
Temporary Immersion System as an Innovative Approach for In Vitro Propagation of Sorbus domestica L.



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Cultural Environment
2.2. Data Collection and Statistical Analysis
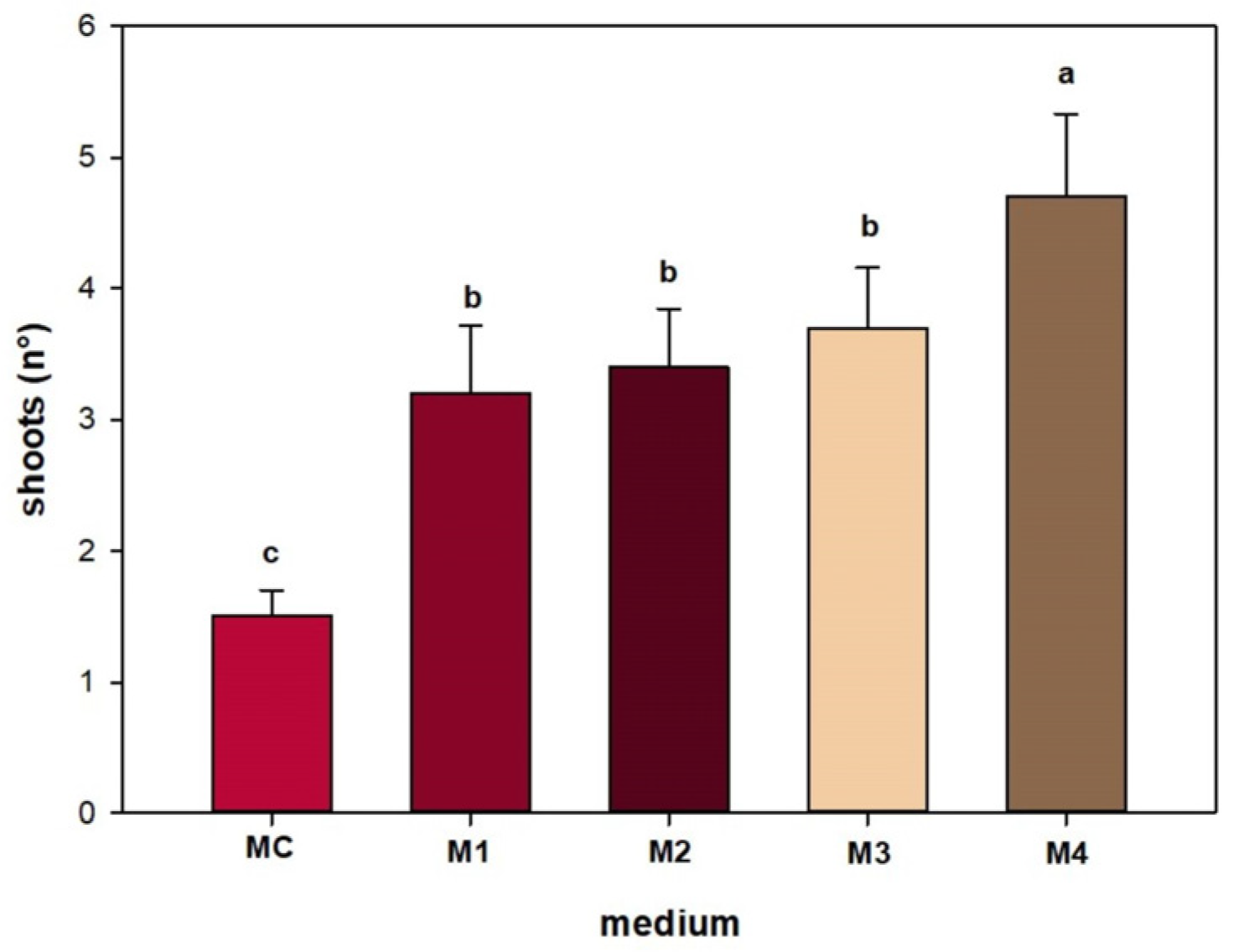
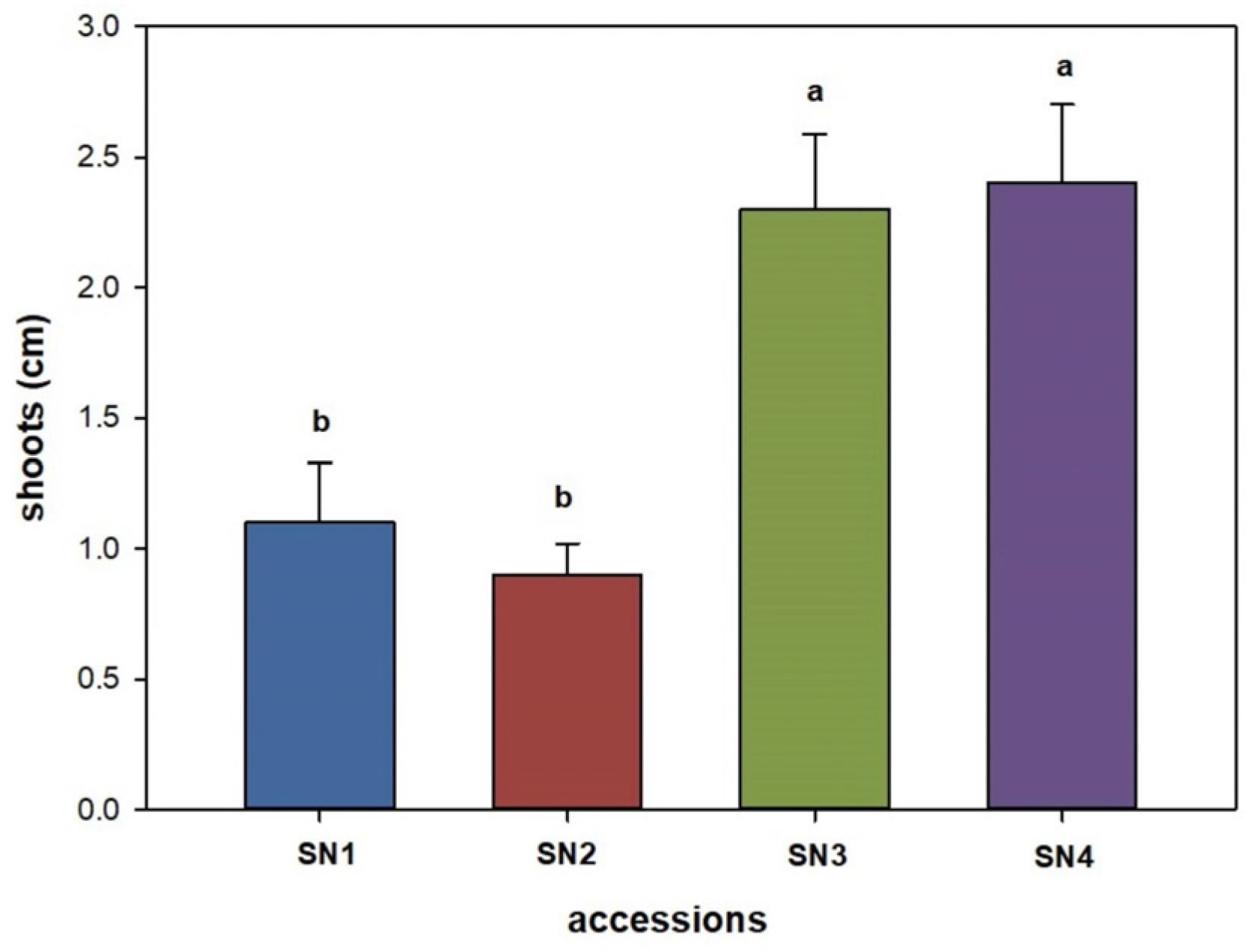
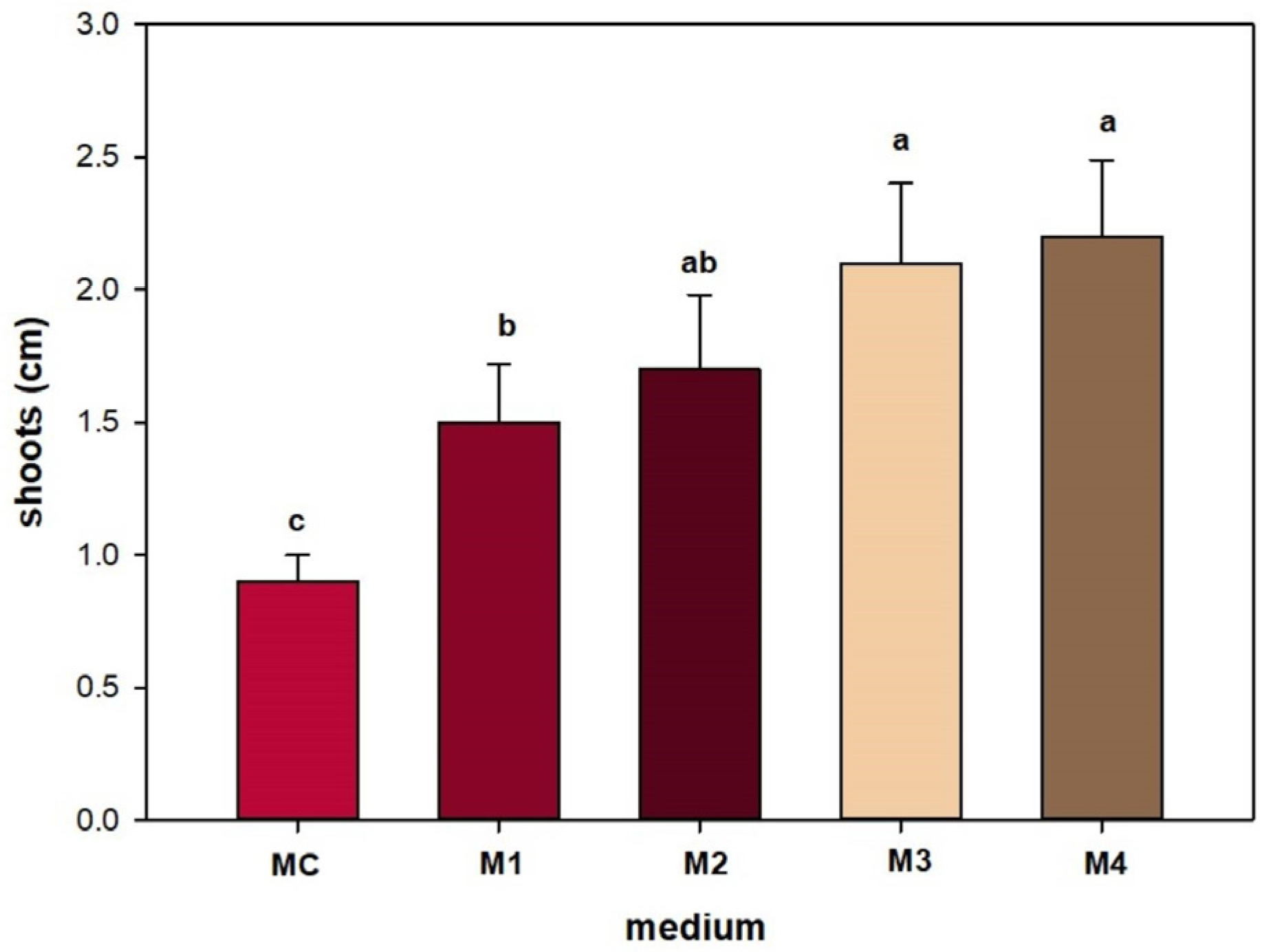
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, M.; Tetsuo, O.T.; Gao, Y.-D.; Xu, B.; Zhu, Z.-M.; Ju, W.-B.; Gao, X.-F. Molecular phylogenetics and historical biogeography of Sorbus sensu stricto (Rosaceae). Mol. Phylogenetics Evol. 2017, 111, 76–86. [Google Scholar] [CrossRef]
- Špíšek, Z.; Uherková, A.; Svitok, M.; Vašut, R.J. Sorbus domestica L. at its northern Pannonian distribution limits: Distribution of individuals, fruit shapes and dendrometric characteristics. Dendrobiology 2018, 80, 37–47. [Google Scholar] [CrossRef]
- Brütsh, U.; Rotach, P. Der Speierling (Sorbus domestica L.) in der Schweiz: Verbreitung, Ökologie, Standsortsanspruche, Konkurrenzkraft und Waldbauliche Eignung. Schweiz. Z. Forstwes. 1993, 144, 967–991. [Google Scholar]
- Sottile, F.; Girgenti, V.; Del Signore, M.B.; Peano, C. Quality of autochthonous Sicilian plums. Ital. J. Food Sci. 2015, 27, 320–329. [Google Scholar]
- Hobbie, S.E.; Grimm, N.B. Nature-based approaches to managing climate change impacts in cities. Philos. Trans. R. Soc. B 2020, 375, 20190124. [Google Scholar] [CrossRef] [PubMed]
- Pereira, P.; Wang, F.; Inacio, M.; Kalinauskas, M.; Bogdzevič, K.; Bogunovic, I.; Zhao, W.; Barcelo, D. Nature-based solutions for carbon sequestration in urban environments. Curr. Opin. Environ. Sci. Health 2024, 37, 100536. [Google Scholar] [CrossRef]
- Paleari, S. The EU policy on climate change, biodiversity and circular economy: Moving towards a Nexus approach. Environ. Sci. Policy 2024, 151, 103603. [Google Scholar] [CrossRef]
- Salik, A.N. COP21 Paris. Strateg. Stud. 2017, 37, 74–89. [Google Scholar] [CrossRef]
- Golicz, K.; Bellingrath-Kimura, S.; Breuer, L.; Wartenberg, A.C. Carbon accounting in European agroforestry systems—Key research gaps and data needs. Curr. Res. Environ. Sustain. 2022, 4, 100134. [Google Scholar] [CrossRef]
- Bellini, E.; Giordani, E.; La Malfa, S. I fruttiferi minori in Italia, una risorsa tradizionale per l’innovazione frutticola: Il kaki e il melograno come casi di studio. Italus Hortus 2010, 17, 75–90. [Google Scholar]
- Piagnani, M.C.; Debellini, C.; Lo Scalzo, R. Phyllometry and carpometry, chemical and functional characterization of fruits of Sorbus domestica L. (service tree) selections. J. Berry Res. 2012, 2, 7–22. [Google Scholar] [CrossRef]
- Bignami, C. Il sorbo. In Conservazione dei Fruttiferi Minori, Proceedings of the National Workshop on EC Project GENRES 29, Firenze 27–28 November 1998; Department of Horticulture, University of Florence: Florence, Italy, 1998. [Google Scholar]
- Pant, M.; Bhandari, A.; Husen, A. Adventitious root formation and clonal propagation of forest-based tree species. In Environmental, Physiological and Chemical Controls of Adventitious Rooting in Cuttings; Husen, A., Ed.; Academic Press: Cambridge, MA, USA, 2022; Chapter 22; pp. 471–490. [Google Scholar]
- Lall, S.; Mandegaran, Z.; Roberts, A. Shoot multiplication and adventitious regeneration in Sorbus aucuparia. Plant Cell Tissue Organ Cult. 2006, 85, 23–29. [Google Scholar] [CrossRef]
- Arrillaga, I.; Marzo, T.; Segura, J. Micropropagation of juvenile and adult Sorbus domestica L. Plant Cell Tissue Organ Cult. 1991, 27, 341–348. [Google Scholar] [CrossRef]
- Meier-Dinkel, A. In vitro Vermehrung von Speierling (Sorbus domestica L.). Corminaria 1998, 9, 9–13. [Google Scholar]
- Miko, M.; Gažo, J.; Biroščíková, M. In vitro klonové množenie genetických zdrojov jarabiny oskorušovej (Sorbus domestica L.) zúzemia Slovenska. Acta Fytotech. Zootech. 2004, 7, 85–89. [Google Scholar]
- Nikolaou, P.; Zagas, D.; Scaltsoyiannes, V.; Balas, E.; Xilogianni, V.; Tsoulpha, P.; Tsaktsira, M.; Voulgaridou, E.; Iliev, I.; Triantafyllou, K.; et al. Advances in the micropropagation of service tree (Sorbus domestica L.). Propag. Ornam. Plants 2008, 8, 154–157. [Google Scholar]
- Durkovič, J.; Mišalová, A. Wood formation during ex vitro acclimatisation in micropropagated true service tree (Sorbus domestica L.). Plant Cell Tissue Organ Cult. 2009, 96, 343–348. [Google Scholar] [CrossRef]
- Malá, J.; Cvrčková, H.; Máchová, P.; Dostál, J. Mikropropagace Jeřábu Oskeruše (Sorbus domestica L.); Certified Methodology 4/2011; Forestry and Game Management Research Institute: Strnady, Czech Republic, 2011; pp. 1–17. (In Czech) [Google Scholar]
- Ördögh, M.; Jàmbor-Benczùr, E.; Tilly-Màndy, A.; Lelik, L. The effects of growth regulators in proliferation of Sorbus rediliana “Burokvolgy”. Int. J. Hortic. Sci. 2006, 12, 77–83. [Google Scholar] [CrossRef]
- Jana, Š.; Jiří, V.; Daniel, Z. Micropropagation as a Tool for the Conservation of Autochthonous Sorbus Species of Czechia. Plants 2023, 12, 488. [Google Scholar] [CrossRef]
- Bengi, E.; Yeald, E. In vitro micropropagation of Anthemis xylopoda O. Schwarz, a Critically Endangered Species from Turkey. Pak. J. Biol. Sci. 2005, 8, 691–695. [Google Scholar]
- Trejgell, A.; Michalska, M.; Tretyn, A. Micropropagation of Senecio Macrophyllus M. Bieb. Acta Biol. Cracoviensia Ser. Bot. 2010, 52, 67–72. [Google Scholar] [CrossRef]
- Gianguzzi, V.; Barone, G.; Di Gristina, E.; Sottile, F.; Domina, G. Micropropagation of Endemic Endangered Taxa of the Italian Flora: Adenostyles alpina subsp. macrocephala (Asteraceae), as a Case Study. Plants 2023, 12, 1530. [Google Scholar] [CrossRef] [PubMed]
- Iliev, I.A. Factors affecting the axillary and adventitious shoot formation in woody plants in vitro (Conference Paper). Acta Horticulturae 2017, 1155, 15–28. [Google Scholar] [CrossRef]
- Liu, C.; Callow, P.; Rowland, L.J.; Hancock, J.F.; Song, G.Q. Adventitious shoot regeneration from leaf explants of southern highbush blueberry cultivars. Plant Cell Tissue Organ Cult. 2010, 103, 137–144. [Google Scholar] [CrossRef]
- Durkovič, J.; Mišalová, A. Micropropagation of temperate noble hardwoods: An overview. Funct. Plant Sci. Bio-technol. 2008, 2, 1–19. [Google Scholar]
- Gianguzzi, V.; Di Gristina, E.; Barone, G.; Sottile, F.; Domina, G. Seed germination and vegetative and in vitro propagation of Hieracium lucidum subsp. lucidum(Asteraceae), a critically endangered endemic taxon of the Sicilian flora. PeerJ 2024, 12, e16839. [Google Scholar] [CrossRef]
- Werbrouck, S.P.O.; Van der Jeugt, B.; Dewitte, W.; Prinsen, E.; Van Onckelen, H.A.; Debergh, P.C. The metabolism of benzyladenine in Spathiphyllum floribundum ‘Schott Petite’ in relation to acclimatisation problems. Plant Cell Tissue Organ Cult. 1995, 14, 662–665. [Google Scholar] [CrossRef]
- Centeno, M.L.; Rodrígue, A.; Feito, I.; Fernández, B. Relationship between endogenous auxin and cytokinin levels and morphogenic responses in Actinidia deliciosa tissue cultures. Plant Cell Rep. 1996, 16, 58–62. [Google Scholar] [CrossRef]
- Tarkowská, D.; Doležal, K.; Tarkowski, P.; Astot, C.; Holub, J.; Fuksová, K.; Schmülling, T.; Sandberg, G.; Strnad, M. Identification of new aromatic cytokinins in Arabidopsis thaliana and Populus × canadensis leaves by LC-(+)ESI-MS and capillary liquid chromatography/frit-fast atom bombardment mass spektrometry. Physiol. Plant. 2003, 117, 579–590. [Google Scholar] [CrossRef]
- Strnad, M.; Hanuš, J.; Vanˇek, T.; Kamínek, M.; Ballantine, J.A.; Fussell, B.; Hanke, D.E. Meta-topolin, a highly active aromatic cytokinin from poplar leaves (Populus × canadensis Moench., cv. Robusta). Phytochemistry 1997, 45, 213–218. [Google Scholar] [CrossRef]
- Chalupa, V. European hardwoods. In Cell and Tissue Culture in Forestry; Bonga, J.M., Durzan, D.J., Eds.; Martinus Nijhoff Publishers: Dordrecht, The Netherlands, 1987; Volume 3, pp. 224–246. [Google Scholar]
- Phillips, G.C.; Garda, M. Plant tissue culture media and practices: An overview. Vitr. Cell. Dev. Biol.-Plant 2019, 55, 242–257. [Google Scholar] [CrossRef]
- Dutra, L.F.; Wendling, I.; Brondani, G.E. A micropropagação de eucalipto [The micropropagation of eucalyptus]. Pesqui. Florest. Bras. 2009, 58, 49–59. [Google Scholar]
- Oliveira, M.L.; Xavier, A.; Penchel, R.M.; Santos, A.F. Multiplicação in vitro de Eucalyptus grandis x E. urophylla cultivado em meio semissólido e em biorreator de imersão temporária [In vitro multiplication of Eucalyptus grandis × E. urophylla grown in semisolidù medium and in temporary immersion bioreactor]. Sci. For. 2011, 39, 309–315. [Google Scholar]
- Welander, M.; Persson, J.; Asp, H.; Zhu, L.H. Evaluation of a new vessel system based on temporary immersion system for micropropagation. Sci. Hortic. 2014, 179, 227–232. [Google Scholar] [CrossRef]
- Businge, E.; Trifonova, A.; Schneider, C.; Rödel, P.; Egertsdotter, U. Evaluation of a new temporary immersion bioreactor system for micropropagation of cultivars of eucalyptus, birch and fir. Forests 2017, 8, 196. [Google Scholar] [CrossRef]
- Etienne, H.; Berthouly, M. Temporary immersion systems in plant micropropagation. Plant Cell Tissue Organ Cult. 2002, 69, 215–231. [Google Scholar] [CrossRef]
- Sottile, F.; Caltagirone, C.; Peano, C.; Del Signore, M.B.; Barone, E. Can the Caper (Capparis spinosa L.) Still Be Considered a Difficult-to-Propagate Crop? Horticulturae 2021, 7, 316. [Google Scholar] [CrossRef]
- Alvard, D.; Cote, F.; Teisson, C. Comparison of methods of liquid medium culture for banana micropropagation. Effects of temporary immersion of explants. Plant Cell Tissue Organ Cult. 1993, 32, 55–60. [Google Scholar] [CrossRef]
- Akdemir, H.; Süzerer, V.; Onay, A.; Tilkat, E.; Ersali, Y.; Çiftçi, Y.O. Micropropagation of the pistachio and its rootstocks by temporary immersion system. Plant Cell Tissue Organ Cult. 2014, 117, 65–76. [Google Scholar] [CrossRef]
- Albarran, J.; Bertrand, B.; Lartaud, M.; Etienne, H. Cycle characteristics in a temporary immersion bioreactors affects regeneration, morphology, water and mineral status of coffee (Coffea arabica) somatic embryos. Plant Cell Tissue Organ Cult. 2005, 81, 27–36. [Google Scholar] [CrossRef]
- Zhu, L.H.; Li, X.Y.; Welander, M. Optimisation of growing conditions of the apple rootstock M26 grown in RITA® container using temporary immersion principle. Plant Cell Tissue Organ Cult. 2005, 81, 313–318. [Google Scholar] [CrossRef]
- McAlister, B.; Finnie, J.; Watt, M.P.; Blakeway, F. Use of temporary immersion system (RITA®) for production of commercial Eucalyptus clones in Mondi Forests (SA). Plant Cell Tissue Organ Cult. 2005, 81, 347–358. [Google Scholar] [CrossRef]
- Debnath, S.C. A two-step procedure for in vitro multiplication of cloudberry (Rubus chamaemorus L.) shoots using bioreactor. Plant Cell Tissue Organ Cult. 2007, 88, 185–191. [Google Scholar] [CrossRef]
- Sankar-Thomas, Y.D.; Surminski, K.S.; Lieberi, R. Plant regeneration via somatic embryogenesis of Camptotheca acuminata in temporary immersion system (TIS). Plant Cell Tissue Organ Cult. 2008, 95, 163–173. [Google Scholar] [CrossRef]
- Escalona, M.; Lorenzo, J.C.; Gonzales, B.; Daquinta, M.; Gonzalez, J.L.; Desjardins, Y.; Borroto, C.G. Pineapple (Ananas comosus L. Merr) micropropagation in temporary immersion systems. Plant Cell Rep. 1999, 18, 743–748. [Google Scholar] [CrossRef]
- Welander, M.; Zhu, L.H.; Li, X.Y. Factors influencing conventional and semi-automated micropropagation. Propag. Ornam. Plants 2007, 7, 103–111. [Google Scholar]
- Roels, S.; Noceda, C.; Escalona, M.; Sandoval, J.; Canal, M.J.; Rodríguez, R.; Debergh, P. The effect of headspace renewal in a temporary immersion bioreactor of plantain (Musa AAB) shoot proliferation and quality. Plant Cell Tissue Organ Cult. 2006, 84, 155–163. [Google Scholar] [CrossRef]
- Xiao, Y.; Niu, G.; Kozai, T. Development and application of photoautotrophic micropropagation plant system. Plant Cell Tissue Organ Cult. 2011, 105, 149–158. [Google Scholar] [CrossRef]
- Sottile, F.; Del Signore, M.B.; Giuggioli, N.; Peano, C. The potential of the Sorb (Sorbus domestica L.) as a minor fruit species in the Mediterranean areas: Description and quality traits of underutilized accessions. Prog. Nutr. 2017, 19, 41–48. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F.A. Revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Nitsch, J.P.; Nitsch, C. Haploid plants from pollen grains. Science 1969, 163, 85–87. [Google Scholar] [CrossRef]
- Gianguzzi, V.; Inglese, P.; Barone, E.; Sottile, F. In Vitro Regeneration of Capparis spinosa L. by Using a Temporary Immersion System. Plants 2019, 8, 177. [Google Scholar] [CrossRef]
- Vidal, N.; Sánchez, C. Use of bioreactor systems in the propagation of forest trees. Eng. Life Sci. 2019, 19, 896–915. [Google Scholar] [CrossRef] [PubMed]
- Schönherr, J. Characterization of aqueous pores in plant cuticles and permeation of ionic solutes. J. Exp. Bot. 2006, 57, 2471–2491. [Google Scholar] [CrossRef] [PubMed]
- De Klerk, G.J.; Ter Brugge, J. Micropropagation of dahlia in static liquid medium using slow-release tools of medium ingredients. Sci. Hortic. 2011, 127, 542–547. [Google Scholar] [CrossRef]
- Benelli, C.; De Carlo, A. In vitro multiplication and growth improvement of Olea europea L. cv Canino with temporary immersion system (Plantform™). 3 Biotech 2018, 8, 317. [Google Scholar] [CrossRef] [PubMed]
- Regueira, M.; Rial, E.; Blanco, B.; Bogo, B.; Aldrey, A.; Correa, B.; Varas, E.; Sánchez, C.; Vidal, N. Micropropagation of axillary shoots of Salix viminalis using a temporary immersion system. Trees 2018, 32, 61–71. [Google Scholar] [CrossRef]
- Gatti, E.; Sgarbi, E.; Ozudogru, E.A.; Lambardi, M. The effect of Plantform™ bioreactor on micropropagation of Quercus robur in comparison to a conventional in vitro culture on gelled medium, and assessment of the microenvironment influence on leaf structure. Plant Biosyst. 2017, 151, 1129–1136. [Google Scholar] [CrossRef]
- Vidal, N.; Blanco, B.; Cuenca, B. A temporary immersion system for micropropagation of axillary shoots of hybrid chestnut. Plant Cell Tissue Organ Cult. 2015, 126, 229–243. [Google Scholar] [CrossRef]
- Nayyef, M.N.; Ali Abdulhussein, M.A.; Ali Almusawi, A.H. The efficiency of plant form as temporary immersion system in bud and multiplication shoot via direct organogenesis of date palm under LED light. Plant Arch. 2019, 19, 1419–1426. [Google Scholar]
- Bairu, M.W.; Stirk, W.A.; Doležal, K.; Van Staden, J. Optimizing the micropropagation protocol for the endangered Aloe polyphylla: Can meta-topolin and its derivatives serve as replacement for benzyladenine and zeatin? Plant Cell Tissue Organ Cult. 2007, 90, 15–23. [Google Scholar] [CrossRef]
- Werbrouck, S.P.O.; Strnad, M.; Van Onckelen, H.A.; Debergh, P.C. Meta-topolin, an alternative to benzyladenine in tissue culture. Physiol. Plant. 2008, 98, 291–297. [Google Scholar] [CrossRef]
- Kubalakova, M.; Strnad, M. The effects of aromatic cytokinins on micropropagation and regeneration of sugar beet in vitro. Biol. Plant 1992, 34, 578–579. [Google Scholar]
- Chiancone, B.; Karasawa, M.M.G.; Gianguzzi, V.; Abdelgalel, A.M.; Bárány, I.; Testillano, D.; Marinoni, D.; Botta, D.; Germanà, M.A. Early embryo achievement through isolated microspore culture in Citrus clementina Hort. ex Tan., cvs. ‘Monreal Rosso’ and ‘Nules’. Front. Plant Sci. 2015, 6, 413. [Google Scholar] [CrossRef]
- Chiancone, B.; Martorana, L.; Gianguzzi, V.; Germanà, M.A. The effects of meta-Topolin and benzyladenine on in vitro organogenesis from epicotyl cuttings of Troyer citrange (Citrus sinensis [L.] Osbeck × Poncirus trifoliata [L.] Raf.). Acta Hortic. 2017, 1155, 185–192. [Google Scholar] [CrossRef]
- Kucharska, D.; Orlikowska, T.; Maciorowski, R.; Kunka, M.; Wójcik, D.; Pluta, S. Application of meta-Topolin for improving micropropagation of gooseberry (Ribes grossularia). Sci. Hortic. 2020, 272, 109529. [Google Scholar] [CrossRef]
- Malá, J.; Máchová, P.; Cvrčková, H.; Karady, M.; Novák, O.; Mikulík, J.; Hauserová, E.; Greplová, J.; Strnad, M.; Doležal, K. Micropropagation of wild service tree (Sorbus torminalis (L.) Crantz): The regulative role of different aromatic cytokinins during organogenesis. J. Plant Growth Regul. 2009, 28, 341–348. [Google Scholar] [CrossRef]
- Martini, A.N.; Papafotiou, M. Season and explant origin affect phenolic content, browning of explants, and micropropagation of ×Malosorbus florentina (Zucc.) Browicz. HortScience 2013, 48, 102–107. [Google Scholar] [CrossRef]
- Chalupa, V. In vitro propagation of mature trees of Sorbus aucuparia L. and field performance of micropropagated tree. J. For. Sci. 2002, 48, 529–535. [Google Scholar] [CrossRef]
- Ördögh, M.; Jambor-Benczúr, E.; Tilly-Mándy, A.; Lelik, L. Effects of different cytokinins on proliferation of Sorbus borbasii ‘Herkulesfürdö’. Propag. Ornam. Plants 2009, 9, 43–46. [Google Scholar]
- Jeong, B.R.; Sivanesan, I. Micropropagation of Sorbus commixta Hedl. Propag. Ornam. Plants 2015, 15, 142–146. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium | PGR | ||
|---|---|---|---|
| BAP (mg L−1) | mT (mg L−1) | IBA (mg L−1) | |
| MC | 0 | 0 | 0 |
| M1 | 1 | 0 | 0 |
| M2 | 0 | 1 | 0 |
| M3 | 1 | 0 | 0.5 |
| M4 | 0 | 1 | 0.5 |
| Medium | Accessions | |||
|---|---|---|---|---|
| SN1 | SN2 | SN3 | SN4 | |
| MC | 2.0 ± 0.26 e | 1.4 ± 0.17 c | 1.2 ± 0.10 c | 1.8 ± 0.18 e |
| M1 | 3.2 ± 0.42 d | 4.5 ± 0.58 b | 2.2 ± 0.40 b | 2.9 ± 0.45 d |
| M2 | 3.6 ± 0.42 c | 4.4 ± 0.51 b | 2.4 ± 0.24 b | 3.3 ± 0.42 c |
| M3 | 4.3 ± 0.29 b | 4.1 ± 0.43 b | 2.5 ± 0.38 b | 3.9 ± 0.55 b |
| M4 | 4.8 ± 0.65 a | 5.9 ± 0.73 a | 3.5 ± 0.46 a | 4.4 ± 0.46 a |
| Medium | Accessions | |||
|---|---|---|---|---|
| SN1 | SN2 | SN3 | SN4 | |
| MC | 0.68 ± 0.06 c | 0.73 ± 0.04 c | 1.1 ± 0.10 d | 1.1 ± 0.13 c |
| M1 | 0.89 ± 0.12 c | 0.94 ± 0.13 b | 2.0 ± 0.23 c | 2.1 ± 0.25 b |
| M2 | 1.08 ± 0.13 b | 0.93 ± 0.14 b | 2.4 ± 0.31 b | 2.5 ± 0.30 b |
| M3 | 1.30 ± 0.17 a | 1.20 ± 0.15 a | 2.8 ± 0.30 b | 3.1 ± 0.32 a |
| M4 | 1.50 ± 0.13 a | 1.10 ± 0.12 a | 3.1 ± 0.31 a | 3.3 ± 0.27 a |
| RGR | ||||
|---|---|---|---|---|
| Medium | Accessions | |||
| SN1 | SN2 | SN3 | SN4 | |
| MC | 0.9 | 0.98 | 1.2 | 1.3 |
| M1 | 1.5 | 1.6 | 1.8 | 2.4 |
| M2 | 2.6 | 2.0 | 2.3 | 3.2 |
| M3 | 3.0 | 3.5 | 4.2 | 5.4 |
| M4 | 4.3 | 4.7 | 5.2 | 8.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gianguzzi, V.; Sottile, F. Temporary Immersion System as an Innovative Approach for In Vitro Propagation of Sorbus domestica L. Horticulturae 2024, 10, 164. https://doi.org/10.3390/horticulturae10020164
Gianguzzi V, Sottile F. Temporary Immersion System as an Innovative Approach for In Vitro Propagation of Sorbus domestica L. Horticulturae. 2024; 10(2):164. https://doi.org/10.3390/horticulturae10020164
Chicago/Turabian StyleGianguzzi, Valeria, and Francesco Sottile. 2024. "Temporary Immersion System as an Innovative Approach for In Vitro Propagation of Sorbus domestica L." Horticulturae 10, no. 2: 164. https://doi.org/10.3390/horticulturae10020164
APA StyleGianguzzi, V., & Sottile, F. (2024). Temporary Immersion System as an Innovative Approach for In Vitro Propagation of Sorbus domestica L. Horticulturae, 10(2), 164. https://doi.org/10.3390/horticulturae10020164