1. Introduction
Juglans mandshurica Maxim. is a valuable deciduous hardwood species native to Northeast China. Its natural range extends from Jilin and Liaoning provinces westward to Inner Mongolia and southward to Shanxi, Henan, Hebei, and Shandong. In Heilongjiang Province, it occurs in mountainous regions including the Lesser Khingan Range and Wanda Mountains, becoming less common in northern areas. Internationally, the species is also found in North Korea, the Russian Far East, and Japan [
1].
As a valuable tree species in Northeast China, the
J. mandshurica is widely utilized in the production of high-end furniture and woodworking crafts due to its dense hardwood and attractive grain [
2]. The seeds are highly nutritious, containing various essential fatty acids and amino acids required by the human body [
3]. Additionally, the fresh root bark, branch bark, and immature fruit peel are rich in terpenes, flavonoids, quinones, and phenolic compounds, which have significant antibacterial and antitumor properties [
4]. Therefore, germplasm selection should simultaneously consider the synergistic optimization of medicinal compound content and fruit agronomic traits to achieve the dual enhancement of both oil production and medicinal value. The natural forests of
J. mandshurica in Northeast China have experienced extensive logging and destruction due to their high economic and medicinal value, leading to a significant decline in population numbers. This species was once classified as a Level II rare species, an endangered species, and a Level III protected plant in China [
5]. The existing genetic resources have been severely depleted, posing challenges to its sustainable utilization. Currently, with the growing awareness of natural forest conservation,
J. mandshurica germplasm resources have also recovered to a certain degree. However, due to its slow growth and the challenges of vegetative propagation and hybrid breeding, the research primarily focuses on growth traits [
6] (tree height, diameter at breast height) and other target traits [
7] (wood mechanical properties). Research on other target traits remains limited, leading to a narrow improvement focus and slow progress in genetic enhancement. To advance the development and conservation of existing germplasm resources, foster the cultivation of elite varieties, and accelerate the genetic improvement of
J. mandshurica, it is essential to conduct targeted studies on fruit traits. Identifying and selecting families with elite fruit characteristics can significantly increase walnut oil yield, providing both economic and ecological benefits. This approach will support the development of high-yielding, oil-rich varieties, ensuring the sustainable improvement and utilization of
J. mandshurica for walnut oil production.
The phenotypic traits of plant populations are influenced by geographic and environmental conditions. Understanding the characteristics and geographic distribution of a species provides valuable insights into the diversity of germplasm resources, genetic variation, and phylogenetic relationships across different regions [
8]. This knowledge is essential for the regional classification of germplasm resources and is crucial in formulating rational development, utilization plans, and breeding strategies for
J. mandshurica [
9,
10]. Research indicates that
J. mandshurica exhibits abundant phenotypic variation in fruit traits across and within different regions, revealing a geographic variation trend from southeast to northwest. The genetic variation coefficients among provenances range from 1.86% to 10.56% [
11]. Therefore, the aim of this study is to analyze the genetic variation patterns of the fruit traits in
J. mandshurica. By analyzing 13 key traits, it also offers a reference for the selection of superior germplasm with high oil content. The specific research objectives are as follows: (1) quantify the contributions of genetic and environmental factors to trait variation; (2) investigate the correlations among high-yield, high-quality, and high-adaptability traits to reveal synergistic and trade-off relationships in breeding selection; (3) utilizing principal component analysis and other multi-indicator evaluation methods, this study aims to select superior germplasm resources with high yield, stable production, and strong adaptability; (4) integrate genetic diversity analysis results to propose germplasm conservation and sustainable utilization strategies, providing a scientific basis for variety improvement and resource protection.
4. Discussion
The selection process is the most critical aspect of elite cultivar breeding. The greater the variation in traits, among and within provenances, the more potential there is for selective breeding [
22]. The selection of elite provenances will inevitably lead to a narrowing of genetic diversity. Therefore, it is necessary to continuously supplement new elite provenances in the breeding population [
23]. To investigate the phenotypic variation in fruit traits of
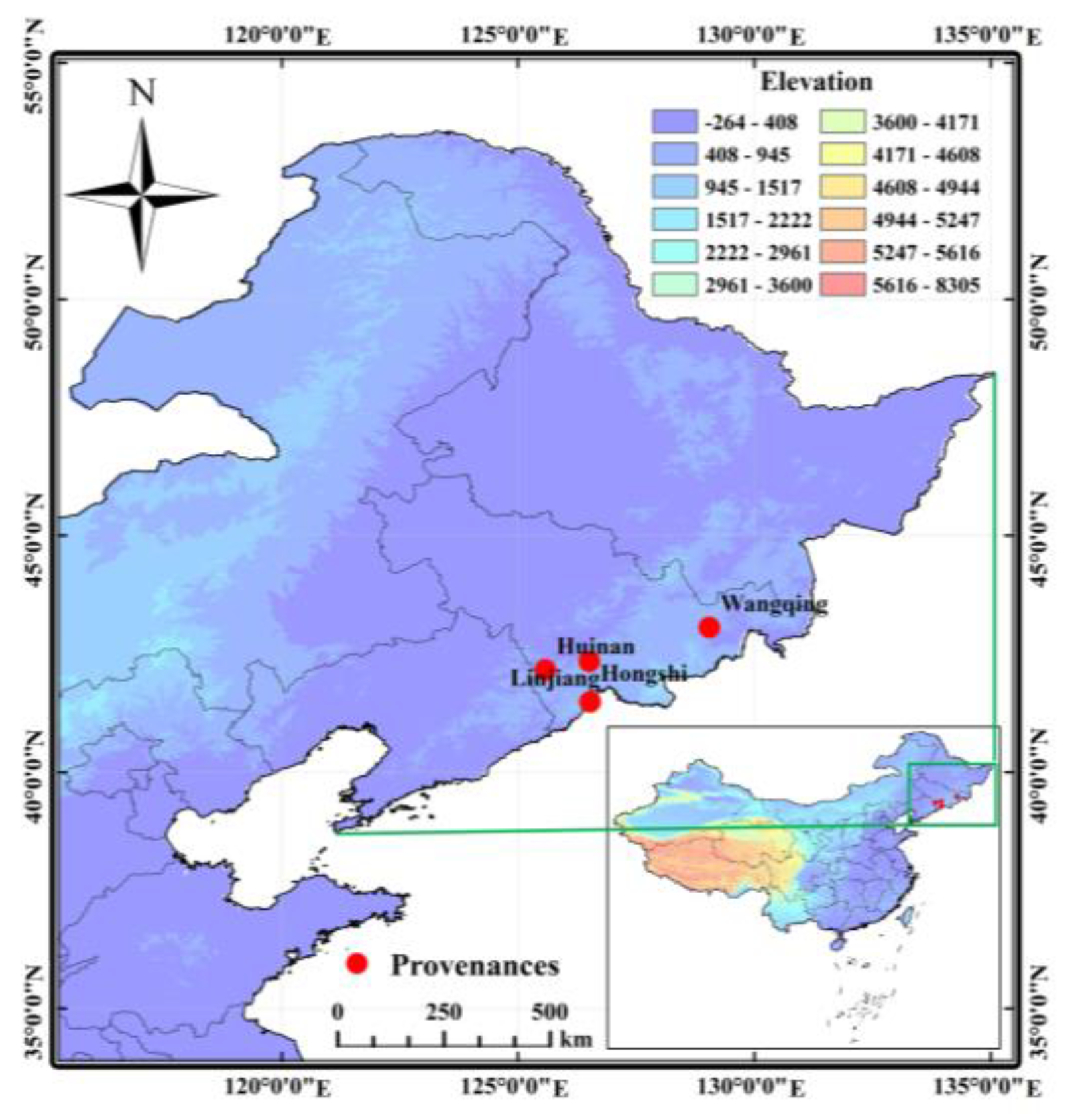
J. mandshurica from different provenances, this study conducted an analysis of 13 fruit traits across four provenances in Jilin Province. The results of the variance analysis exhibited highly significant differences for all traits, both among provenances and among families. This finding is consistent with the research by Yu on
J. mandshurica, which also identified substantial and significant variation in fruit traits across provenances and families [
24]. These findings suggested the potential for developing elite provenances and families, providing a basis for germplasm resource evaluation. This demonstrates that the significant phenotypic variations in
J. mandshurica fruits across provenances and families provide valuable genetic resources for elite cultivar selection while necessitating the ongoing diversification of breeding populations to preserve genetic diversity.
The coefficient of variation is an important measure used to assess the degree of trait variation, which plays a crucial role in the breeding of elite tree species [
25]. In this study, traits with relatively low genetic coefficients of variation included fruit length, fruit width, fruit type index, and five other traits (with genetic coefficients of variation below 10%). Conversely, the traits with higher genetic coefficients of variation were nut weight, kernel weight, and seed yield, suggesting that these traits can be relatively stable and inherited by the next generation. When examining phenotypic coefficients of variation, lower values were observed for fruit length, fruit width, and the three-diameter mean (all below 10%). In contrast, traits with higher phenotypic coefficients of variation, such as nut weight, kernel weight, seed yield, and kernel yield, indicated significant potential as selection indicators due to their rich phenotypic diversity. Furthermore, the study demonstrated that the GCV accounted for a large proportion of the PCV for all traits, which aligns with the findings of Li in their analysis of fruit traits across 12 provenances [
26]. This suggested that the growth variation among families is mainly controlled by genetic factors, providing valuable insights for the selection of elite provenances and families. The mean data indicated significant differences in fruit traits among various provenances and families of
J. mandshurica. In this study, the mean values for various fruit and nut phenotypic traits of
J. mandshurica in Jilin Province were higher than those of Carya cathayensis in the Dabie Mountains of Anhui Province [
27]. However, the mean fruit type index and kernel yield were lower than those of walnuts from Guizhou Province [
28]. This may be due to geographic differences, suggesting that geographic and environmental factors can influence the fruit traits of
J. mandshurica. Heritability reflects the ability of a parent plant to pass a trait to its offspring. Moreover, a higher heritability indicates that the trait can be stably inherited. In this study, the heritability range of
J. mandshurica families was 0.968–0.988, indicating a higher level of heritability, which exceeds the heritability of growth traits in
Fraxinus mandshurica [
29]. This indicated that the fruit traits of
J. mandshurica can be stably inherited. Additionally, fruit weight and nut weight exhibited the highest heritability, suggesting that these two traits possess the greatest potential for genetic gain. The findings confirm that
J. mandshurica exhibits highly heritable fruit traits with substantial genetic variation, particularly in nut-related characteristics, making them ideal targets for selective breeding programs.
Mean analysis is a statistical method that reflects the average level of a trait within a population. Due to the interaction between genetics and the environment, significant differences in traits exist among different groups. By comparing means, the dependencies between traits can be analyzed [
30]. In this study, the Hongshi provenance demonstrated the highest mean values for eight fruit traits among all provenances, such as fruit width and nut longitudinal diameter, while it exhibited the lowest mean fruit type index. Conversely, the Wangqing provenance recorded the lowest mean values for nine fruit traits, including fruit length and fruit width. However, it recorded the highest kernel yield among all provenances. The reason may be that, to support kernel development, the fruit and nut reduce their own weight to ensure sufficient energy for kernel growth, thereby increasing the kernel yield. In addition, differences in geographic location among provenances may lead to considerable variability in the mean values of fruit traits in
J. mandshurica. Similarly, a study by Han on Acer truncatum found significant differences in seed traits among provenances [
31], indicating abundant variation in fruit traits. This diversity is beneficial for the evaluation and selection of
J. mandshurica from different provenances. The results demonstrate significant differences in fruit traits among
J. mandshurica provenances, with the Hongshi provenance exhibiting optimal performance in most morphological traits while the Wangqing provenance shows the greatest potential in kernel yield.
The relationships between variables can be represented through correlation analysis. In a study by Pang, it is suggested that comprehensive analysis is elite to direct correlation analysis, as multiple indicators are needed to reflect the correlations between traits, and environmental factors can influence the physiological growth traits of trees [
32]. Furthermore, the research by Tore confirmed that geographical and ecological conditions also play a decisive role in determining the fruit quality of plants [
33]. In this study, correlation analysis of various traits revealed that nut longitudinal diameter exhibited a high positive correlation with fruit length, and the three-diameter mean was highly positively correlated with nut longitudinal, transverse, and side diameters, with correlation coefficients of 0.872, 0.826, and 0.814, respectively, all reaching highly significant levels. This suggests that the fruit phenotype influences nut development, likely because a larger fruit volume provides more space within the fruit for nut growth, resulting in a greater longitudinal diameter. Additionally, the nut roundness index demonstrated a high negative correlation with fruit length, fruit type index, and nut longitudinal diameter, with correlation coefficients of −0.531, −0.594, and −0.585, respectively, all reaching highly significant levels. This suggested that the fruit type index influences the nut roundness index: as the fruit length increases, the transverse and side diameters of the nut decrease, further indicating that the fruit phenotype affects nut development. Correlation analysis with environmental factors revealed a significant negative correlation between longitude and seed yield, while other traits did not reach significant levels with geographical conditions. This indicates that longitude has a substantial effect on seed yield, possibly because the lower longitudes of these four provenances are associated with higher precipitation and longer sunlight exposure, which in turn increase nut weight, thereby boosting seed yield. Additionally, the correlation coefficients between annual rainfall and various traits were relatively high, further suggesting that annual rainfall significantly impacts fruit traits. Conversely, altitude demonstrated the lowest correlation with the traits, indicating a minimal effect on fruit traits. Also, a study by Deng shows that the seedling stage is most sensitive to external environments during tree regeneration [
34]. These results further confirm that geographical and environmental factors influence the traits of
J. mandshurica fruit and seeds. This may be because
J. mandshurica adjusts its phenotype to adapt to external environmental conditions, allowing more seeds space for growth and development. The findings conclusively demonstrate that both intrinsic trait correlations and extrinsic geographical factors significantly shape the phenotypic expression of
J. mandshurica fruit characteristics, and longitude may affect seed traits.
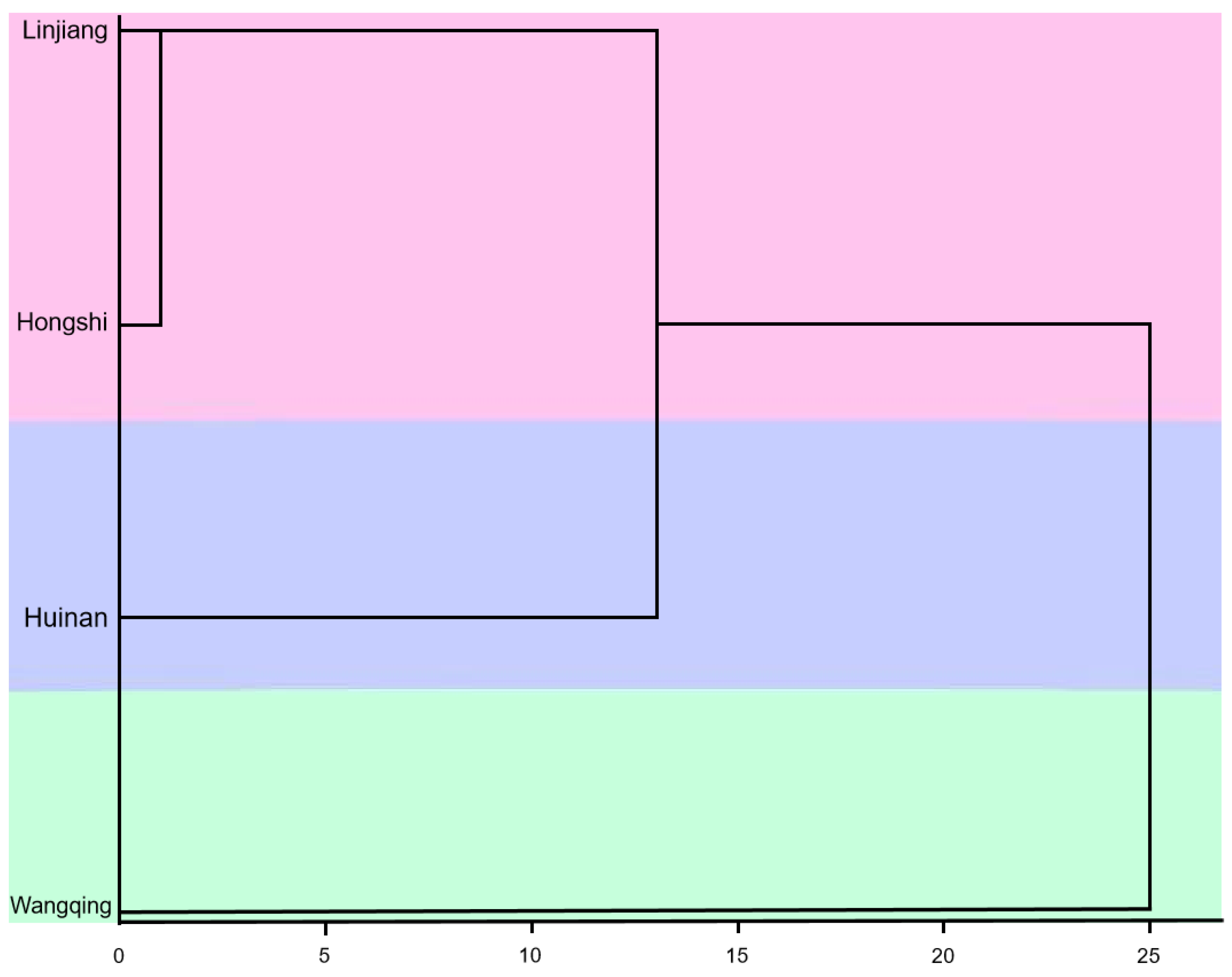
Cluster analysis in data analysis enables the discovery of underlying structures and patterns in complex datasets. Selecting the appropriate clustering algorithm and parameter settings is critical for the validity and reliability of the results of these analyses. Cluster analysis based on squared Euclidean distance can categorize materials from different provenances according to distinct trait types [
35], thereby enhancing the value of each type of
J. mandshurica fruit seed. In this study, cluster analysis was conducted on the fruit traits of four
J. mandshurica provenances. When the genetic distance was set to 5, the analysis grouped the materials into three clusters. The first cluster included the Hongshi and Linjiang provenances, which have higher mean values for nine traits, including fruit length and fruit width, and a lower mean fruit type index. This suggested that materials in this group exhibit prominent fruit phenotypic traits with larger weights. The second cluster included the Huinan provenance, characterized by a higher nut roundness index and seed yield, as well as lower fruit weight and kernel yield. This indicated that materials in this group have prominent nut phenotypic traits and a rounder nut shape. The third cluster included the Wangqing provenance, characterized by a higher fruit type index and kernel yield, indicating that materials in this group have distinct kernel traits and a higher kernel yield. The cluster analysis effectively classified
J. mandshurica provenances into three distinct groups based on fruit characteristics, with each cluster exhibiting unique combinations of superior traits for either fruit morphology, nut quality, or kernel production potential.
PCA can condense multiple variables into a few representative indicators, clearly reflecting the original set of variables [
36]. In this study, the eigenvalues of the four principal components were 5.08, 2.80, 1.91, and 1.53, with a cumulative contribution rate as high as 87.00%, encompassing most of the information on fruit traits. PCI demonstrated the highest contribution rate and included traits such as fruit length, nut longitudinal diameter, nut transverse diameter, nut side diameter, three-diameter mean, and nut weight. This indicated that the morphology and weight of the nut have a significant impact on the fruit phenotype. This may be because the fruit needs to provide the necessary nutrients and space for nut growth, leading to changes in fruit length and width based on nut morphology. The three-diameter mean of the nut affects kernel development as a larger three-diameter allows more space for kernel growth, while smaller means restrict it, resulting in variations among provenances. Finally, the linear equations were established based on the two principal components, which carried out the comprehensive evaluation of multiple traits, and selected one elite provenance and five elite families. The results revealed that the Hongshi provenance achieved the highest comprehensive score, establishing it as an elite provenance. With a selection rate of 5%, five families, including HS-08 and HS-18, were selected as elite families. Among these, five elite families, such as LJ-06 and LJ-15, belong to the elite provenance and hold the potential for further promotion and application in
J. mandshurica cultivation. The selected families performed excellently in kernel weight, indicating a significant increase in kernel production, high genetic gain, and the capacity to produce more
J. mandshurica kernels for refining oil. Future research could further explore the correlations between phenotypic traits and environmental factors to better elucidate the influence of environmental conditions on the growth and development of
J. mandshurica. Additionally, in-depth studies on the genetic mechanisms underlying these traits will help accelerate breeding processes and enhance crop production efficiency.




{kind=link}
{kind=link}
{kind=link}