 ± 0.82 | 5.19 ± 0.38 | 6.51 ± 0.80 | ns | 4.25 ± 0.19 b | 7.17 ± 0.42 a | 3.41 ± 0.35 c | *** | 7 ** | 40 *** | 44 *** |
| Gly | 4.55 ± 0.08 b | 4.69 ± 0.06 b | 5.52 ± 0.22 a | *** | 7.66 ± 1.03 b | 7.61 ± 0.51 b | 9.68 ± 0.85 a | * | 4.58 ± 0.16 b | 6.08 ± 0.33 a | 4.74 ± 0.18 b | *** | 6 ** | 78 *** | 11 *** |
| Thr | 3.40 ± 0.14 | 3.48 ± 0.03 | 3.66 ± 0.13 | ns | 4.19 ± 0.22 | 4.00 ± 0.15 | 4.57 ± 0.31 | ns | 2.91 ± 0.13 b | 3.54 ± 0.12 a | 2.95 ± 0.09 b | *** | 3 * | 76 *** | 14 *** |
| β-Ala | 2.37 ± 0.03 b | 2.42 ± 0.12 b | 2.61 ± 0.08 a | * | 3.64 ± 0.78 | 3.29 ± 0.21 | 3.73 ± 0.02 | ns | 2.04 ± 0.11 b | 2.22 ± 0.08 a | 1.89 ± 0.03 b | ** | 0 | 85 *** | 4 |
| Arg | 4.29 ± 0.29 | 4.67 ± 0.14 | 5.28 ± 0.64 | ns | 10.82 ± 2.68 b | 10.87 ± 0.98 b | 15.65 ± 1.73 a | * | 2.90 ± 0.12 b | 6.44 ± 0.85 a | 3.43 ± 0.61 b | *** | 4 ** | 80 *** | 10 *** |
| GABA | 4.75 ± 0.21 b | 4.75 ± 0.12 b | 7.12 ± 1.45 a | * | 15.42 ± 3.95 b | 14.20 ± 1.31 b | 20.86 ± 2.02 a | * | 3.33 ± 0.43 c | 9.54 ± 1.39 a | 5.65 ± 0.66 b | *** | 6 ** | 79 *** | 10 *** |
| α-Ala | 2.18 ± 0.04 b | 2.30 ± 0.07 b | 3.03 ± 0.53 a | * | 5.49 ± 0.93 b | 5.33 ± 0.57 b | 8.18 ± 1.70 a | * | 1.30 ± 0.17 c | 3.71 ± 0.54 a | 2.20 ± 0.26 b | *** | 8 ** | 72 *** | 13 *** |
| Pro | 102.73 ± 35.67 b | 109.34 ± 22.22 b | 263.41 ± 57.22 a | ** | 564.93 ± 88.04 ab | 449.66 ± 59.60 b | 679.66 ± 48.61 a | * | 94.82 ± 26.79 b | 272.28 ± 19.55 a | 75.11 ± 23.36 b | *** | 3 ** | 81 *** | 13 *** |
| NH4+ | 2.52 ± 0.13 | 2.45 ± 0.07 | 5.73 ± 2.77 | ns | 4.08 ± 1.15 | 4.13 ± 0.65 | 6.33 ± 1.11 | ns | 3.17 ± 0.28 a | 3.34 ± 0.61 a | 2.31 ± 0.26 b | * | 19 * | 24 ** | 27 * |
| Tyr | 3.18 ± 0.35 b | 3.76 ± 0.08 b | 4.57 ± 0.54 a | * | 5.36 ± 0.30 b | 4.46 ± 0.06 c | 6.44 ± 0.68 a | ** | 2.64 ± 0.14 | 3.08 ± 0.43 | 2.82 ± 0.43 | ns | 11 *** | 72 *** | 11 *** |
| Val | 2.56 ± 0.07 | 2.46 ± 0.05 | 2.58 ± 0.09 | ns | 2.86 ± 0.13 b | 2.92 ± 0.02 b | 3.32 ± 0.19 a | * | 2.40 ± 0.21 a | 2.68 ± 0.14 a | 2.08 ± 0.04 b | ** | 1 | 63 *** | 28 *** |
| Met | 2.67 ± 0.37 | 2.84 ± 0.14 | 2.84 ± 0.13 | ns | 3.12 ± 0.25 | 3.18 ± 0.09 | 3.47 ± 0.27 | ns | 2.19 ± 0.13 a | 2.11 ± 0.10 a | 1.56 ± 0.04 b | *** | 0 | 83 *** | 10 ** |
| Cys | 3.18 ± 0.06 | 3.05 ± 0.11 | 3.15 ± 0.34 | ns | 4.61 ± 0.62 | 4.14 ± 0.27 | 3.51 ± 0.42 | ns | 5.58 ± 0.20 a | 5.14 ± 0.61 a | 3.88 ± 0.16 b | ** | 18 *** | 60 *** | 11 * |
| Iso | 3.62 ± 0.17 | 3.44 ± 0.34 | 3.96 ± 0.26 | ns | 4.72 ± 0.37 | 3.98 ± 0.37 | 4.47 ± 0.25 | ns | 2.61 ± 0.09 b | 2.89 ± 0.12 a | 2.40 ± 0.03 c | ** | 1 | 83 *** | 8 ** |
| Leu | 3.90 ± 0.10 b | 4.06 ± 0.02 b | 4.32 ± 0.17 a | * | 5.25 ± 0.56 | 4.97 ± 0.26 | 5.80 ± 0.31 | ns | 3.73 ± 0.12 b | 4.92 ± 0.25 a | 4.03 ± 0.17 b | *** | 7 ** | 63 *** | 20 *** |
| Trp | 4.03 ± 0.09 b | 4.13 ± 0.16 b | 4.59 ± 0.17 a | ** | 4.97 ± 0.21 | 4.81 ± 0.32 | 5.40 ± 0.28 | ns | 3.54 ± 0.28 b | 4.17 ± 0.16 a | 3.36 ± 0.08 b | ** | 3 * | 74 *** | 16 *** |
| Phe | 2.7 ± 0.28 | 3.10 ± 0.23 | 3.19 ± 0.17 | ns | 3.74 ± 0.24 | 3.80 ± 0.27 | 4.49 ± 0.57 | ns | 2.18 ± 0.22 b | 3.24 ± 0.36 a | 2.15 ± 0.10 b | ** | 8 ** | 66 *** | 16 ** |
| Orn | 1.97 ± 0.05 | 2.21 ± 0.10 | 2.34 ± 0.28 | ns | 3.11 ± 0.32 | 3.05 ± 0.15 | 3.62 ± 0.50 | ns | 1.48 ± 0.02 b | 1.64 ± 0.04 a | 1.52 ± 0.04 b | ** | 3 * | 88 *** | 3 |
| Lys | 4.25 ± 1.29 | 3.77 ± 0.04 | 4.99 ± 0.55 | ns | 8.64 ± 2.71 | 8.13 ± 1.27 | 10.24 ± 0.90 | ns | 2.68 ± 0.35 c | 6.77 ± 0.61 a | 4.69 ± 0.66 b | *** | 6 * | 67 *** | 14 * |
| Total | 171.90 ± 38.69 b | 181.07 ± 22.75 b | 356.03 ± 57.68 a | ** | 693.04 ± 108.50 ab | 571.83 ± 67.16 b | 839.58 ± 58.68 a | * | 166.03 ± 27.28 b | 374.54 ± 26.75 a | 144.00 ± 27.34 b | *** | 3 ** | 80 *** | 13 *** |
| Function 1 | Function 2 | |
|---|---|---|
| Asp | −4.470 | −4.528 |
| Glu | 3.265 | 2.850 |
| Asn+Ser | 11.249 | 10.866 |
| Gln | −8.503 | −4.487 |
| His | 17.221 | 2.873 |
| Gly | −4.831 | −2.567 |
| Thr | 7.317 | 4.214 |
| β-Ala | 5.397 | 2.069 |
| Arg+GABA | −13.709 | −5.698 |
| α-Ala | 5.186 | 2.527 |
| Pro | 2.150 | −0.303 |
| NH4+ | −7.363 | −2.951 |
| Tyr | −7.018 | 0.122 |
| Val | 19.354 | 0.668 |
| Met | 1.728 | 1.517 |
| Cys | 0.793 | 0.465 |
| Iso | −1.497 | −8.096 |
| Leu | −16.820 | 1.317 |
| Trp | −10.434 | −8.579 |
| Phe | 2.176 | 7.474 |
| Orn | 0.578 | 0.339 |
| Lys | −0.274 | 0.542 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giménez-Bañón, M.J.; Moreno-Olivares, J.D.; Bleda-Sánchez, J.A.; Gómez-Martínez, J.C.; Cebrián-Pérez, A.; Parra-Torrejón, B.; Ramírez-Rodríguez, G.B.; Delgado-López, J.M.; Gil-Muñoz, R. Foliar Treatments with Urea and Nano-Urea Modify the Nitrogen Profile of Monastrell Grapes and Wines. Horticulturae 2025, 11, 570. https://doi.org/10.3390/horticulturae11060570
Giménez-Bañón MJ, Moreno-Olivares JD, Bleda-Sánchez JA, Gómez-Martínez JC, Cebrián-Pérez A, Parra-Torrejón B, Ramírez-Rodríguez GB, Delgado-López JM, Gil-Muñoz R. Foliar Treatments with Urea and Nano-Urea Modify the Nitrogen Profile of Monastrell Grapes and Wines. Horticulturae. 2025; 11(6):570. https://doi.org/10.3390/horticulturae11060570
Chicago/Turabian StyleGiménez-Bañón, María José, Juan Daniel Moreno-Olivares, Juan Antonio Bleda-Sánchez, José Cayetano Gómez-Martínez, Ana Cebrián-Pérez, Belén Parra-Torrejón, Gloria Belén Ramírez-Rodríguez, José Manuel Delgado-López, and Rocío Gil-Muñoz. 2025. "Foliar Treatments with Urea and Nano-Urea Modify the Nitrogen Profile of Monastrell Grapes and Wines" Horticulturae 11, no. 6: 570. https://doi.org/10.3390/horticulturae11060570
APA StyleGiménez-Bañón, M. J., Moreno-Olivares, J. D., Bleda-Sánchez, J. A., Gómez-Martínez, J. C., Cebrián-Pérez, A., Parra-Torrejón, B., Ramírez-Rodríguez, G. B., Delgado-López, J. M., & Gil-Muñoz, R. (2025). Foliar Treatments with Urea and Nano-Urea Modify the Nitrogen Profile of Monastrell Grapes and Wines. Horticulturae, 11(6), 570. https://doi.org/10.3390/horticulturae11060570