

Modeling the Performance Parameters of Pollen Grains of Male Date Palms Using an Artificial Neural Network Based on the Mineral Composition and Morphological Properties of Their Leaves

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. The Experimental Location
2.2. Morphological Features of Male Date Palm Leaves
2.3. Pollen Grains’ Viability
2.4. Determination of Nutritional Status of the Male Trees’ Leaves
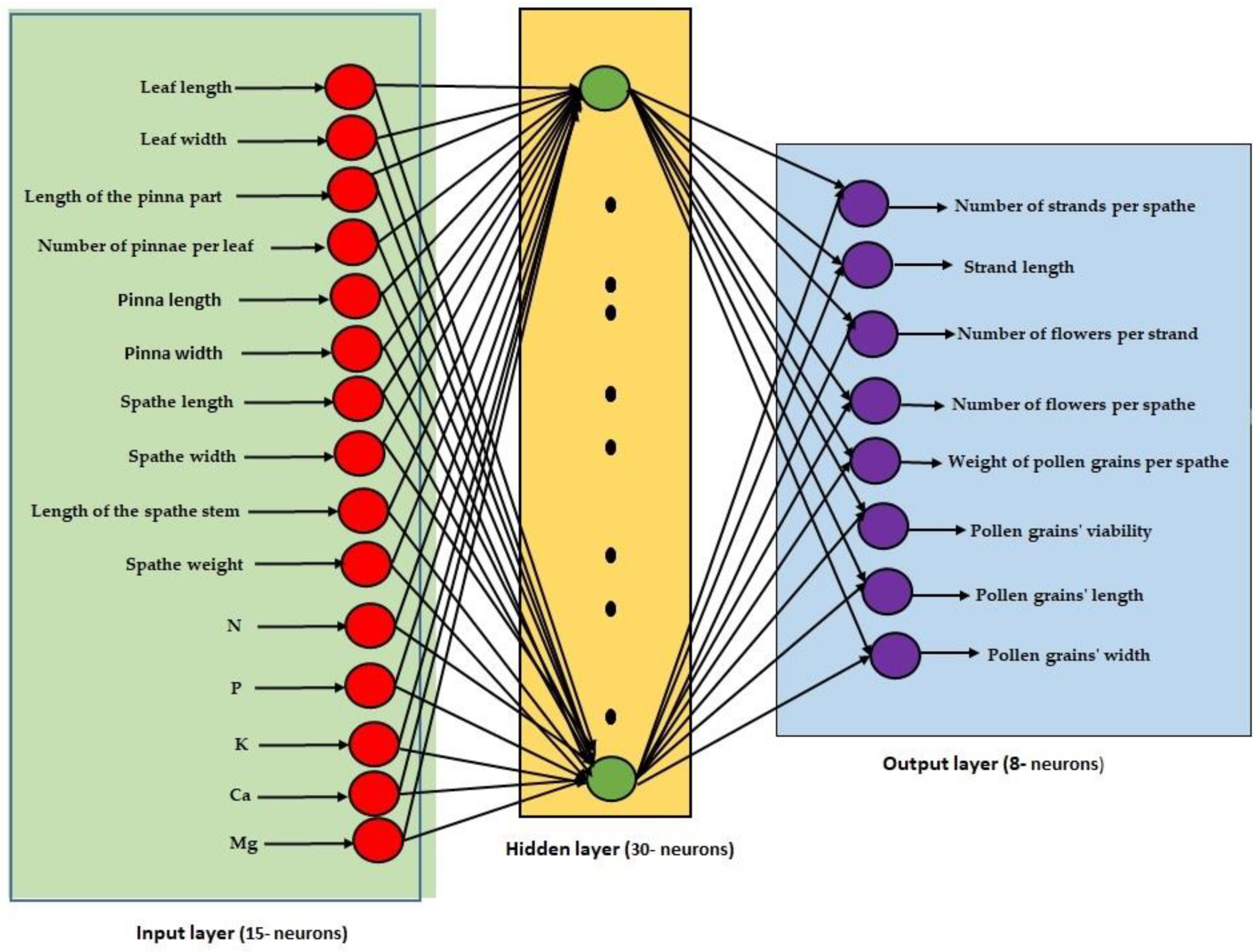
2.5. Structure of the Artificial Neural Network Model
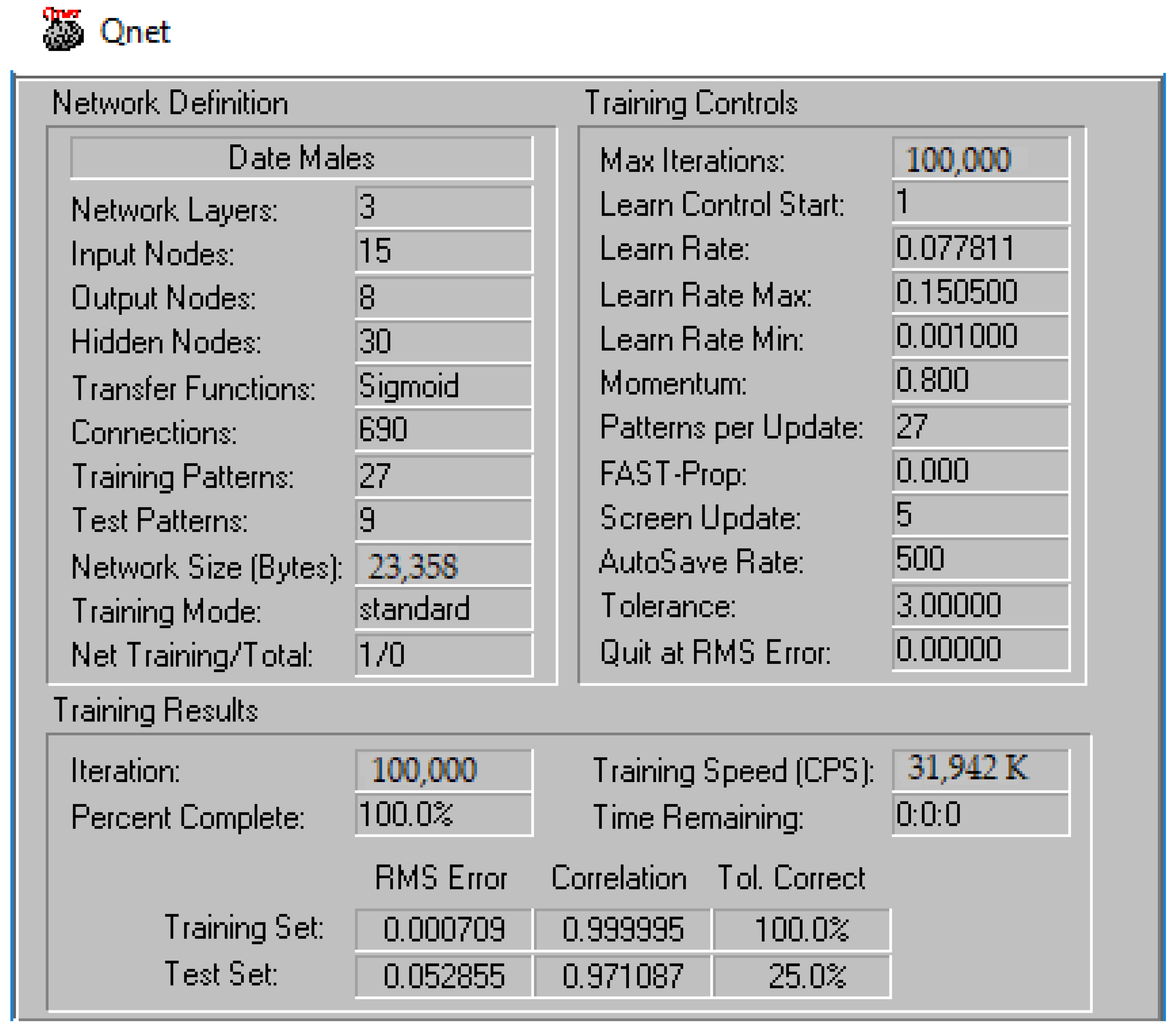
2.6. Evaluation of the ANN Model
2.7. Sensitivity Analysis
3. Results and Discussion
3.1. Statistical Analysis of the Features Used as Input Parameters in the Newly Developed ANN Model
3.2. Distribution of the Investigated Performance Parameters of Pollen Grains
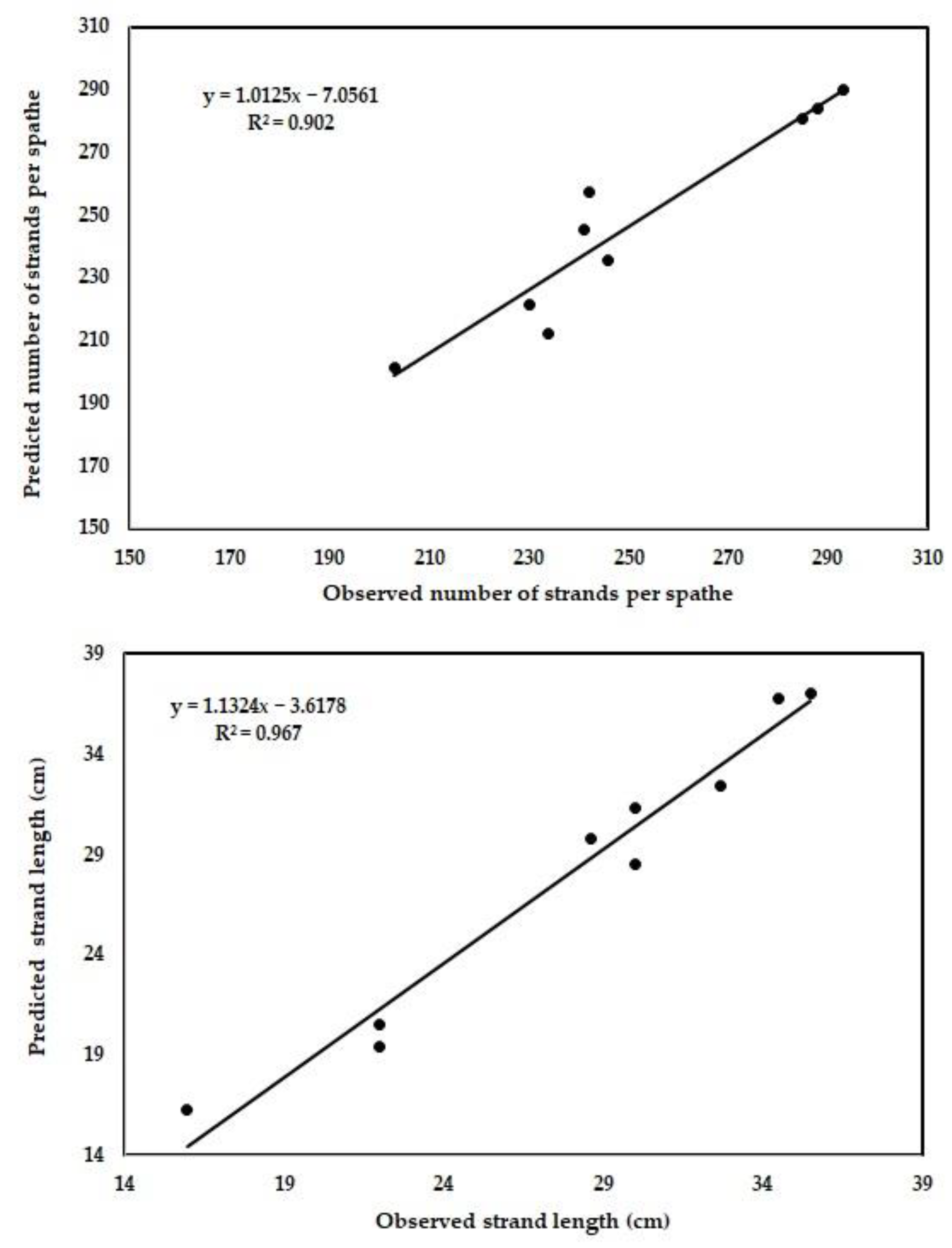
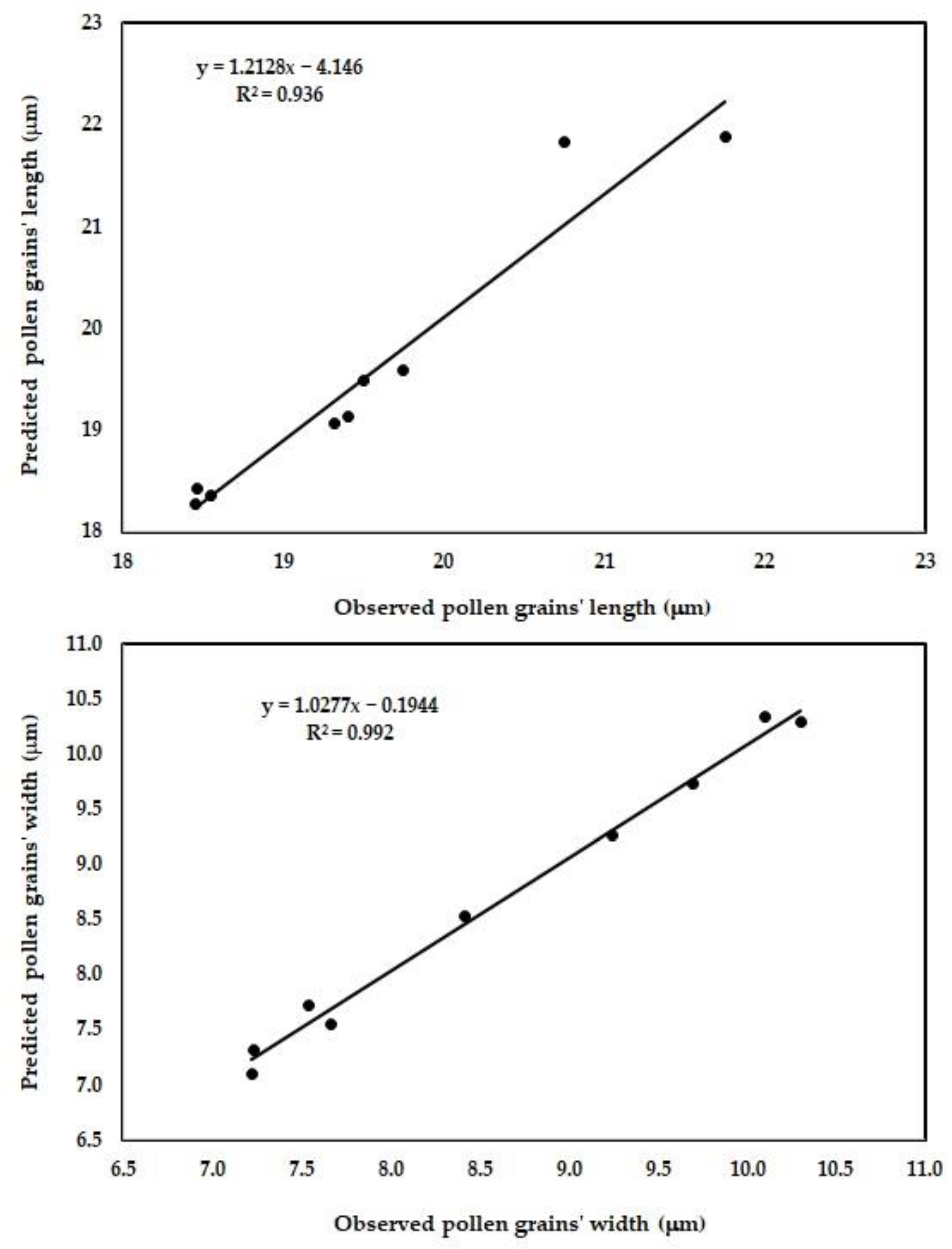
3.3. Performance of the Newly Developed ANN Model
3.4. Outcome of the Sensitivity Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Al-Abbad, A.; Al-Jamal, M.; Al-Elaiw, Z.; Al-Shreed, F.; Belaif, H. A study on the economic feasibility of date palm cultivation in the Al-Hassa Oasis of Saudi Arabia. Afr. J. Agric. Mark. 2018, 6, 1–6. [Google Scholar]
- Elfeky, A.; Elfaki, J.A. Review: Date Palm Irrigation Methods and Water Resources in the Kingdom of Saudi Arabia. J. Eng. Res. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Alkhaldi, G.; Emam, A.; Mosbah, E. An Economic Study: Date Palm Farmers in Al Ahsa Oasis-Kingdom of Saudi Arabia. Univers. J. Agric. Res. 2022, 10, 794–803. [Google Scholar] [CrossRef]
- General Authority of Statestics. Agricultural Production Survey Bulletin 2019, Kingdom of Saudi Arabia. Available online: https://www.stats.gov.sa/sites/default/files/Agriculture%20Production%20Survey%202019%20EN.pdf (accessed on 8 May 2024).
- Baloch, M.K.; Saleem, S.A.; Ahmad, K.; Baloch, A.K.; Baloch, W.A. Impact of controlled atmosphere on the stability of Dhakki dates. Swiss Soc. Food Sci. Tech. 2006, 39, 671–676. [Google Scholar] [CrossRef]
- Al-Farsi, M.; Alasalvar, C.; Morris, A.; Barron, M.; Shahidi, F. Compositional and sensory characteristics of three native sundried date (Phoenix dactylifera L.) varieties grown in Oman. J. Agric. Food Chem. 2005, 53, 7586–7591. [Google Scholar] [CrossRef]
- Mohammed, M.; Munir, M.; Aljabr, A. Prediction of Date Fruit Quality Attributes during Cold Storage Based on Their Electrical Properties Using Artificial Neural Networks Models. Foods 2022, 11, 1666. [Google Scholar] [CrossRef]
- Hachef, A.; Bourguiba, H.; Cherif, E.; Ivorra, S.; Terral, J.-F.; Zehdi-Azouzi, S. Agro-morphological traits assessment of Tunisian male date palms (Phoenix dactylifera L.) for preservation and sustainable utilization of local germplasm. Saudi J. Biol. Sci. 2023, 30, 103574. [Google Scholar] [CrossRef]
- Halbritter, H.; Ulrich, S.; Grímsson, F.; Weber, M.; Zetter, R.; Hesse, M.; Buchner, R.; Svojtka, M.; Frosch-Radivo, A. Illustrated Pollen Terminology; Springer International Publishing: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Soliman, S.S.; Al-Obeed, R.S. Investigations on the pollen morphology of some date palm males (Phoenix dactylifera L.) in Saudi Arabia. Aust. J. Crop Sci. 2013, 7, 1355–1360. [Google Scholar]
- Al-Khalifah, N.S. Metaxenia: Influence of pollen on the maternal tissue of fruits of two cultivars of date palm (Phoenix dactylifera L.). Bangladesh J. Bot. 2006, 35, 151–161. [Google Scholar]
- Rizk, R.M.; El-Sharabasy, S.F.; Soliman, K.A. Characterization and evaluation of sex males date palm (Phoenix dactylifera L.) genotypes in Egypt. In Proceedings of the Fourth Symposium on the Date Palm in Saudi Arabia, Al-Hassa, Saudi Arabia, 20–22 February 2007; p. 238. [Google Scholar]
- Sajindra, H.; Abekoon, T.; Wimalasiri, E.M.; Mehta, D.; Rathnayake, U. An Artificial Neural Network for Predicting Groundnut Yield Using Climatic Data. AgriEngineering 2023, 5, 1713–1736. [Google Scholar] [CrossRef]
- Dahikar, S.S.; Rode, S.V. Agricultural Crop Yield Prediction Using Artificial Neural Network Approach. Int. J. Innov. Res. Electr. Electron. Instrum. Control. Eng. 2014, 2, 683–686. [Google Scholar]
- Aubakirova, G.; Ivel, V.; Gerassimova, Y.; Moldakhmetov, S.; Petrov, P. Application of artificial neural network for wheat yield forecasting. East. -Eur. J. Enterp. Technol. 2022, 3, 31–39. [Google Scholar] [CrossRef]
- Chlingaryan, A.; Sukkarieh, S.; Whelan, B. Machine learning approaches for crop yield prediction and nitrogen status estimation in precision agriculture: A review. Comput. Electron. Agric. 2018, 151, 61–69. [Google Scholar] [CrossRef]
- Zaid, A.; Arias-Jimenez, E.J. Date Palm Cultivation; Food and Agricultural Organization: Rome, Italy, 2002. [Google Scholar]
- Shawky, B.A.; El-Sharabasy, S.F. Date Palm Status and Perspective in Egypt. In Date Palm Genetic Resources and Utilization: Africa and the Americas, 1st ed.; Al-Khayri, J.M., Mohan, S., Johnson, D.V., Eds.; Springer: New York, NY, USA, 2015; Volume 1, pp. 75–124. [Google Scholar]
- Munir, M. An appraisal of pollen germination and viability of varied male pollen sources of date palm (Phoenix dactylifera L.). J. Appl. Hortic. 2021, 23, 254–259. [Google Scholar] [CrossRef]
- Elboghdady, A.E.A.; Gomma, A.H.; Hamed, A.M.; Abdallatif, A.M. Assessment of phenotypic diversity of some date palm male genotypes growing under Egyptian conditions. Rev. Bras. Frutic. 2023, 45, e-896. [Google Scholar] [CrossRef]
- Shafique, M.; Khan, A.S.; Malik, A.U.; Shahid, M.; Rajwana, I.A.; Saleem, B.A.; Amin, M.; Ahmad, I. Influence of pollen source and pollination frequency on fruit drop, yield and quality of date palm (Phoenix dactylifera L.) cv. Dhakki. Pak. J. Bot. 2011, 43, 831–839. [Google Scholar]
- Iqbal, M.; Niamatullah, M.; Munir, M. Effect of various Dactylifera males pollinizer on pomological traits and economical yield index of cv’s Shakri, Zahidi and Dhakki date palm (Phoenix dactylifera L.). J. Anim. Plant Sci. 2012, 22, 376–383. [Google Scholar]
- Rezazadeh, R.; Hassanzadeh, H.; Hosseini, Y.; Karami, Y.; Williams, R.R. Influence of pollen source on fruit production of date palm (Phoenix dactylifera L.) cv. Barhi in humid coastal regions of southern Iran. Sci. Hortic-Amst. 2013, 160, 182–188. [Google Scholar] [CrossRef]
- Hafez, O.M.; Saleh, M.A.; Mostafa, E.A.M.; Naguib, M.M.; Ashour, N.E. Effect of pollen grain sources on yield and fruit quality of Samany date palm. Int. J. Agric. Res. 2014, 9, 164–168. [Google Scholar] [CrossRef]
- Benamor, B.; Boughediri, L.; Chala, A. Selection of male date palms (Phoenix dactylifera L.) at “Daouia” station (Oued Souf, Algeria). Adv. Environ. Biol. 2014, 8, 29–36. [Google Scholar]
- Mohammadi, K. Groundwater Table Estimation Using MODFLOW and Artificial Neural Networks. In Practical Hydroinformatics; Robert, J., Abrahart Linda, M., See, D.P.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 127–138. [Google Scholar]
- Basheer, I.A.; Hajmeer, M. Artificial neural networks: Fundamentals, computing, design, and application. J. Microbiol. Methods 2000, 43, 3–31. [Google Scholar] [CrossRef] [PubMed]
- Salem, A.O.M.; Rhouma, S.; Zehdi, S.; Marrakchi, M.; Trifi, M. Morphological variability of Mauritanian date-palm (Phoenix dactylifera L.) cultivars as revealed by vegetative traits. Acta Bot. Croat. 2008, 67, 81–90. [Google Scholar]
- Shaheen, M. Identification of some seedling male date palms by pollen ultrastructure. J. Coll. Agric. King Saud Univ. 1983, 5, 137–142. [Google Scholar]
- Javady, T.; Arzani, K. Pollen morphology of five Iranian olive (Olea europaea L.) cultivars. J. Agric. Sci. Technol. 2011, 3, 37–42. [Google Scholar]
- Pearson, H.M.; Harey, P.M. Pollen Viability in Rosa. Hort Sci. 1984, 19, 710–771. [Google Scholar] [CrossRef]
- Rizk, S.A.Y. Studied on the Effect of Fertilization on Date Palm Production. Ph.D. Thesis, Ain Shams University, Cairo, Egypt, 1987. [Google Scholar]
- Evenhuis, B.; De Waard, P.W. Principles and practices in plant analysis. FAO Soils Bull. 1980, 38, 152–163. [Google Scholar]
- Chapman, H.D.; Pratt, P.F. Methods of analysis for soils, plants and waters. Soil Sci. 1962, 93, 68. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphorus in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Evenhuis, B. Nitrogen Determination; Department of Agricultural Research Royal Tropical Institute: Amsterdam, The Netherlands, 1976. [Google Scholar]
- Silva, F.A.N.; Delgado, J.M.P.Q.; Cavalcanti, R.S.; Azevedo, A.C.; Guimarães, A.S.; Lima, A.G.B. Use of nondestructive testing of ultrasound and artificial neural networks to estimate compressive strength of concrete. Buildings 2021, 11, 44. [Google Scholar] [CrossRef]
- Bakas, I.; Kontoleon, K.J. Performance Evaluation of Artificial Neural Networks (ANN) Predicting Heat Transfer through Masonry Walls Exposed to Fire. Appl. Sci. 2021, 11, 11435. [Google Scholar] [CrossRef]
- Al-Sager, S.M.; Almady, S.S.; Marey, S.A.; Al-Hamed, S.A.; Aboukarima, A.M. Prediction of Specific Fuel Consumption of a Tractor during the Tillage Process Using an Artificial Neural Network Method. Agronomy 2024, 14, 492. [Google Scholar] [CrossRef]
- Wu, W.; Dandy, G.C.; Maier, H. Protocol for developing ANN models and its application to the assessment of the quality of the ANN model development process in drinking water quality modelling. Environ. Model. Softw. 2014, 54, 108–127. [Google Scholar] [CrossRef]
- Jørgensen, C.; Grastveit, R.; Garzón-Roca, J.; Payá-Zaforteza, I.; Adam, J.M. Bearing capacity of steel-caged RC columns under combined bending and axial loads: Estimation based on Artificial Neural Networks. Eng. Struct. 2013, 56, 1262–1270. [Google Scholar] [CrossRef]
- Naser, M.Z. Mechanistically Informed Machine Learning and Artificial Intelligence in Fire Engineering and Sciences. Fire Technol. 2021, 57, 2741–2784. [Google Scholar] [CrossRef]
- Nov, P.; Peansupap, V. Using artificial neural network for selecting type of subcontractor relationships in construction projec. Eng. J. 2020, 24, 73–88. [Google Scholar] [CrossRef]
- Cybenko, G. Approximation by superpositions of a sigmoidal function. Math. Control Signals Syst. 1989, 2, 303–314. [Google Scholar] [CrossRef]
- Tsae, N.B.; Adachi, T.; Kawamura, Y. Application of artificial neural network for the prediction of copper ore grade. Minerals 2023, 13, 658. [Google Scholar] [CrossRef]
- Sammen, S.S.; Kisi, O.; Ehteram, M.; El-Shafie, A.; Al-Ansari, N.; Ghorbani, M.A.; Bhat, S.A.; Ahmed, A.N.; Shahid, S. Rainfall modeling using two different neural networks improved by metaheuristic algorithms. Environ. Sci. Eur. 2023, 35, 112. [Google Scholar] [CrossRef]
- Green, P.E.; Carroll, J.D. Mathematical Tools for Applied Multivariate Analysis. Academic Press: New York, NY, USA, 1978. [Google Scholar]
- Elhoumaizi, M.A.; Saaidi, M.; Oihabi, A.; Cilas, C. Phenotypic diversity of date palm cultivars (Phoenix dactylifera L.) from Morocco. Genet. Resour. Crop Evol. 2002, 49, 483–490. [Google Scholar] [CrossRef]
- Shaheen, M. Evaluation of date palm males using pollen viability and ultrastructure. Acta Hortic. 2004, 632, 37–43. [Google Scholar] [CrossRef]
- Kassem, H.A. The response of date palm to calcareous soil fertilization. J. Soil Sci. Plant Nutr. 2012, 12, 45–58. [Google Scholar] [CrossRef]
- Ibrahim, M.M.; El-Beshbeshy, R.T.; Kamh, N.R.; Abou-Amer, A.I. Effect of NPK and biofertilizer on date palm trees grown in Siwa Oasis, Egypt. Oil Use Manag. 2013, 29, 315–321. [Google Scholar] [CrossRef]
- Soliman, S.S.; Al-Obeed, R.S.; Omar, A.A.; Ahmed, M.A. A Comparative study of the morphological characteristics of some seedling date palm males. J. Appl. Sci. Res. 2013, 9, 4463–4468. [Google Scholar]
- Nesiem, M.R.A.; Darwish, E.; Gadalla, E.G.; Shaimaa, F.A. Comparison study among pollen grains of three date palm males cultivars (Phoenix dactylifera L.). J. Biol. Chem. Environ. Sci. 2016, 11, 263–275. [Google Scholar]
- El-Salhy, A.M.; Abdalla, A.Y.; Mostafa, R.A.A. Evaluation of some date palm male seedlings in pollination of Zaghloul and Samany date palms under Assiut conditions. Assiut J. Agric. Sci. 1997, 28, 79–89. [Google Scholar]
- Islam, S.E. Evaluation of Date Palm Males Types as Pollinators for Zaghloul and Samany Date Palm cvs. Grown in Qalyubia Governorate. Middle East J. Agric. Res. 2017, 6, 1049–1056. [Google Scholar]
- Djerouni, A.; Chala, A.; Simozrag, A.A.; Benmehaia, R.; Baka, M. Evaluation of male palms used in pollination and the extent of its relationship with cultivars of date-palms (Phoenix dactylifera L.) grown in region of oued right, Algeria. Pak. J. Bot. 2015, 47, 2295–2300. [Google Scholar]
- Ibrahim, M.M.; Ghonimy, M.; Abd El Rahman, E. Physical and aerodynamic properties of date palm pollen grains. Acta Agric. Slov. 2023, 119, 1–11. [Google Scholar] [CrossRef]
- Hessane, A.; EL Youssefi, A.; Farhaoui, Y.; Aghoutane, B.; Qaraai, Y. Artificial Intelligence Applications in Date Palm Cultivation and Production: A Scoping Review. In Artificial Intelligence and Smart Environment, ICAISE 2022; Farhaoui, Y., Rocha, A., Brahmia, Z., Bhushab, B., Eds.; Lecture Notes in Networks and Systems 2023; Springer: Cham, Switzerland, 2023; Volume 635. [Google Scholar] [CrossRef]
- Ismail, A.I.H.; Hassaballa, A.A.; Almadini, A.M.; Daffalla, S. Analyzing the Spatial Correspondence between Different Date Fruit Cultivars and Farms’ Cultivated Areas, Case Study: Al-Ahsa Oasis, Kingdom of Saudi Arabia. Appl. Sci. 2022, 12, 5728. [Google Scholar] [CrossRef]
- Husain, M.; Khan, M.Z. Performance Analysis of Dates Production in Madinah Region (KSA) Using Ensemble Learning. Int. J. Comput. Sci. Netw. Secur. 2019, 19, 31–36. [Google Scholar]
- Naqvi, S.A.; Khan, I.A.; Pintaud, J.C.; Jaskani, M.J.; Ali, A. Morphological characterization of Pakistani date palm (Phoenix dactylifera L.) genotypes. Pak. J. Agri. Sci. 2015, 52, 645–650. [Google Scholar]
- Aly, H.S.H. Evaluation of pollen grains germination, viability and chemical composition of some date palm males. Middle East J. Agric. Res. 2018, 7, 235–247. [Google Scholar]
- Abd, M.A.; Taain, D.A.; Al-Thahab, E.A.M. Morphological study (vegetative and floral) of twelve date palm male cultivars. Plant Arch. 2019, 19, 1349–1357. [Google Scholar]
- Elmeer, K.; Ahmed, A.; Serqiwa, S. Morphological diversity of three elite date palm (Phoenix dactylifera L.) cultivars grown in South of Libya. In Proceedings of the Fourth Scientific Conference of Environment and Sustainable Development in the Arid and Semi-Arid Regions, Ejdabya, Libya, 20–22 November 2016; pp. 85–92. [Google Scholar]
- Mohammed, M.; Alqahtani, N.K.; Munir, M.; Eltawil, M.A. Applications of AI and IoT for Advancing Date Palm Cultivation in Saudi Arabia. In Internet Things New Insights; Habib, M.K., Ed.; IntechOpen: London, UK, 2024; 256p, Chapter 3; ISBN 978-1-83768-989-7. [Google Scholar] [CrossRef]
- Mohammed, M.; El-Shafie, H.; Munir, M. Development and Validation of Innovative Machine Learning Models for Predicting Date Palm Mite Infestation on Fruits. Agronomy 2023, 13, 494. [Google Scholar] [CrossRef]
- Dehghanisanij, H.; Salamati, N.; Emami, S.; Emami, H.; Fujimaki, H. An intelligent approach to improve date palm crop yield and water productivity under different irrigation and climate scenarios. Appl. Water Sci. 2023, 13, 56. [Google Scholar] [CrossRef]
- Hobani, A.I.; Thottam, A.M.; Ahmed, K.A. Development of a neural network classifier for date fruit varieties using some physical attributes. King Saud Univ. Agric. Res. Cent. 2003, 126, 5–18. [Google Scholar]
- Khriji, L.; Ammari, A.C.; Awadalla, M. Artificial Intelligent Techniques for Palm Date Varieties Classification. Int. J. Adv. Comput. Sci. Appl. 2020, 11, 489–495. [Google Scholar] [CrossRef]
- Rybacki, P.; Niemann, J.; Derouiche, S.; Chetehouna, S.; Boulaares, I.; Seghir, N.M.; Diatta, J.; Osuch, A. Convolutional Neural Network (CNN) Model for the Classification of Varieties of Date Palm Fruits (Phoenix dactylifera L.). Sensors 2024, 24, 558. [Google Scholar] [CrossRef]
- Koklu, M.; Kursun, R.; Taspinar, Y.S.; Cinar, I. Classification of Date Fruits into Genetic Varieties Using Image Analysis. Math. Probl. Eng. 2021, 2021, 4793293. [Google Scholar] [CrossRef]
- Shoukry, N. Artificial Neural Networks Based Change Detection for Monitoring Palm Trees Plantation in Al Madinah-Saudi Arabia. Bull. Soc. Geo. Egypt 2017, 90, 167–200. [Google Scholar] [CrossRef]
- Eskandari, M.; Zeinadini, A.; Seyedmohammadi, J.; Navidi, M. Estimating Quantity of Date Yield Using Soil Properties by Regression and Artificial Neural Network. Commun. Soil Sci. Plant Anal. 2023, 54, 36–47. [Google Scholar] [CrossRef]
- Schalkoff, R.J. Artificial Neural Networks; McGraw-Hill Higher Education: New York, NY, USA, 1997; ISBN 007057118X. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Male Date Palms | Leaf Length | Leaf Width | Length of Pinna Part | Number of Pinnae per Leaf | Pinna Length | Pinna Width | Spathe Length | Spathe Width | Length of Spathe Stem | Spathe Weight |
|---|---|---|---|---|---|---|---|---|---|---|
| (m) | (m) | (m) | (-) | (cm) | (cm) | (cm) | (cm) | (cm) | (kg) | |
| DPM1 | 4.37 ± 0.04 | 0.35 ± 0.01 | 3.08 ± 0.02 | 204 ± 2.65 | 48.00 ± 3.65 | 3.43 ± 0.06 | 153.75 ± 0.96 | 18.13 ± 0.32 | 76.50 ± 1.29 | 4.10 ± 0.14 |
| DPM2 | 3.68 ± 0.05 | 0.72 ± 0.02 | 2.78 ± 0.02 | 176 ± 1.83 | 43.98 ± 0.21 | 3.45 ± 0.02 | 123.56 ± 0.66 | 15.63 ± 0.32 | 51.25 ± 0.96 | 2.95 ± 0.13 |
| DPM3 | 3.97 ± 0.10 | 0.49 ± 0.01 | 3.07 ± 0.02 | 178 ± 1.26 | 58.50 ± 0.74 | 4.17 ± 0.02 | 73.00 ± 2.16 | 14.75 ± 0.46 | 26.00 ± 0.82 | 2.04 ± 0.08 |
| DPM4 | 4.14 ± 0.05 | 0.60 ± 0.01 | 3.04 ± 0.03 | 206 ± 3.65 | 44.25 ± 1.71 | 3.51 ± 0.06 | 94.00 ± 1.83 | 20.41 ± 0.17 | 52.50 ± 1.29 | 3.09 ± 0.10 |
| DPM5 | 3.86 ± 0.03 | 0.59 ± 0.01 | 3.13 ± 0.02 | 188 ± 0.96 | 52.33 ± 0.06 | 3.52 ± 0.07 | 70.50 ± 1.29 | 17.64 ± 0.11 | 21.50 ± 0.54 | 2.23 ± 0.06 |
| DPM6 | 3.57 ± 0.02 | 0.59 ± 0.01 | 2.62 ± 0.07 | 196 ± 0.82 | 47.44 ± 0.13 | 3.74 ± 0.05 | 117.89 ± 1.16 | 18.69 ± 0.13 | 47.45 ± 0.09 | 3.70 ± 0.04 |
| DPM7 | 3.67 ± 0.04 | 0.47 ± 0.01 | 2.87 ± 0.02 | 210 ± 2.75 | 50.60 ± 0.29 | 3.86 ± 0.04 | 60.50 ± 1.29 | 17.68 ± 0.06 | 23.50 ± 0.58 | 1.65 ± 0.02 |
| DPM8 | 3.84 ± 0.02 | 0.87 ± 0.02 | 2.75 ± 0.03 | 204 ± 1.29 | 47.95 ± 0.14 | 3.93 ± 0.03 | 72.00 ± 0.82 | 18.00 ± 0.09 | 16.66 ± 0.09 | 0.78 ± 0.02 |
| DPM9 | 4.45 ± 0.03 | 0.49 ± 0.01 | 2.61 ± 0.03 | 202 ± 2.94 | 49.72 ± 0.30 | 3.77 ± 0.02 | 97.28 ± 0.55 | 13.02 ± 0.07 | 38.28 ± 0.22 | 1.58 ± 0.05 |
| Male Date Palms | N | P | K | Ca | Mg |
|---|---|---|---|---|---|
| (%) | (%) | (%) | (%) | (%) | |
| DPM1 | 0.98 ± 0.02 | 0.04 ± 0.001 | 1.01 ± 0.02 | 0.54 ± 0.01 | 0.15 ± 0.01 |
| DPM2 | 0.94 ± 0.02 | 0.04 ± 0.001 | 0.81 ± 0.02 | 0.55 ± 0.01 | 0.14 ± 0.01 |
| DPM3 | 0.86 ± 0.02 | 0.04 ± 0.002 | 1.25 ± 0.01 | 0.54 ± 0.01 | 0.15 ± 0.01 |
| DPM4 | 0.96 ± 0.01 | 0.04 ± 0.0012 | 1.27 ± 0.02 | 0.46 ± 0.01 | 0.16 ± 0.01 |
| DPM5 | 1.03 ± 0.04 | 0.04 ± 0.001 | 1.21 ± 0.01 | 0.47 ± 0.02 | 0.12 ± 0.01 |
| DPM6 | 1.12 ± 0.04 | 0.06 ± 0.002 | 1.09 ± 0.03 | 0.43 ± 0.01 | 0.16 ± 0.01 |
| DPM7 | 0.91 ± 0.03 | 0.05 ± 0.001 | 1.20 ± 0.02 | 0.49 ± 0.02 | 0.18 ± 0.01 |
| DPM8 | 0.94 ± 0.02 | 0.05 ± 0.001 | 1.34 ± 0.01 | 0.35 ± 0.01 | 0.11 ± 0.01 |
| DPM9 | 0.85 ± 0.01 | 0.05 ± 0.002 | 1.00 ± 0.02 | 0.52 ± 0.01 | 0.16 ± 0.01 |
| Male Date Palms | Number of Strands per Spathe | Strand Length | Number of Flowers per Strand | Number of Flowers per Spathe | Weight of Pollen Grains per Spathe | Pollen Grains’ Viability | Pollen Grains’ Length | Pollen Grains’ Width |
|---|---|---|---|---|---|---|---|---|
| (-) | (cm) | (-) | (-) | (g) | (%) | (μm) | (μm) | |
| DPM1 | 234 ± 2.63 | 35.31 ± 0.90 | 57 ± 1.29 | 13,205 ± 195.31 | 27.19 ± 0.69 | 84.25 ± 1.71 | 21.50 ± 0.54 | 10.23 ± 0.10 |
| DPM2 | 286 ± 2.38 | 31.50 ± 1.29 | 62 ± 1.29 | 17,556 ± 227.90 | 18.78 ± 0.51 | 94.25 ± 1.26 | 19.51 ± 0.12 | 7.70 ± 0.12 |
| DPM3 | 294 ± 1.29 | 30.50 ± 1.29 | 65 ± 0.96 | 19,152 ± 343.61 | 17.43 ± 0.06 | 85.50 ± 1.29 | 19.63 ± 0.10 | 8.45 ± 0.05 |
| DPM4 | 246 ± 3.50 | 28.64 ± 0.11 | 50 ± 1.29 | 12,188 ± 309.93 | 19.45 ± 0.07 | 80.00 ± 1.83 | 19.38 ± 0.19 | 9.58 ± 0.10 |
| DPM5 | 222 ± 2.89 | 26.78 ± 0.10 | 60 ± 1.29 | 13,178 ± 276.05 | 15.14 ± 0.05 | 88.25 ± 0.96 | 17.20 ± 0.08 | 6.94 ± 0.06 |
| DPM6 | 284 ± 1.71 | 32.20 ± 0.37 | 51 ± 1.29 | 14,353 ± 298.46 | 15.65 ± 0.05 | 92.50 ± 1.29 | 19.34 ± 0.06 | 9.25 ± 0.02 |
| DPM7 | 243 ± 2.65 | 21.50 ± 0.58 | 90 ± 2.16 | 21,823 ± 476.13 | 17.44 ± 0.02 | 83.50 ± 1.29 | 18.44 ± 0.09 | 7.21 ± 0.02 |
| DPM8 | 201 ± 3.51 | 15.94 ± 0.02 | 59 ± 0.82 | 11,830 ± 276.74 | 14.75 ± 0.04 | 92.50 ± 1.29 | 18.49 ± 0.06 | 7.64 ± 0.02 |
| DPM9 | 151 ± 3.16 | 27.62 ± 0.15 | 57 ± 0.96 | 8643 ± 88.29 | 29.01 ± 0.17 | 79.75 ± 0.96 | 20.27 ± 0.04 | 8.08 ± 0.10 |
| Output Nodes | Standard Deviation | Bias | Maximum Error |
|---|---|---|---|
| Number of strands per spathe | 0.19 | −0.0021 | 0.55 |
| Strand length (cm) | 0.01 | −4.09 × 10−5 | 0.04 |
| Number of flowers per strand | 0.02 | 0.0006 | 0.06 |
| Number of flowers per spathe | 12.33 | −0.11 | 43.85 |
| Weight of pollen grains per spathe (g) | 0.03 | −7.74 × 10−6 | 0.07 |
| Pollen grains’ viability (%) | 0.006 | 0.0002 | 0.015 |
| Pollen grains’ length (μm) | 0.006 | 3.95 × 10−5 | 0.019 |
| Pollen grains’ width (μm) | 0.002 | 2.80 × 10−5 | 0.003 |
| Output Nodes | Standard Deviation | Bias | Maximum Error |
|---|---|---|---|
| Number of strands per spathe | 10.42 | −3.92 | 22.09 |
| Strand length (cm) | 1.54 | 0.08 | 2.60 |
| Number of flowers per strand | 3.14 | −1.07 | 5.57 |
| Number of flowers per spathe | 977.61 | −360.97 | 1946.16 |
| Weight of pollen grains per spathe (g) | 0.88 | 0.18 | 1.74 |
| Pollen grains’ viability (%) | 2.10 | −0.06 | 3.52 |
| Pollen grains’ length (μm) | 0.40 | 0.013 | 1.094 |
| Pollen grains’ width (μm) | 0.12 | 0.04 | 0.23 |
| Output Nodes | Training Dataset | Testing Dataset | ||||
|---|---|---|---|---|---|---|
| RMSE | MAE | R2 | RMSE | MAE | R2 | |
| Number of strands per spathe | 0.19 | 2.88 | 1.000 | 10.42 | 2.88 | 0.902 |
| Strand length (cm) | 0.01 | 1.16 | 0.999 | 1.549 | 1.16 | 0.967 |
| Number of flowers per strand | 0.02 | 1.67 | 0.999 | 3.143 | 1.67 | 0.963 |
| Number of flowers per spathe | 12.33 | 28.03 | 0.999 | 977.61 | 28.03 | 0.941 |
| Weight of pollen grains per spathe (g) | 0.03 | 0.80 | 0.999 | 0.88 | 0.80 | 0.985 |
| Pollen grains’ viability (%) | 0.006 | 1.32 | 0.999 | 2.10 | 1.32 | 0.810 |
| Pollen grains’ length (μm) | 0.006 | 0.51 | 0.999 | 0.40 | 0.51 | 0.936 |
| Pollen grains’ width (μm) | 0.002 | 0.32 | 0.999 | 0.12 | 0.32 | 0.992 |
| Output Node’s Number | Output Node’s Name | Input Node’s Number | Input Node’s Name | Percentage Contributed |
|---|---|---|---|---|
| 1 | Number of strands per spathe | 1 | Leaf length | 10.96 |
| 2 | Leaf width | 2.77 | ||
| 3 | Length of the pinna part | 4.93 | ||
| 4 | Number of pinnae per leaf | 9.61 | ||
| 5 | Pinna length | 7.16 | ||
| 6 | Pinna width | 12.5 | ||
| 7 | Spathe length | 4.52 | ||
| 8 | Spathe width | 8.72 | ||
| 9 | Length of spathe stem | 17.66 | ||
| 10 | Spathe weight | 6.66 | ||
| 11 | N | 3.16 | ||
| 12 | P | 4.16 | ||
| 13 | K | 1.80 | ||
| 14 | Ca | 2.47 | ||
| 15 | Mg | 2.93 | ||
| 2 | Strand length | 1 | Leaf length | 4.50 |
| 2 | Leaf width | 4.89 | ||
| 3 | Length of the pinna part | 7.65 | ||
| 4 | Number of pinnae per leaf | 5.90 | ||
| 5 | Pinna length | 2.03 | ||
| 6 | Pinna width | 6.21 | ||
| 7 | Spathe length | 4.92 | ||
| 8 | Spathe width | 4.35 | ||
| 9 | Length of spathe stem | 4.68 | ||
| 10 | Spathe weight | 26.29 | ||
| 11 | N | 6.38 | ||
| 12 | P | 4.71 | ||
| 13 | K | 2.83 | ||
| 14 | Ca | 10.67 | ||
| 15 | Mg | 3.98 | ||
| 3 | Number of flowers per strand | 1 | Leaf length | 8.28 |
| 2 | Leaf width | 13.18 | ||
| 3 | Length of the pinna part | 3.33 | ||
| 4 | Number of pinnae per leaf | 10.69 | ||
| 5 | Pinna length | 3.82 | ||
| 6 | Pinna width | 1.39 | ||
| 7 | Spathe length | 11.85 | ||
| 8 | Spathe width | 2.59 | ||
| 9 | Length of spathe stem | 10.45 | ||
| 10 | Spathe weight | 6.62 | ||
| 11 | N | 4.98 | ||
| 12 | P | 2.54 | ||
| 13 | K | 13.98 | ||
| 14 | Ca | 3.14 | ||
| 15 | Mg | 3.17 | ||
| 4 | Number of flowers per spathe | 1 | Leaf length | 11.38 |
| 2 | Leaf width | 12.55 | ||
| 3 | Length of the pinna part | 4.54 | ||
| 4 | Number of pinnae per leaf | 7.01 | ||
| 5 | Pinna length | 4.56 | ||
| 6 | Pinna width | 6.23 | ||
| 7 | Spathe length | 12.37 | ||
| 8 | Spathe width | 5.94 | ||
| 9 | Length of spathe stem | 4.66 | ||
| 10 | Spathe weight | 7.35 | ||
| 11 | N | 4.4 | ||
| 12 | P | 2.37 | ||
| 13 | K | 10.99 | ||
| 14 | Ca | 3.27 | ||
| 15 | Mg | 2.38 | ||
| 5 | Weight of pollen grains per spathe | 1 | Leaf length | 12.57 |
| 2 | Leaf width | 8.15 | ||
| 3 | Length of the pinna part | 2.23 | ||
| 4 | Number of pinnae per leaf | 9.4 | ||
| 5 | Pinna length | 2.97 | ||
| 6 | Pinna width | 2.17 | ||
| 7 | Spathe length | 2.32 | ||
| 8 | Spathe width | 10.09 | ||
| 9 | Length of spathe stem | 17.85 | ||
| 10 | Spathe weight | 2.44 | ||
| 11 | N | 5.90 | ||
| 12 | P | 1.91 | ||
| 13 | K | 12.14 | ||
| 14 | Ca | 5.37 | ||
| 15 | Mg | 4.48 | ||
| 6 | Pollen grains’ viability | 1 | Leaf length | 9.40 |
| 2 | Leaf width | 4.65 | ||
| 3 | Length of the pinna part | 9.44 | ||
| 4 | Number of pinnae per leaf | 2.89 | ||
| 5 | Pinna length | 6.25 | ||
| 6 | Pinna width | 7.24 | ||
| 7 | Spathe length | 6.23 | ||
| 8 | Spathe width | 3.28 | ||
| 9 | Length of spathe stem | 9.37 | ||
| 10 | Spathe weight | 14.92 | ||
| 11 | N | 5.76 | ||
| 12 | P | 3.41 | ||
| 13 | K | 4.95 | ||
| 14 | Ca | 3.75 | ||
| 15 | Mg | 8.46 | ||
| 7 | Pollen grains’ length | 1 | Leaf length | 4.60 |
| 2 | Leaf width | 5.24 | ||
| 3 | Length of the pinna part | 3.97 | ||
| 4 | Number of pinnae per leaf | 3.46 | ||
| 5 | Pinna length | 5.16 | ||
| 6 | Pinna width | 8.70 | ||
| 7 | Spathe length | 8.37 | ||
| 8 | Spathe width | 13.35 | ||
| 9 | Length of spathe stem | 19.78 | ||
| 10 | Spathe weight | 2.87 | ||
| 11 | N | 5.99 | ||
| 12 | P | 3.47 | ||
| 13 | K | 3.52 | ||
| 14 | Ca | 6.16 | ||
| 15 | Mg | 5.36 | ||
| 8 | Pollen grains’ width | 1 | Leaf length | 5.98 |
| 2 | Leaf width | 4.51 | ||
| 3 | Length of the pinna part | 2.69 | ||
| 4 | Number of pinnae per leaf | 3.69 | ||
| 5 | Pinna length | 3.35 | ||
| 6 | Pinna width | 8.80 | ||
| 7 | Spathe length | 4.51 | ||
| 8 | Spathe width | 4.80 | ||
| 9 | Length of spathe stem | 30.59 | ||
| 10 | Spathe weight | 7.03 | ||
| 11 | N | 2.23 | ||
| 12 | P | 3.86 | ||
| 13 | K | 7.97 | ||
| 14 | Ca | 4.75 | ||
| 15 | Mg | 5.23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Sager, S.M.; Abdel-Sattar, M.; Al-Obeed, R.S.; Almady, S.S.; Aboukarima, A.M. Modeling the Performance Parameters of Pollen Grains of Male Date Palms Using an Artificial Neural Network Based on the Mineral Composition and Morphological Properties of Their Leaves. Horticulturae 2024, 10, 741. https://doi.org/10.3390/horticulturae10070741
Al-Sager SM, Abdel-Sattar M, Al-Obeed RS, Almady SS, Aboukarima AM. Modeling the Performance Parameters of Pollen Grains of Male Date Palms Using an Artificial Neural Network Based on the Mineral Composition and Morphological Properties of Their Leaves. Horticulturae. 2024; 10(7):741. https://doi.org/10.3390/horticulturae10070741
Chicago/Turabian StyleAl-Sager, Saleh M., Mahmoud Abdel-Sattar, Rashid S. Al-Obeed, Saad S. Almady, and Abdulwahed M. Aboukarima. 2024. "Modeling the Performance Parameters of Pollen Grains of Male Date Palms Using an Artificial Neural Network Based on the Mineral Composition and Morphological Properties of Their Leaves" Horticulturae 10, no. 7: 741. https://doi.org/10.3390/horticulturae10070741
APA StyleAl-Sager, S. M., Abdel-Sattar, M., Al-Obeed, R. S., Almady, S. S., & Aboukarima, A. M. (2024). Modeling the Performance Parameters of Pollen Grains of Male Date Palms Using an Artificial Neural Network Based on the Mineral Composition and Morphological Properties of Their Leaves. Horticulturae, 10(7), 741. https://doi.org/10.3390/horticulturae10070741