
The Influence of Varying Wavelengths of LED Light on the Development, Physiology Response, and Metabolism Activities of Micropropagated Dendrobium Hybrid ‘Shuijing’ Plantlets

Abstract
1. Introduction
2. Materials and Methods
2.1. General Growth Conditions
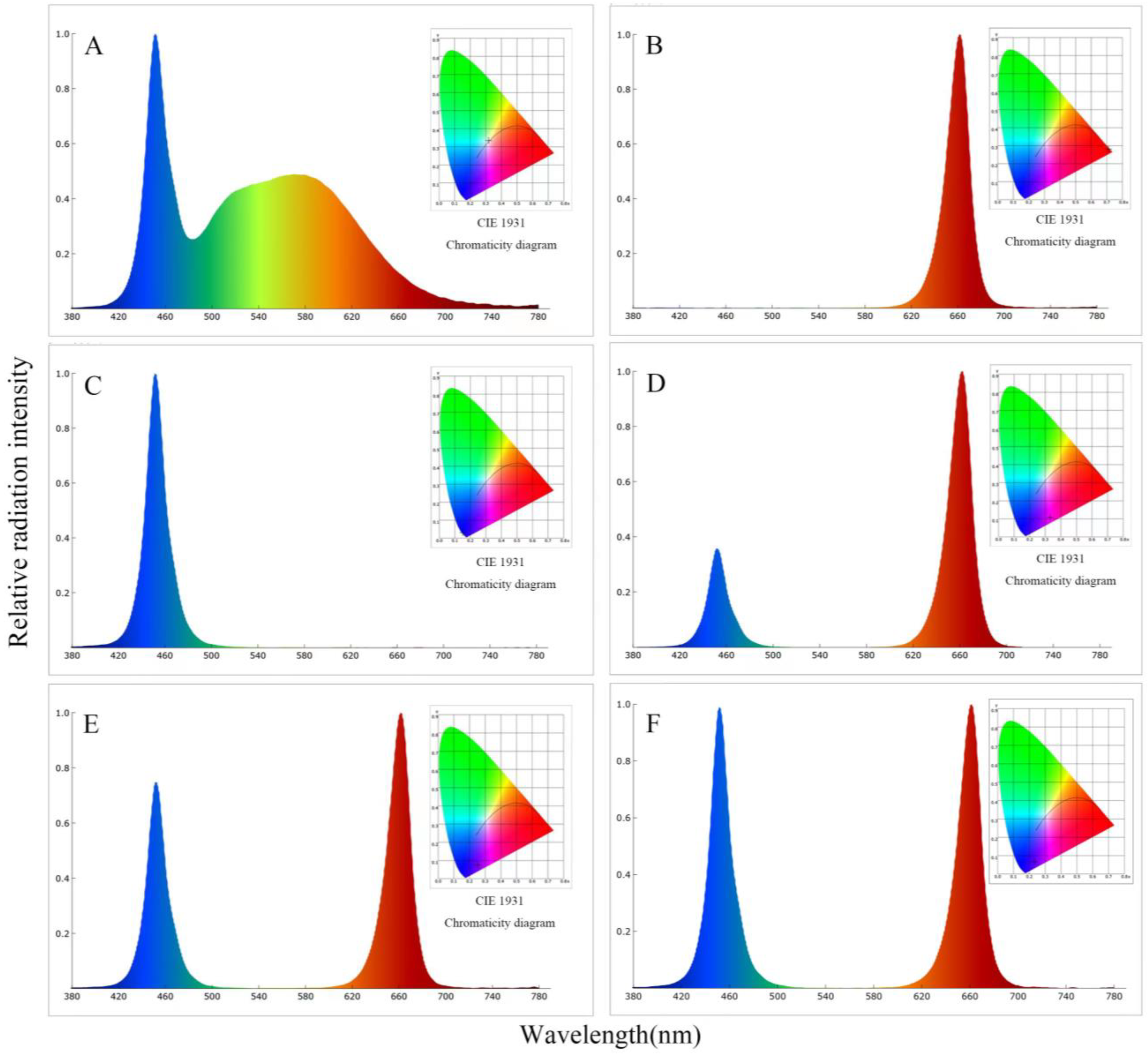
2.2. Experimental Design
2.3. Morphological Assessment
2.4. Assessment of Root Activity
2.5. Antioxidant System and Nutritional Indicators
2.6. Chlorophyll and Carotenoid Contents
2.7. Statistical Analysis
3. Results
3.1. Impact of Diverse Light Spectrums on the Growth Characteristics of ‘Shuijing’ Dendrobium Hybrid Plantlets Cultured In Vitro
3.2. Effects of Different Light Qualities on the Contents of Soluble Sugars (SSC), Soluble Proteins (SPC), and Free Proline of Dendrobium Hybrid ‘Shuijing’ Planets In Vitro
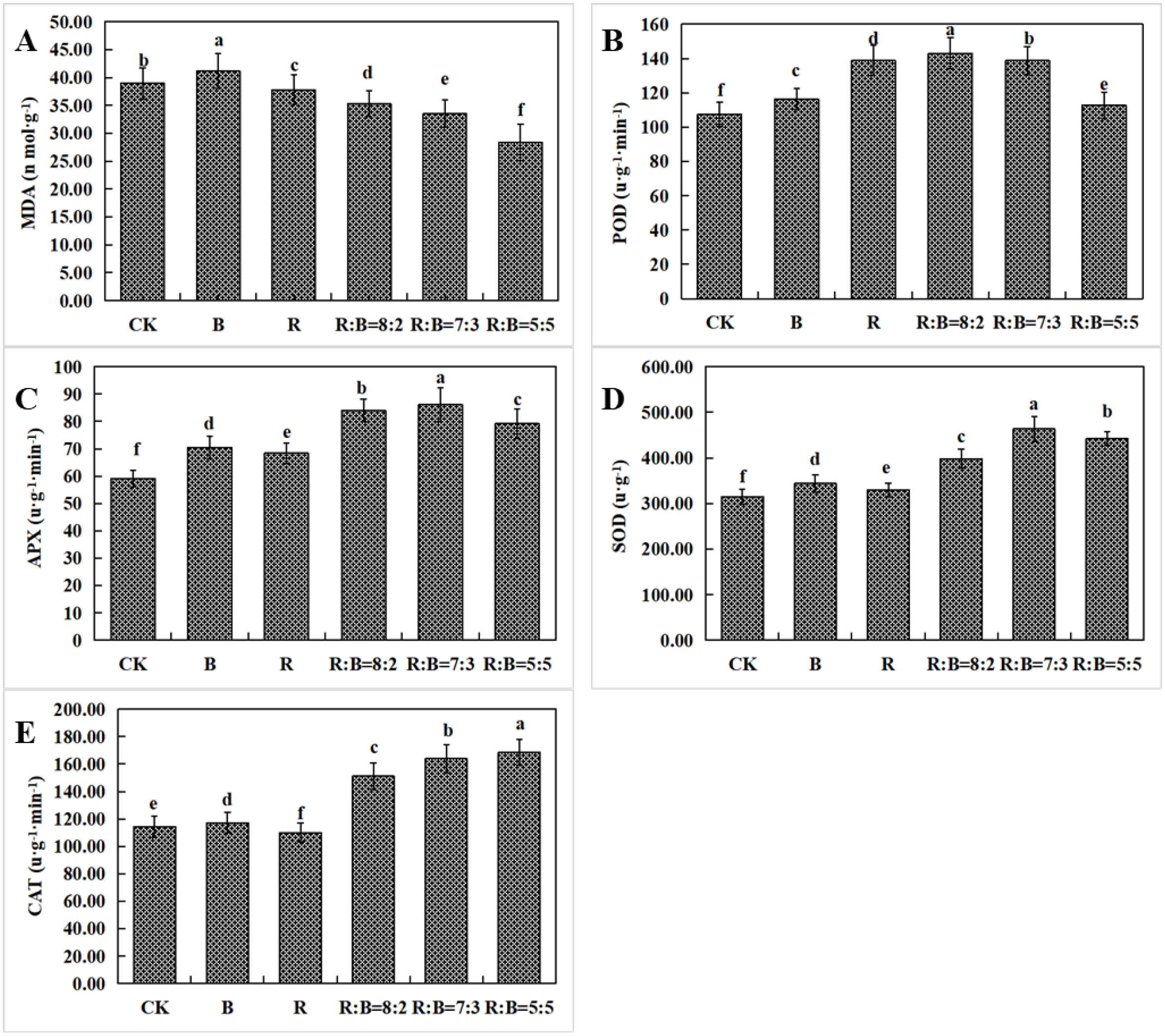
3.3. Effects of Various Light Spectrums on the In Vitro Antioxidant Capacity of Leaves from Dendrobium Hybrid ‘Shuijing’ Plantlets
3.4. Impact of Diverse Light Qualities on the Concentrations of Chlorophyll and Carotenoids in Cultured Dendrobium Hybrid Plantlets
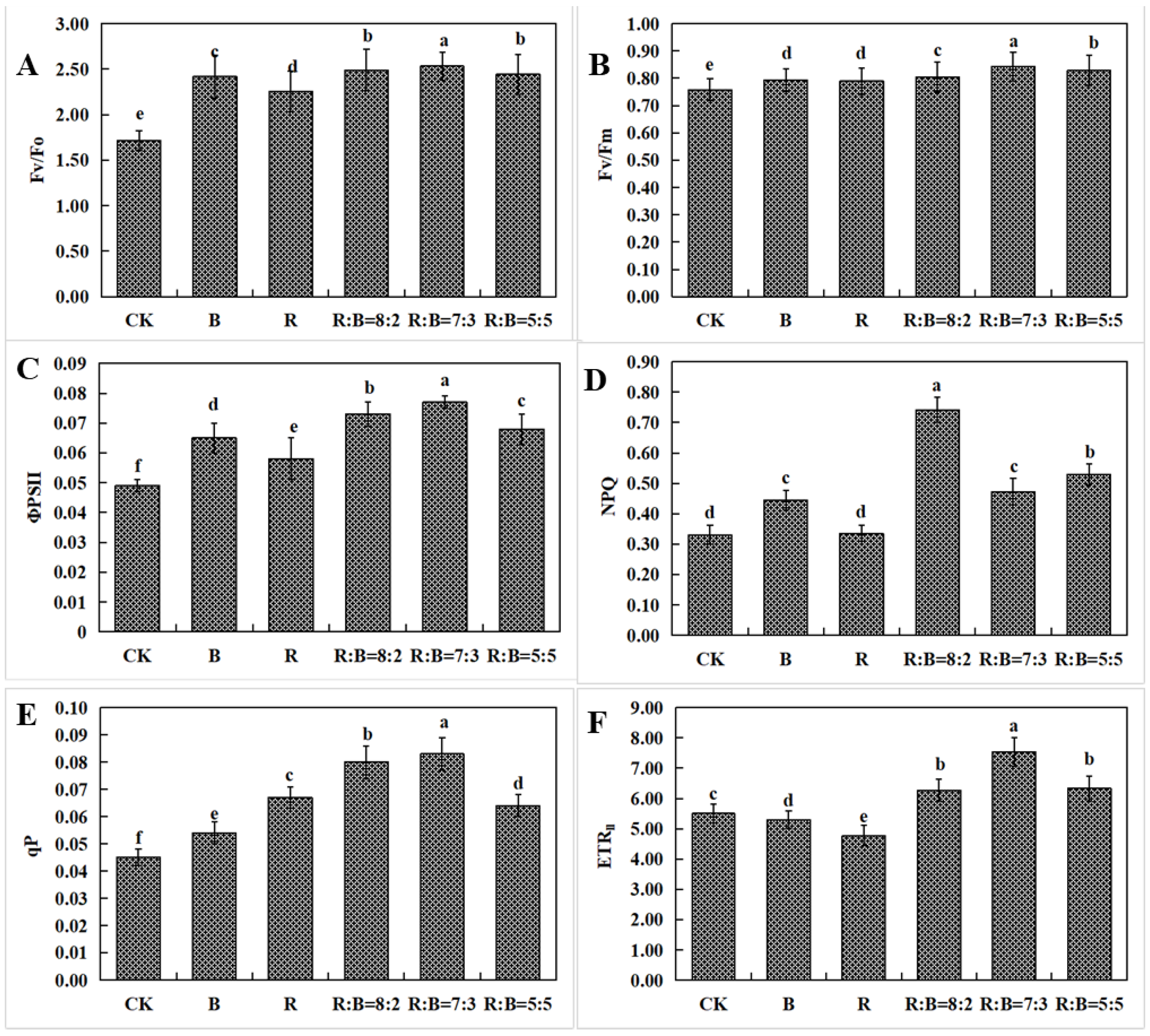
3.5. The Influence of Various Light Characteristics on the Fluorescence of Chlorophyll in Dendrobium Hybrid Plantlets Cultivated in a Controlled Laboratory Setting
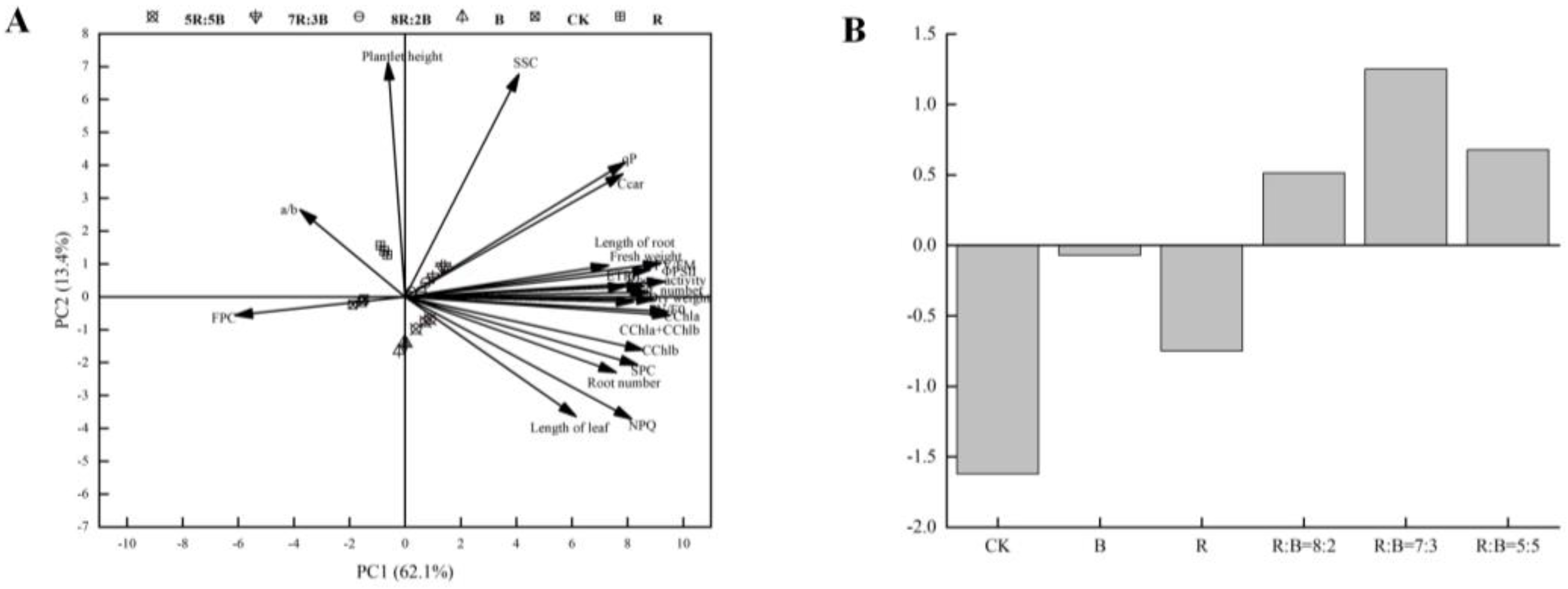
3.6. Relationship between Growth and Antioxidant Parameters and the Chlorophyll and Fluorescence Parameters of Dendrobium Hybrid Plantlets Grown In Vitro
3.7. Linear Regression Models Were Constructed by Integrating Physiological and Biochemical Parameters, along with Chlorophyll Fluorescence Measurements
3.8. Comprehensive Scores of Dendrobium Hybrid Plantlets Grown In Vitro
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Muneer, S.; Kim, E.J.; Park, J.S.; Lee, J.H. Influence of green, red and blue light emitting diodes on multiprotein complex proteins and photosynthetic activity under different light intensities in lettuce leaves (Lactuca sativa L.). Int. J. Mol. Sci. 2014, 15, 4657–4670. [Google Scholar] [CrossRef] [PubMed]
- Barceló Muñoz, A.; Barceló Muñoz, M.; Gago Calderon, A. Effect of LED lighting on physical environment and microenvi-ronment on In vitro plant growth and morphogenesis: The need to standardize lighting conditions and their description. Plants 2021, 11, 60. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Yang, M.; Cheng, F.; Liu, S.; Liang, Y. Effects of LED photoperiods and light qualities on in vitro growth and chlorophyll fluorescence of Cunninghamia lanceolata. Plant Biol. 2020, 23, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Dutta Gupta, S.; Kumar, A.; Agarwal, A. Impact of light-emitting diodes (LEDs) on the growth and morphogenesis of encapsulated shoot buds of Curculigo orchioides Gaertn., an endangered medicinal herb. Acta Physiol. Plant. 2019, 41, 50. [Google Scholar] [CrossRef]
- Akcin, A.; Yalcin, E. Effect of salinity stress on chlorophyll, carotenoid content, and proline in Salicornia prostrata Pall. and Suaeda prostrata Pall. subsp. prostrata (Amaranthaceae). Braz. J. Bot. 2015, 39, 101–106. [Google Scholar] [CrossRef]
- Fan, X.; Zang, J.; Xu, Z.; Guo, S.; Jiao, X.; Liu, X.; Gao, Y. Effects of different light quality on growth, chlorophyll concentration and chlorophyll biosynthesis precursors of non heading chinese cabbage (Brassica campestris L.). Acta Physiol. Plant. 2013, 35, 2721–2726. [Google Scholar] [CrossRef]
- Naznin, M.T.; Lefsrud, M.; Gravel, V.; Azad, O.K. Blue light added with red LEDs enhance growth characteristics, pigments content, and antioxidant capacity in lettuce, spinach, kale, basil, and sweet pepper in a controlled environment. Plants 2019, 8, 93. [Google Scholar] [CrossRef] [PubMed]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue light dose–responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Kwon, A.-R.; Cui, H.-Y.; Lee, H.; Shin, H.; Kang, K.-S.; Park, S.-Y. Light quality affects shoot regeneration, cell division, and wood formation in elite clones of Populus euramericana. Acta Physiol. Plant. 2015, 37, 65. [Google Scholar] [CrossRef]
- Silvestri, C.; Caceres, M.E.; Ceccarelli, M.; Pica, A.L.; Rugini, E.; Cristofori, V. Influence of continuous spectrum light on morphological traits and leaf anatomy of hazelnut plantlets. Front. Plant Sci. 2019, 10, 1318. [Google Scholar] [CrossRef]
- Khairul-Anuar, M.-A.; Mazumdar, P.; Lum, S.; Harikrishna, J.A. Dendrobium hybrid flower number and pedicel curvature is influenced by the application of gibberellic acid and indole-3-acetic acid. Biol. Bull. 2021, 48, 740–745. [Google Scholar] [CrossRef]
- Guo, Y.; Zhong, Y.; Mo, L.; Zhang, W.; Chen, Y.; Wang, Y.-C.; Chen, H.; Wang, Z.; Song, X.; Meng, X. Different combinations of red and blue LED light affect the growth, physiology metabolism and photosynthesis of in vitro-cultured Dendrobium nobile ‘Zixia’. Hortic. Environ. Biotechnol. 2023, 64, 393–407. [Google Scholar] [CrossRef]
- Fan, C.; Manivannan, A.; Wei, H. Light quality-mediated influence of morphogenesis in micropropagated horticultural crops: A comprehensive overview. BioMed Res. Int. 2022, 2022, 4615079. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xu, J.-Q.; Meng, F.-L.; Zhang, Y.-H.; Wang, Z.-X.; Zhao, Q.-H.; Xu, S. Impact of drought stress on the rhizosphere bacterial community structure of Capsicum annuum in Qiubei. CCV 2024, 7, 1–16. [Google Scholar]
- Li, H. Principles and techniques of biochemical experiments in plant physiology. High. Educ. Presse 2000, 258, 195–197. [Google Scholar]
- Gao, J. Experimental Guide to Plant Physiology; Higher Education Press: Beijing, China, 2006. [Google Scholar]
- Sun, M.; Liu, L. Correlation analysis of leaf chlorophyll content, malondialdehyde content and alkali damage index under alkali stress in lily seedlings. North. Hortic. 2018, 20, 93–101. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Yang, Y.; Gou, R.-J.; Fu, L.-Z.; Wang, M.; Wang, D.-S.; Zhang, H.-E. Comparison of photosynthetic characteristics and pigment content of red Chinese chestnut and green Chinese chestnut leaves. Chin. Fruits 2024, 4, 62–67. [Google Scholar]
- Xu, L.-F.; Xu, C.-L.; Li, R.-D.; Wu, Z.-S.; Hua, J.-X.; Yang, L.; Song, W.-W.; Wu, C.-X. Deep side fertilization improved soybean yield by optimizing leaf function and nitrogen accumulation. Acta Agron. Sin. 2024, 75, 1–11. [Google Scholar]
- Kuo, T.C.-Y.; Chen, C.-H.; Chen, S.-H.; Lu, I.-H.; Chu, M.-J.; Huang, L.-C.; Lin, C.-Y.; Chen, C.-Y.; Lo, H.-F.; Jeng, S.-T.; et al. The effect of red light and far-red light conditions on secondary metabolism in agarwood. BMC Plant Biol. 2015, 15, 139. [Google Scholar] [CrossRef]
- Miyagi, A.; Uchimiya, H.; Kawai-Yamada, M. Synergistic effects of light quality, carbon dioxide and nutrients on metabolite compositions of head lettuce under artificial growth conditions mimicking a plant factory. Food Chem. 2017, 218, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Ptak, A.; Morańska, E.; Saliba, S.; Zieliński, A.; Simlat, M.; Laurain-Mattar, D. Elicitation of galanthamine and lycorine biosynthesis by Leucojum aestivum L. and L. aestivum ‘Gravety Giant’ plants cultured in bioreactor RITA®. Plant Cell Tissue Organ Cult. (PCTOC) 2016, 128, 335–345. [Google Scholar] [CrossRef]
- Cioć, M.; Szewczyk, A.; Żupnik, M.; Kalisz, A.; Pawłowska, B. LED lighting affects plant growth, morphogenesis and phytochemical contents of Myrtus communis L. in vitro. Plant Cell Tissue Organ Cult. (PCTOC) 2017, 132, 433–447. [Google Scholar] [CrossRef]
- Younas, M.; Drouet, S.; Nadeem, M.; Giglioli-Guivarc, H.N.; Hano, C.; Abbasi, B.H. Differential accumulation of silymarin induced by exposure of Silybum marianum L. callus cultures to several spectres of monochromatic lights. J. Photochem. Photobiol. B Biol. 2018, 184, 61–70. [Google Scholar] [CrossRef]
- Li, Q.; Xu, J.; Yang, L.; Sun, Y.; Zhou, X.; Zheng, Y.; Zhang, Y.; Cai, Y. LED light quality affect growth, alkaloids contents, and expressions of amaryllidaceae alkaloids biosynthetic pathway genes in Lycoris longituba. J. Plant Growth Regul. 2022, 41, 257–270. [Google Scholar] [CrossRef]
- He, J.; Qin, L.; Chong, E.L.C.; Choong, T.-W.; Lee, S.K. Plant growth and photosynthetic characteristics of Mesembryanthemum crystallinum grown aeroponically under different blue- and red-LEDs. Front. Plant Sci. 2017, 8, 361. [Google Scholar] [CrossRef] [PubMed]
- Lobiuc, A.; Vasilache, V.; Oroian, M.; Stoleru, T.; Burducea, M.; Pintilie, O.; Zamfirache, M.-M. Blue and red LED illumination improves growth and bioactive compounds contents in acyanic and cyanic Ocimum basilicum L. microgreens. Molecules 2017, 22, 2111. [Google Scholar] [CrossRef]
- Yu, L.-L.; Song, C.-M.; Sun, L.-J.; Li, L.-L.; Xu, Z.-G.; Tang, C.-M. Effects of light-emitting diodes on tissue culture plantlets and seedlings of rice (Oryza sativa L.). J. Integr. Agric. 2020, 19, 1743–1754. [Google Scholar] [CrossRef]
- Matsuda, R.; Ohashi-Kaneko, K.; Fujiwara, K.; Goto, E.; Kurata, K. Photosynthetic characteristics of rice leaves grown under red light with or without supplemental blue light. Plant Cell Physiol. 2004, 45, 1870–1874. [Google Scholar] [CrossRef]
- Lin, K.-H.; Huang, M.-Y.; Huang, W.-D.; Hsu, M.-H.; Yang, Z.-W.; Yang, C.-M. The effects of red, blue, and white light-emitting diodes on the growth, development, and edible quality of hydroponically grown lettuce (Lactuca sativa L. var. capitata). Sci. Hortic. 2013, 150, 86–91. [Google Scholar] [CrossRef]
- Lee, S.H.; Tewari, R.K.; Hahn, E.J.; Paek, K.Y. Photon flux density and light quality induce changes in growth, stomatal development, photosynthesis and transpiration of Withania somnifera (L.). Plant Cell Tissue Organ Cult. (PCTOC) 2007, 90, 141–151. [Google Scholar] [CrossRef]
- Wang, H.; Gu, M.; Cui, J.; Shi, K.; Zhou, Y.; Yu, J. Effects of light quality on CO2 assimilation, chlorophyll-fluorescence quenching, expression of Calvin cycle genes and carbohydrate accumulation in Cucumis sativus. J. Photochem. Photobiol. B Biol. 2009, 96, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Liang, Y.; Yang, M. Effects of composite LED light on root growth and antioxidant capacity of Cunninghamia lanceolata tissue culture seedlings. Sci. Rep. 2019, 9, 9766. [Google Scholar] [CrossRef] [PubMed]
- Shohael, A.; Ali, M.; Yu, K.; Hahn, E.; Islam, R.; Paek, K. Effect of light on oxidative stress, secondary metabolites and induction of antioxidant enzymes in Eleutherococcus senticosus somatic embryos in bioreactor. Process Biochem. 2006, 41, 1179–1185. [Google Scholar] [CrossRef]
- Ahmadi, T.; Shabani, L.; Sabzalian, M.R. LED light mediates phenolic accumulation and enhances antioxidant activity in Melissa officinalis L. under drought stress condition. Protoplasma 2020, 257, 1231–1242. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Fu, Y.; Liu, G.; Liu, H. Growth, photosynthetic characteristics, antioxidant capacity and biomass yield and quality of wheat (Triticum aestivum L.) exposed to LED light sources with different spectra combinations. J. Agron. Crop Sci. 2014, 200, 219–230. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Gritz, Y.; Merzlyak, M.N. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for non-destructive chlorophyll assessment in higher plant leaves. J. Plant Physiol. 2003, 160, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Hamedalla, A.M.; Ali, M.M.; Ali, W.M.; Ahmed, M.A.A.; Kaseb, M.O.; Kalaji, H.M.; Gajc-Wolska, J.; Yousef, A.F. Increasing the performance of cucumber (Cucumis sativus L.) seedlings by LED illumination. Sci. Rep. 2022, 12, 852–864. [Google Scholar] [CrossRef]
- Li, H.; Tang, C.; Xu, Z. Effects of different light quality on growth, photosynthetic characteristic and chloroplast ultrastructure of upland cotton (Gossypium hirsutum L.) seedlings. Emir. J. Food Agric. 2017, 29, 104. [Google Scholar] [CrossRef]
- Jarillo, J.A.; Gabrys, H.; Capel, J.; Alonso, J.M.; Ecker, J.R.; Cashmore, A.R. Phototropin-related NPL1 controls chloroplast relocation induced by blue light. Nature 2001, 410, 952–954. [Google Scholar] [CrossRef]
- Batista, D.S.; Felipe, S.H.S.; Silva, T.D.; de Castro, K.M.; Mamedes-Rodrigues, T.C.; Miranda, N.A.; Ríos-Ríos, A.M.; Faria, D.V.; Fortini, E.A.; Chagas, K. Light quality in plant tissue culture: Does it matter? Vitr. Cell. Dev. Biol. Plant 2018, 54, 195–215. [Google Scholar] [CrossRef]
- Folta, K.M.; Childers, K.S. Light as a growth regulator: Controlling plant biology with narrow-bandwidth solid-state lighting systems. HortScience 2008, 43, 1957–1964. [Google Scholar] [CrossRef]
- Yang, X.; Xu, H.; Shao, L.; Li, T.; Wang, Y.; Wang, R. Response of photosynthetic capacity of tomato leaves to different LED light wavelength. Environ. Exp. Bot. 2018, 150, 161–171. [Google Scholar] [CrossRef]
- Zhang, G.; Luo, L.; Tian, K.; Ning, F.; Wu, D.; Sun, Q.; Yu, W.; Yi, S.; Hao, Y. Metabolomics analysis of Dendrobium officinale tissue-cultured seedlings under red-blue composed light by using HPLC and UPLC-Q/TOF-MS. Plant Cell Tissue Organ Cult. (PCTOC) 2024, 156, 49. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth and Antioxidant Parameters | Linear Regression Model | R2 | p |
|---|---|---|---|
| Leaf length | 0.84 | 0.00 | |
| Root number | 0.83 | 0.01 | |
| Root length | 0.93 | 0.00 | |
| Root activity | 0.92 | 0.00 | |
| Fresh weight | 0.93 | 0.00 | |
| Dry weight | 0.99 | 0.00 | |
| Leaf number | 0.96 | 0.00 | |
| SSC | 0.97 | 0.00 | |
| SPC | 0.97 | 0.00 | |
| FPC | 0.85 | 0.00 | |
| SOD | 0.89 | 0.00 | |
| POD | 0.79 | 0.00 | |
| CAT | 0.86 | 0.00 | |
| APX | 0.95 | 0.00 |
| Factor | Principal Components | |||
|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC4 | |
| Dry weight | 0.986 | 0.330 | 0.111 | 0.095 |
| APX | 0.979 | 0.130 | 0.046 | 0.146 |
| ΦPSII | 0.975 | 0.006 | 0.188 | 0.079 |
| Fresh weight | 0.961 | 0.143 | −0.140 | 0.047 |
| Fv/Fm | 0.957 | 0.040 | −0.019 | −0.240 |
| Chlorophyll a + b | 0.953 | −0.132 | 0.148 | 0.098 |
| SOD | 0.948 | 0.045 | −0.269 | −0.162 |
| Leaf number | 0.937 | −0.099 | −0.122 | −0.235 |
| Chlorophyll a | 0.928 | −0.148 | 0.315 | −0.118 |
| Root activity | 0.916 | 0.067 | −0.324 | −0.108 |
| CAT | 0.899 | −0.002 | −0.438 | 0.005 |
| Chlorophgyll b | 0.889 | −0.240 | −0.162 | 0.320 |
| Fv/Fo | 0.885 | −0.095 | 0.381 | 0.106 |
| ETRll | 0.824 | 0.125 | −0.308 | −0.216 |
| qP | 0.823 | 0.513 | 0.188 | 0.156 |
| FPC | −0.803 | −0.100 | 0.088 | 0.512 |
| NPQ | 0.802 | −0.512 | 0.170 | 0.219 |
| Root number | 0.793 | −0.265 | −0.541 | −0.063 |
| SPC | 0.786 | −0.390 | 0.369 | −0.215 |
| Carotenoids | 0.781 | 0.379 | 0.221 | −0.119 |
| POD | 0.728 | 0.364 | 0.244 | 0.326 |
| Root length | 0.698 | −0.015 | 0.681 | −0.219 |
| MDA | −0.679 | −0.092 | 0.628 | 0.040 |
| Leaf length | 0.568 | −0.553 | 0.460 | 0.363 |
| Plantlet length | −0.040 | 0.934 | 0.065 | 0.285 |
| SSC | 0.442 | 0.885 | 0.083 | 0.087 |
| Chlorophgyll a/b | −0.465 | 0.252 | 0.588 | −0.605 |
| Characteristic value | 18.210 | 3.250 | 2.880 | 1.515 |
| Contribution rate | 62.128% | 13.424% | 10.652% | 5.611% |
| Cumulative contribution rate | 91.815% | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huo, J.; Lin, Q.; Mo, L.; Zheng, L.; Meng, X.; Song, X.; Liang, J.; Chen, T. The Influence of Varying Wavelengths of LED Light on the Development, Physiology Response, and Metabolism Activities of Micropropagated Dendrobium Hybrid ‘Shuijing’ Plantlets. Horticulturae 2024, 10, 774. https://doi.org/10.3390/horticulturae10080774
Huo J, Lin Q, Mo L, Zheng L, Meng X, Song X, Liang J, Chen T. The Influence of Varying Wavelengths of LED Light on the Development, Physiology Response, and Metabolism Activities of Micropropagated Dendrobium Hybrid ‘Shuijing’ Plantlets. Horticulturae. 2024; 10(8):774. https://doi.org/10.3390/horticulturae10080774
Chicago/Turabian StyleHuo, Jiaran, Qiongyu Lin, Liwen Mo, Lifang Zheng, Xinya Meng, Xiqiang Song, Jinfeng Liang, and Tiangang Chen. 2024. "The Influence of Varying Wavelengths of LED Light on the Development, Physiology Response, and Metabolism Activities of Micropropagated Dendrobium Hybrid ‘Shuijing’ Plantlets" Horticulturae 10, no. 8: 774. https://doi.org/10.3390/horticulturae10080774
APA StyleHuo, J., Lin, Q., Mo, L., Zheng, L., Meng, X., Song, X., Liang, J., & Chen, T. (2024). The Influence of Varying Wavelengths of LED Light on the Development, Physiology Response, and Metabolism Activities of Micropropagated Dendrobium Hybrid ‘Shuijing’ Plantlets. Horticulturae, 10(8), 774. https://doi.org/10.3390/horticulturae10080774