
Study on Italian Onion Cultivars/Ecotypes towards Onion Yellow Dwarf Virus (OYDV) Infection
 , ,
, ,  ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and OYDV Inoculation
2.2. Evaluation of Susceptibility/Tolerance to OYDV
2.2.1. Assessment of Symptoms Severity
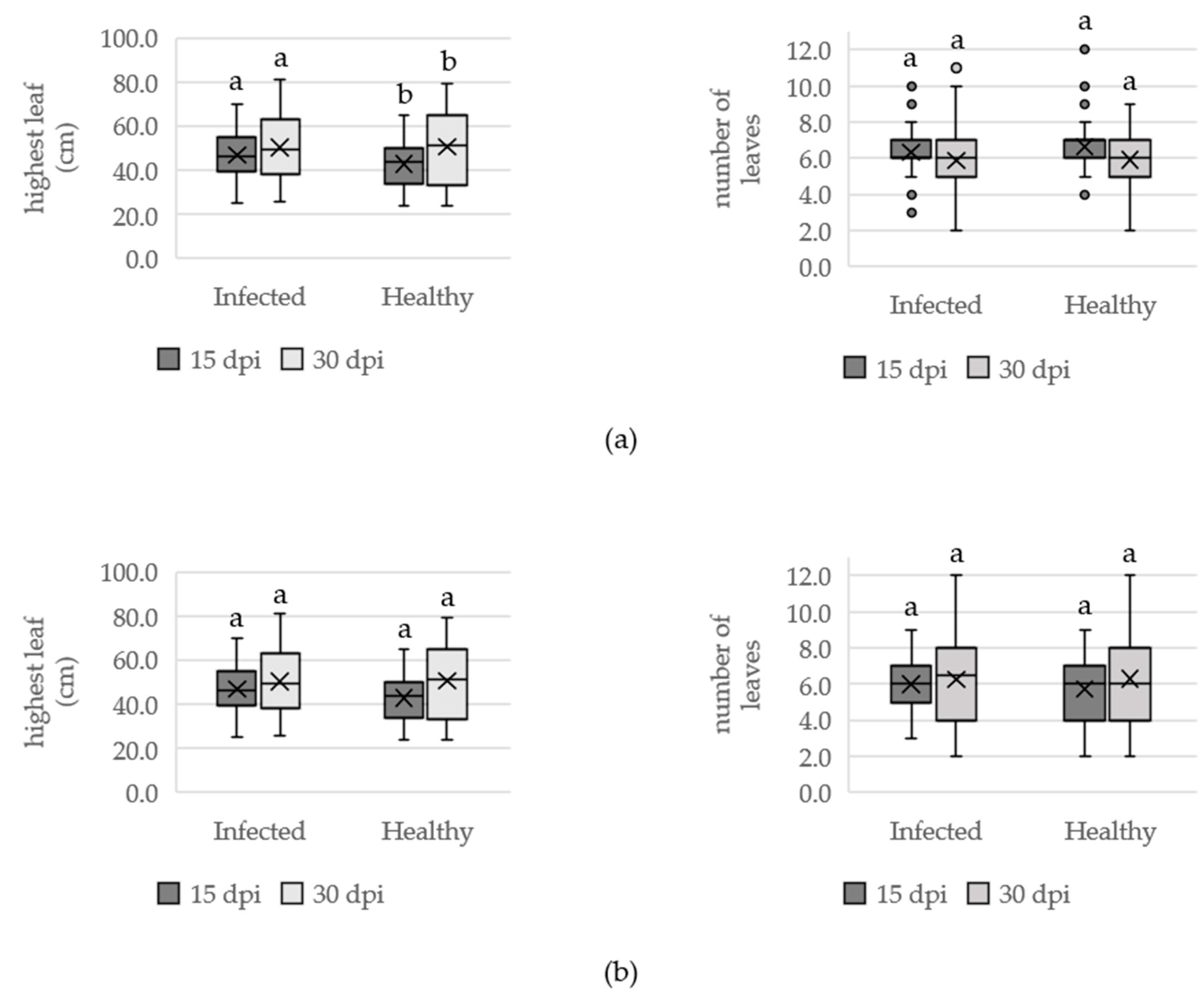
2.2.2. Effect of ODYV on Plant Growth, Bulb Long-Term Storage and Bulb Water Losses
2.3. SSR Genetic Analysis
2.3.1. DNA Extraction and PCR Analysis
2.3.2. Statistical Analysis
2.4. Conserved Motifs among EIF4e and EIF4g
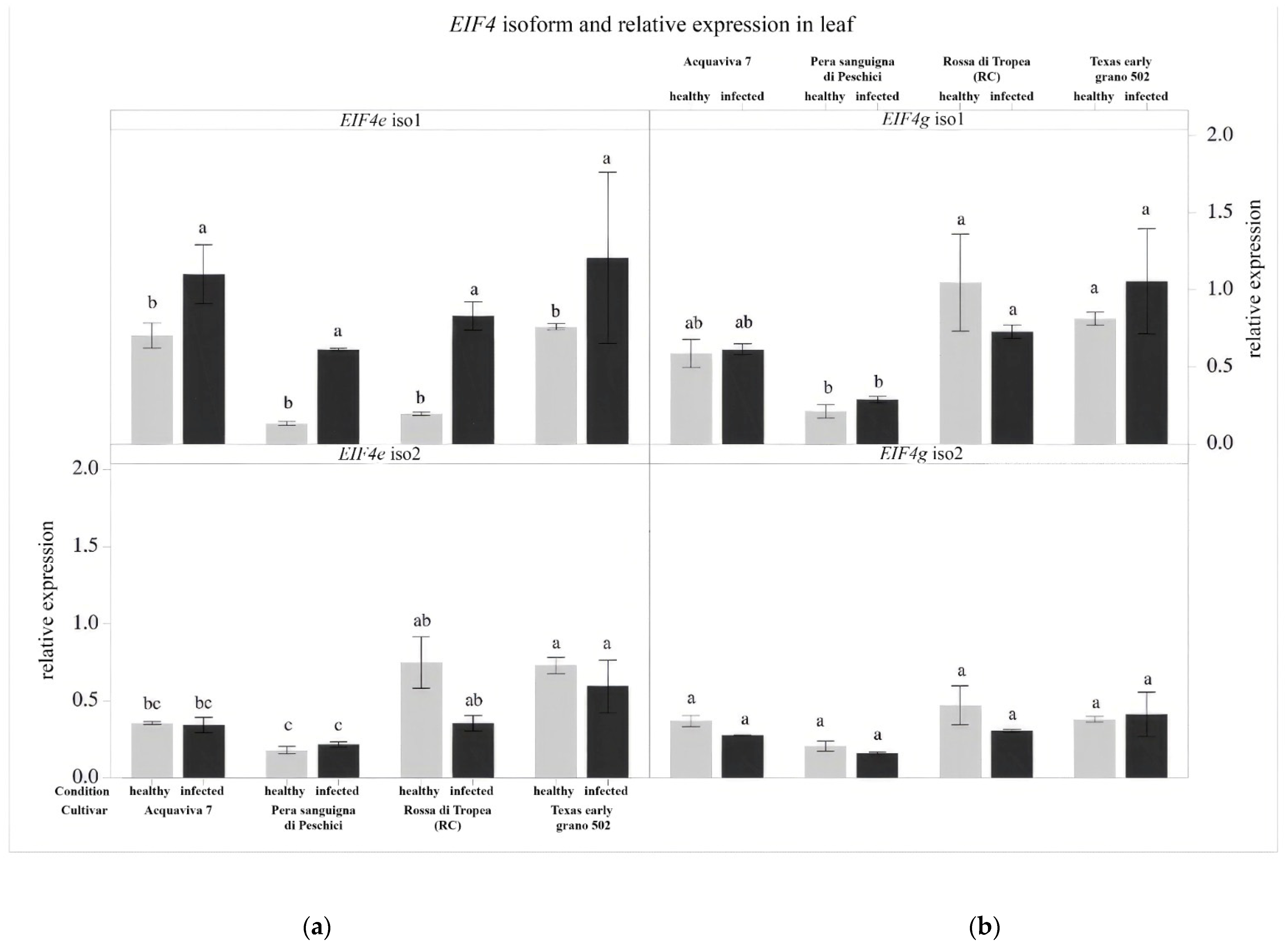
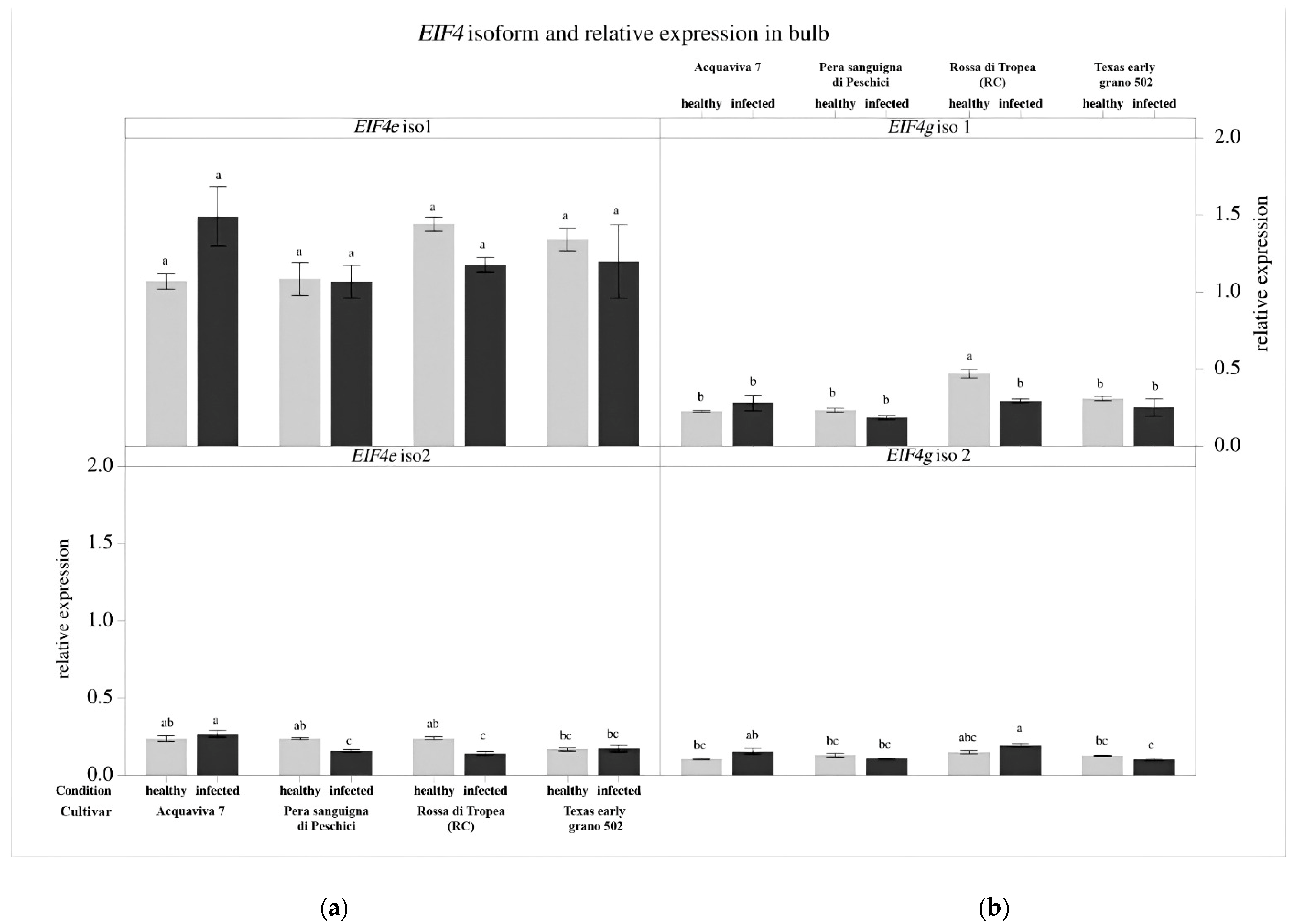
2.5. Gene Expression Analysis of the Eukaryotic Translation Initiation Factors (EIF)
2.5.1. Plant Material and RNA Extraction
2.5.2. Primers Design and Housekeeping Gene Detection
2.5.3. Reverse Transcription and Gene Expression Analysis by qPCR
3. Results
3.1. Evaluation of Susceptibility/Tolerance to OYDV
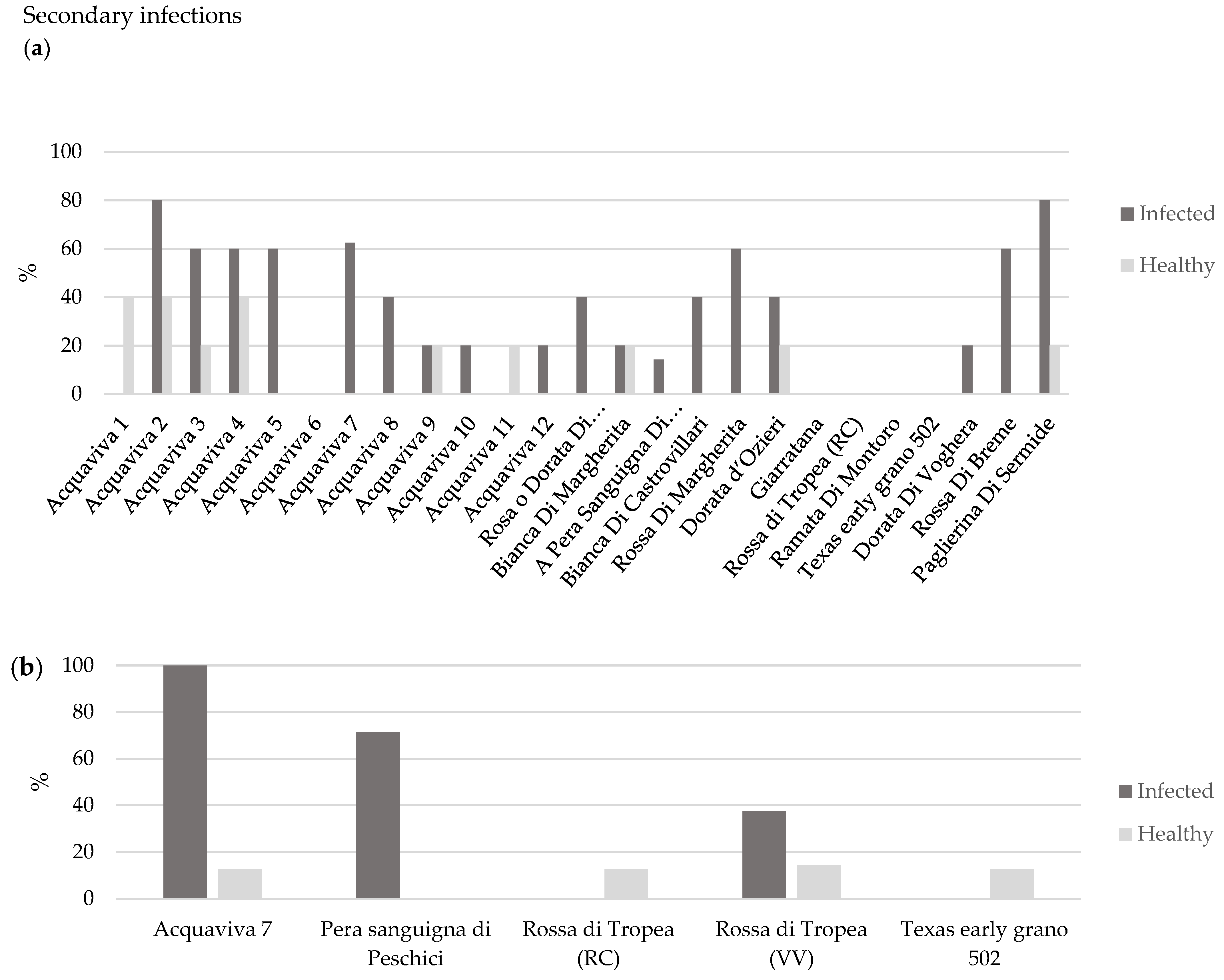
3.1.1. Assessment of Symptoms Severity
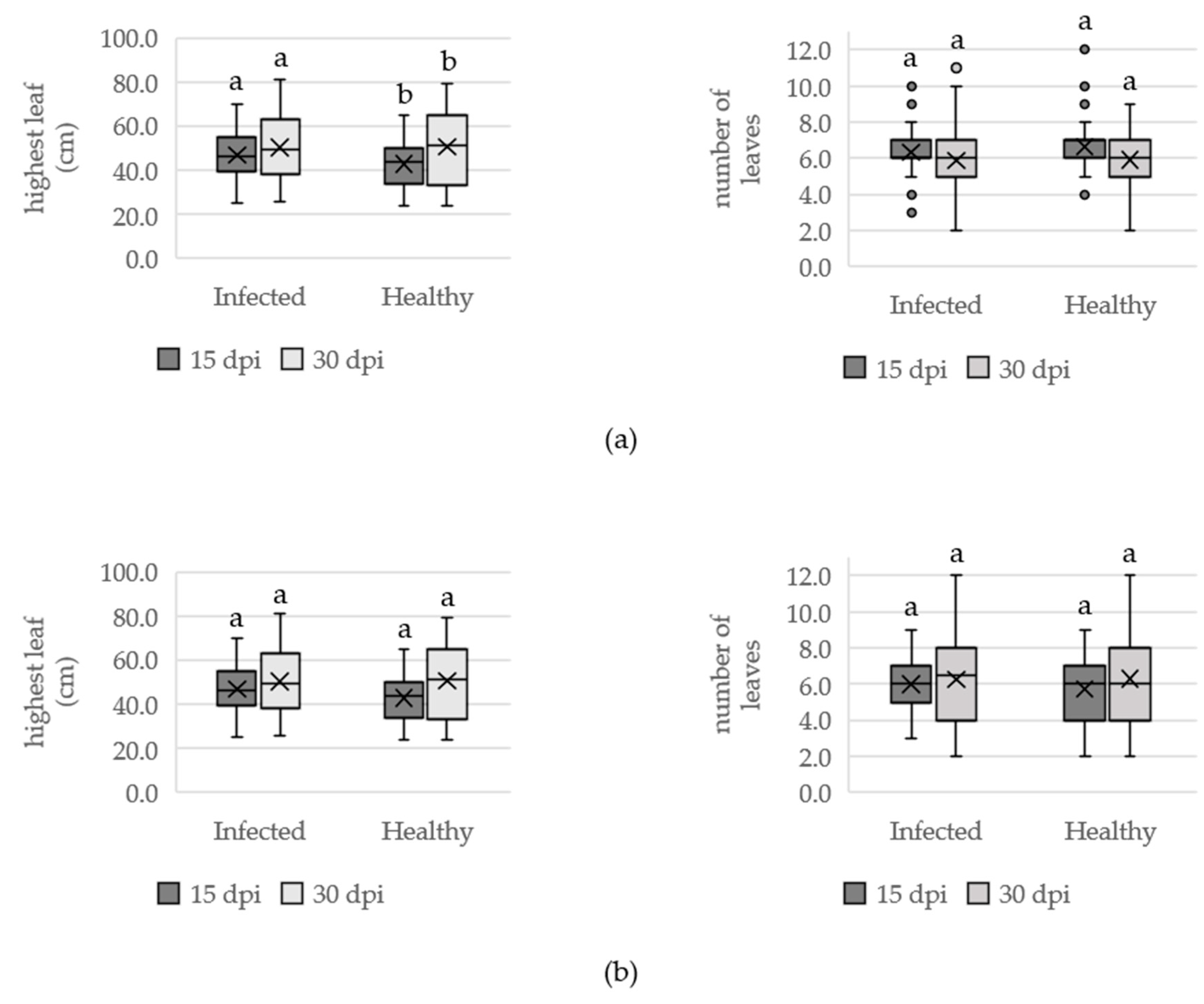
3.1.2. Effect of OYDV on Plant Growth
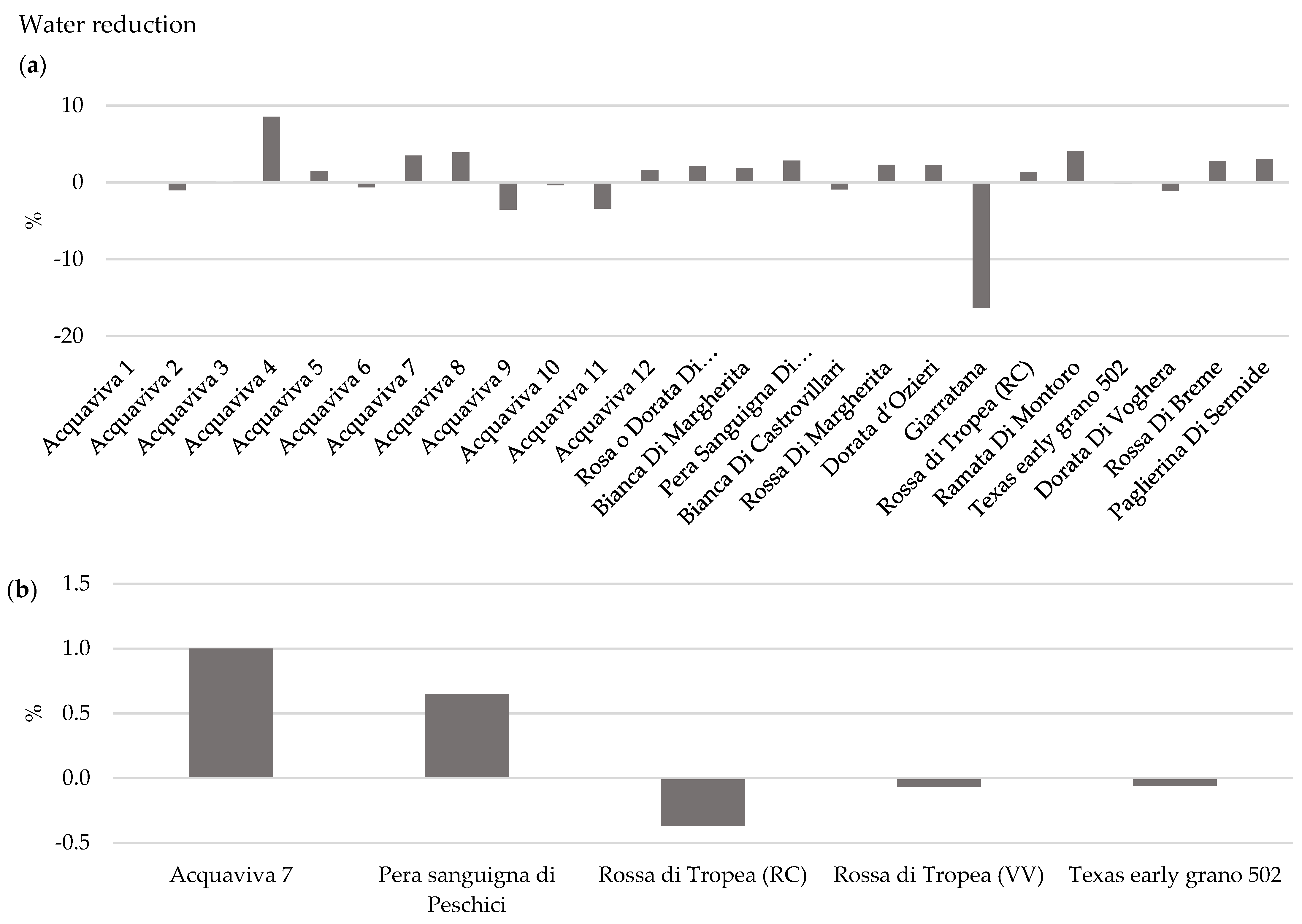
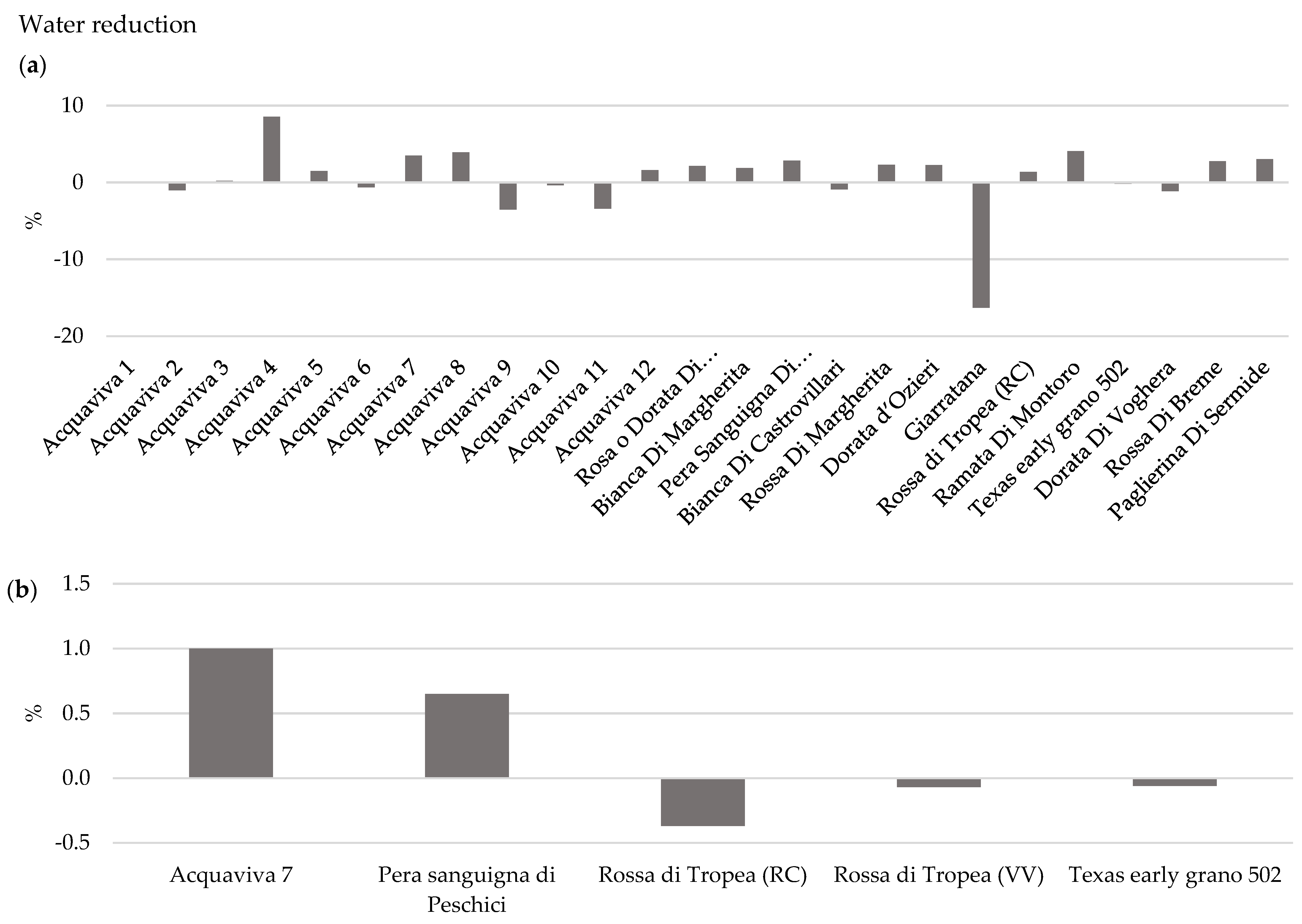
3.1.3. Effect of OYDV on Bulb Long-Term Storage and Water Loss
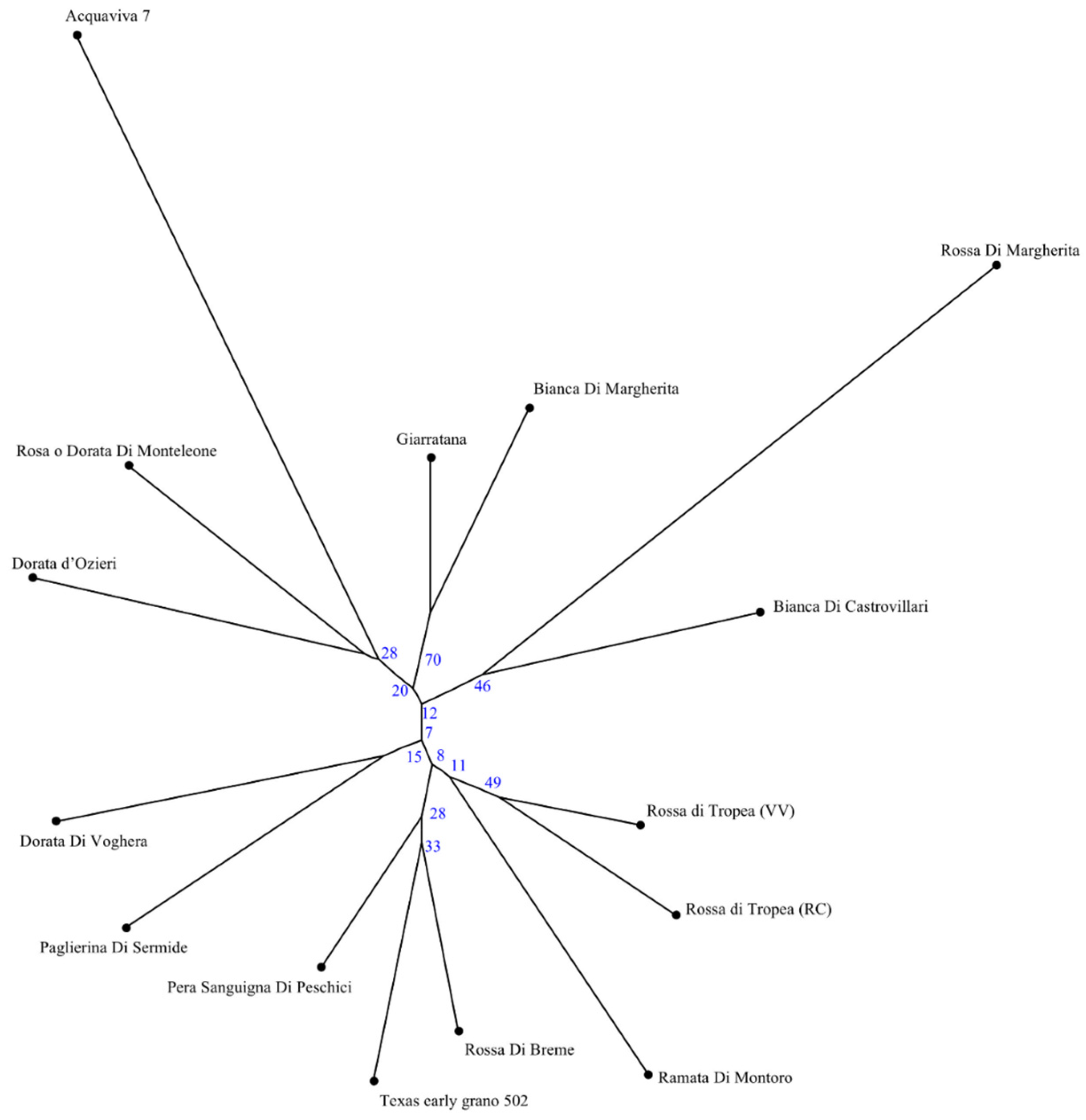
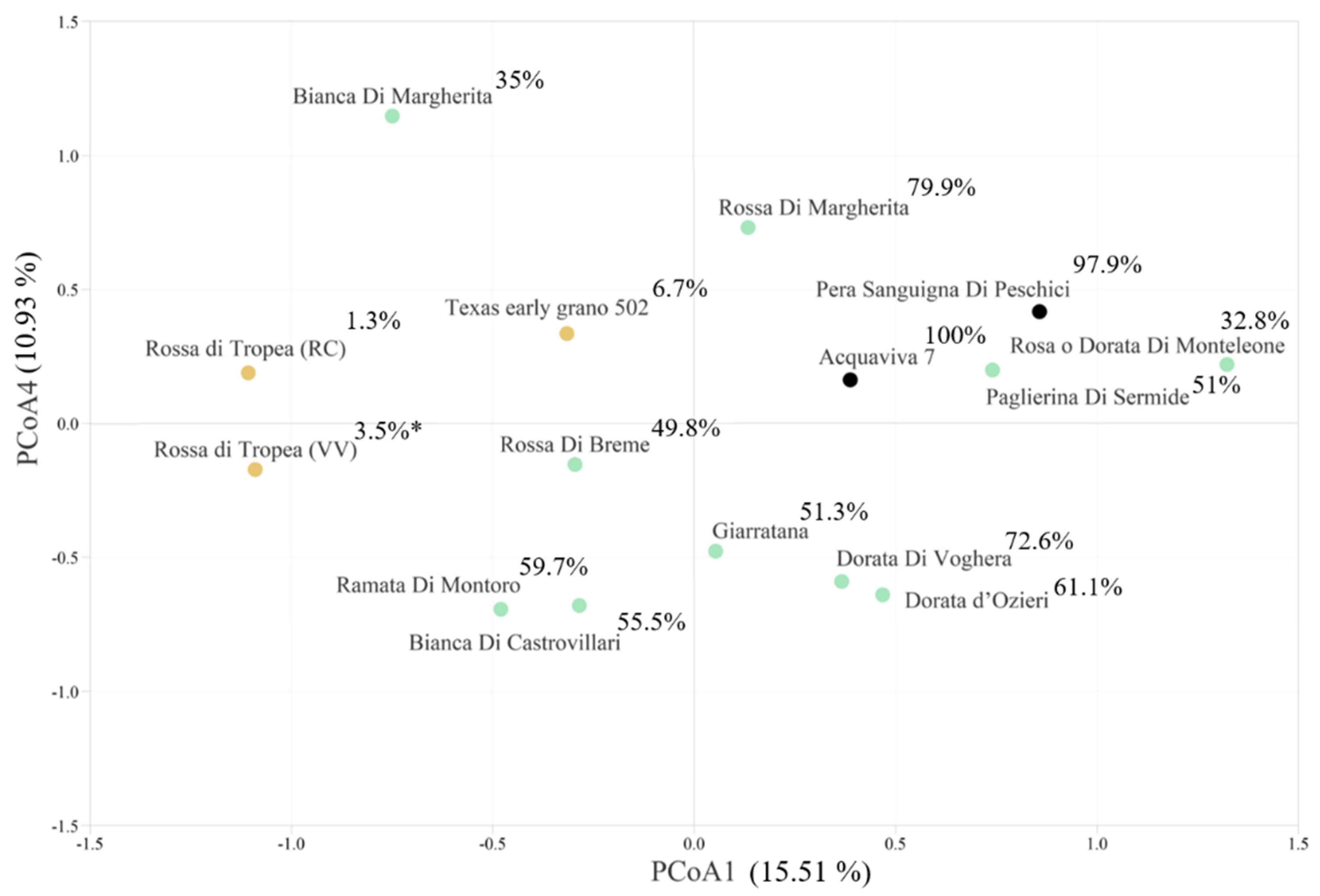
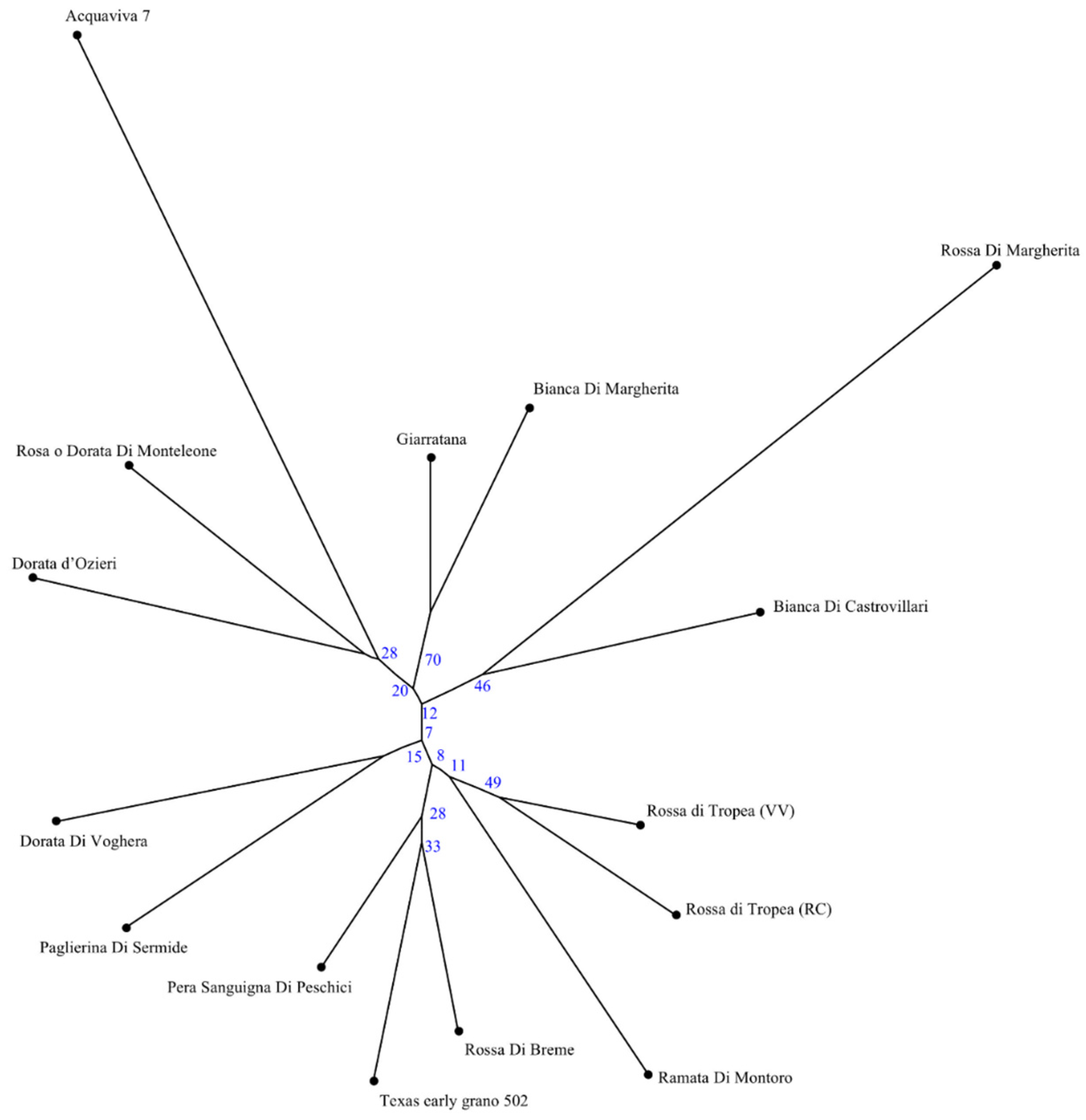
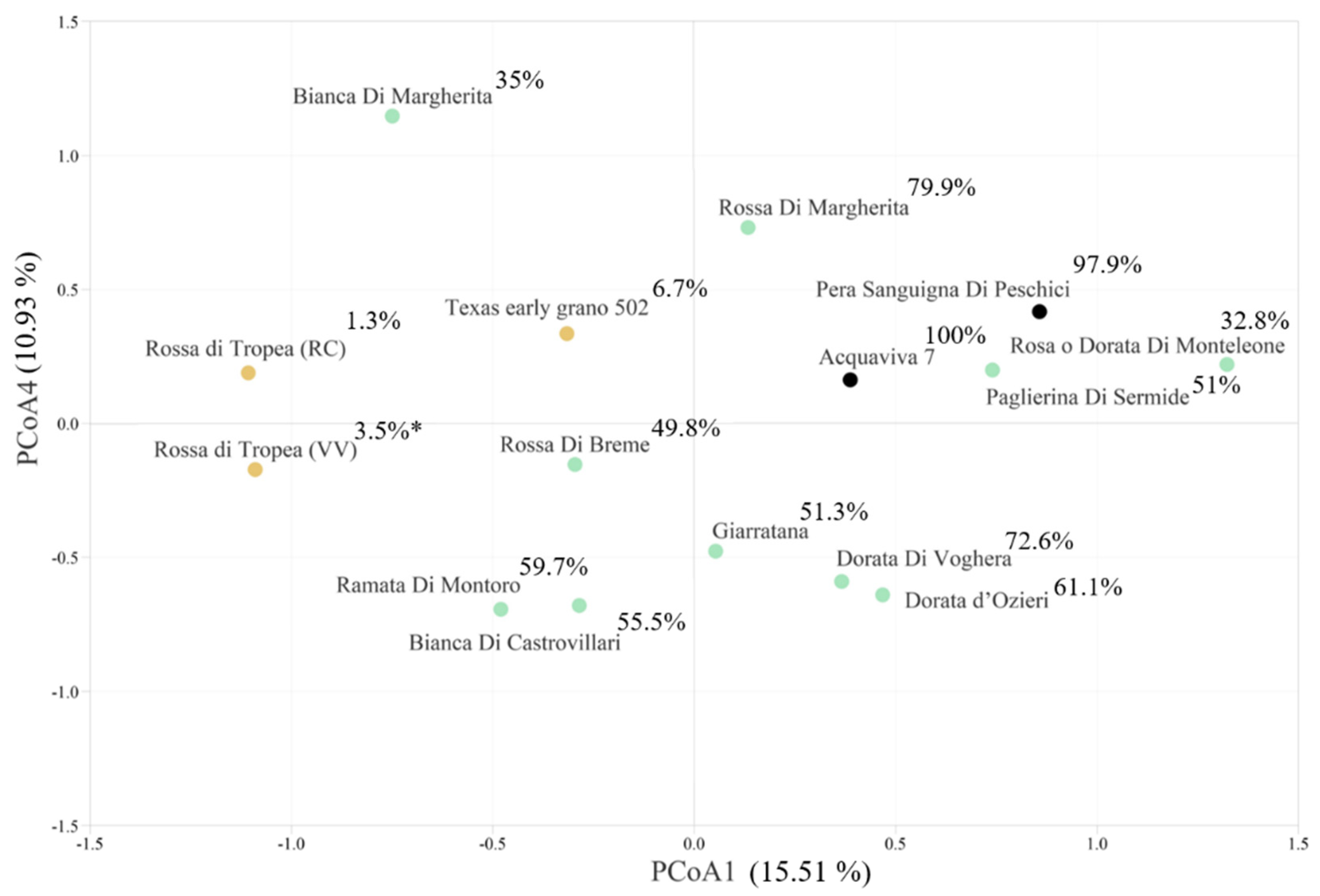
3.2. SSR Genetic Analysis
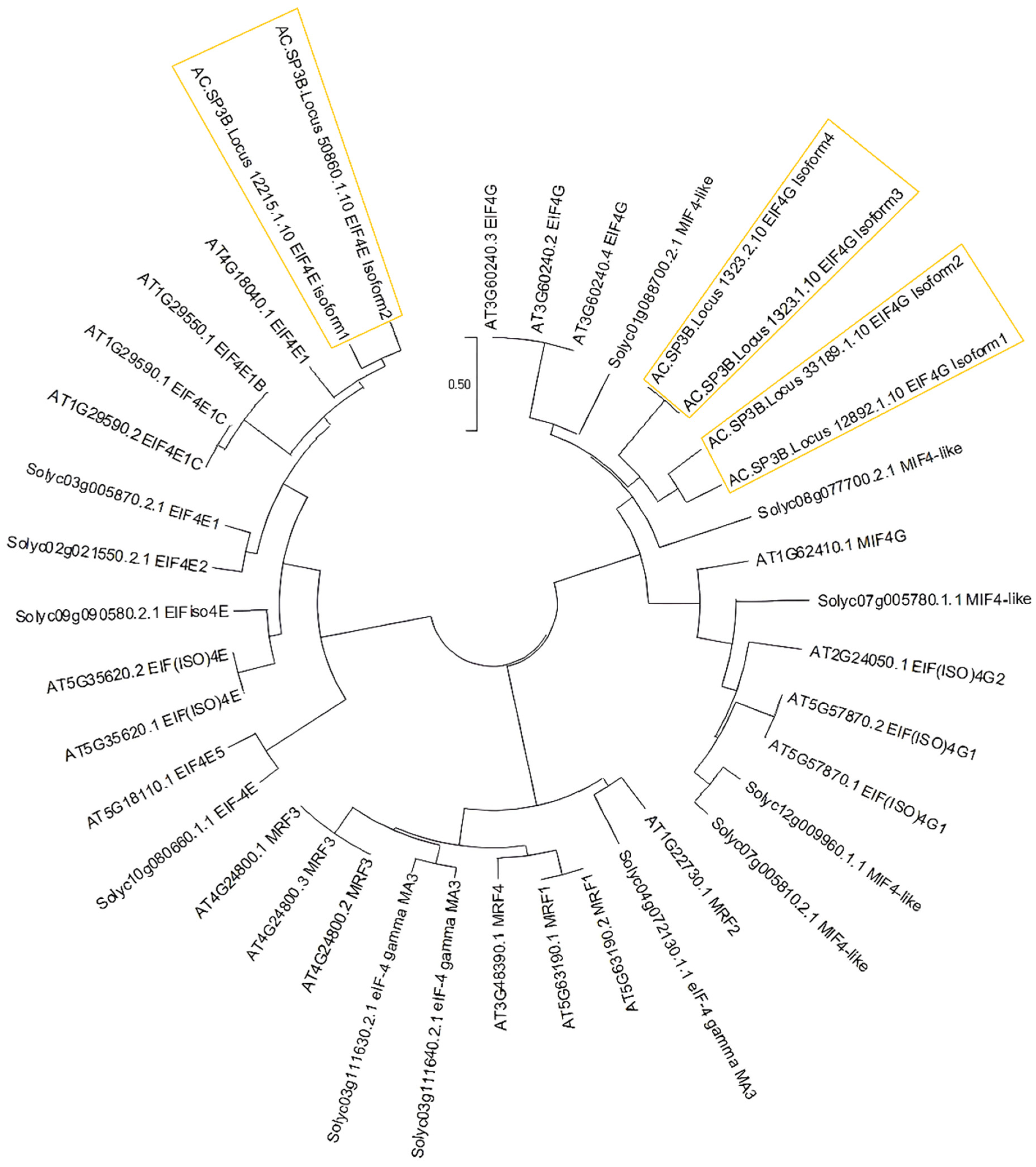
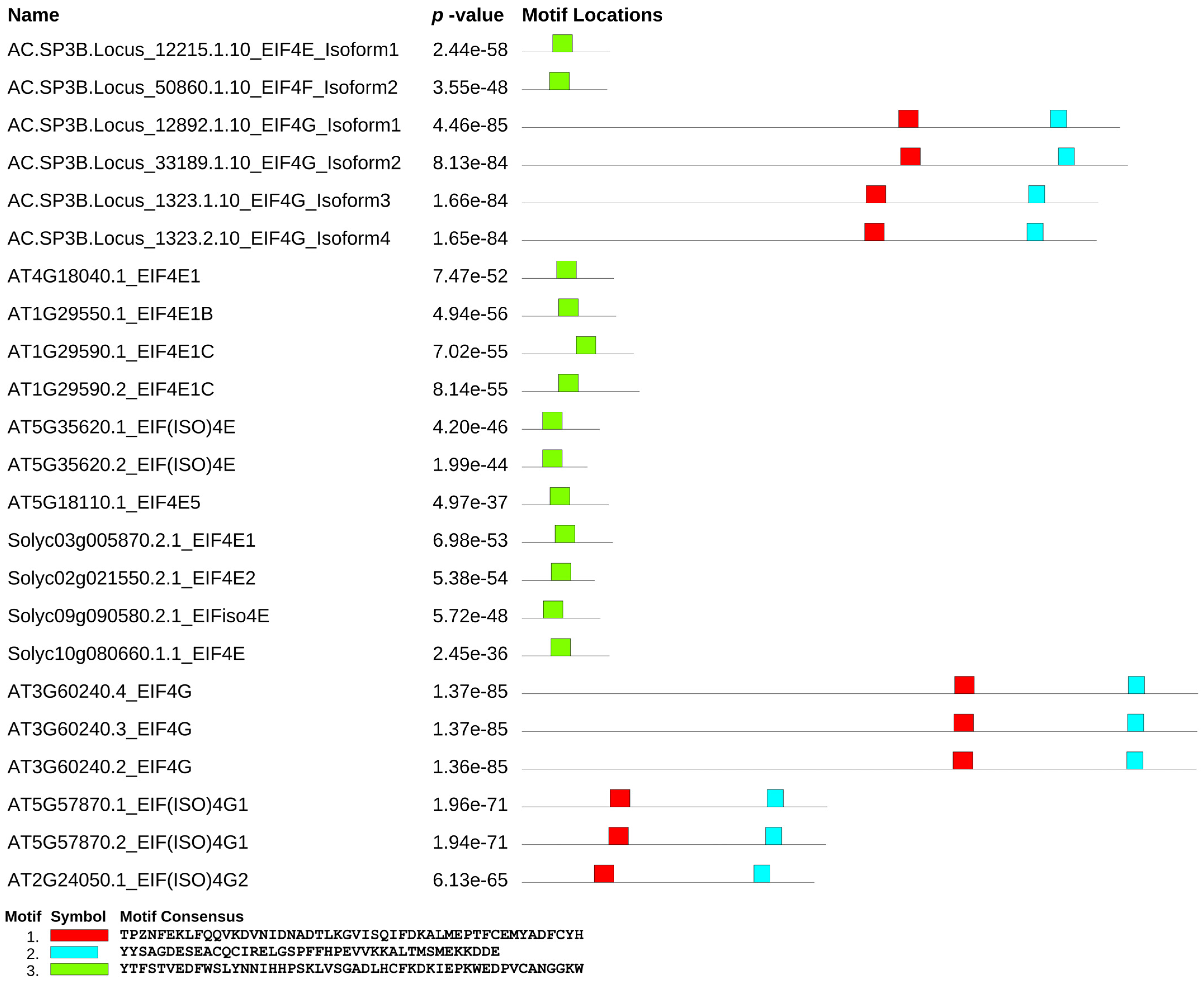
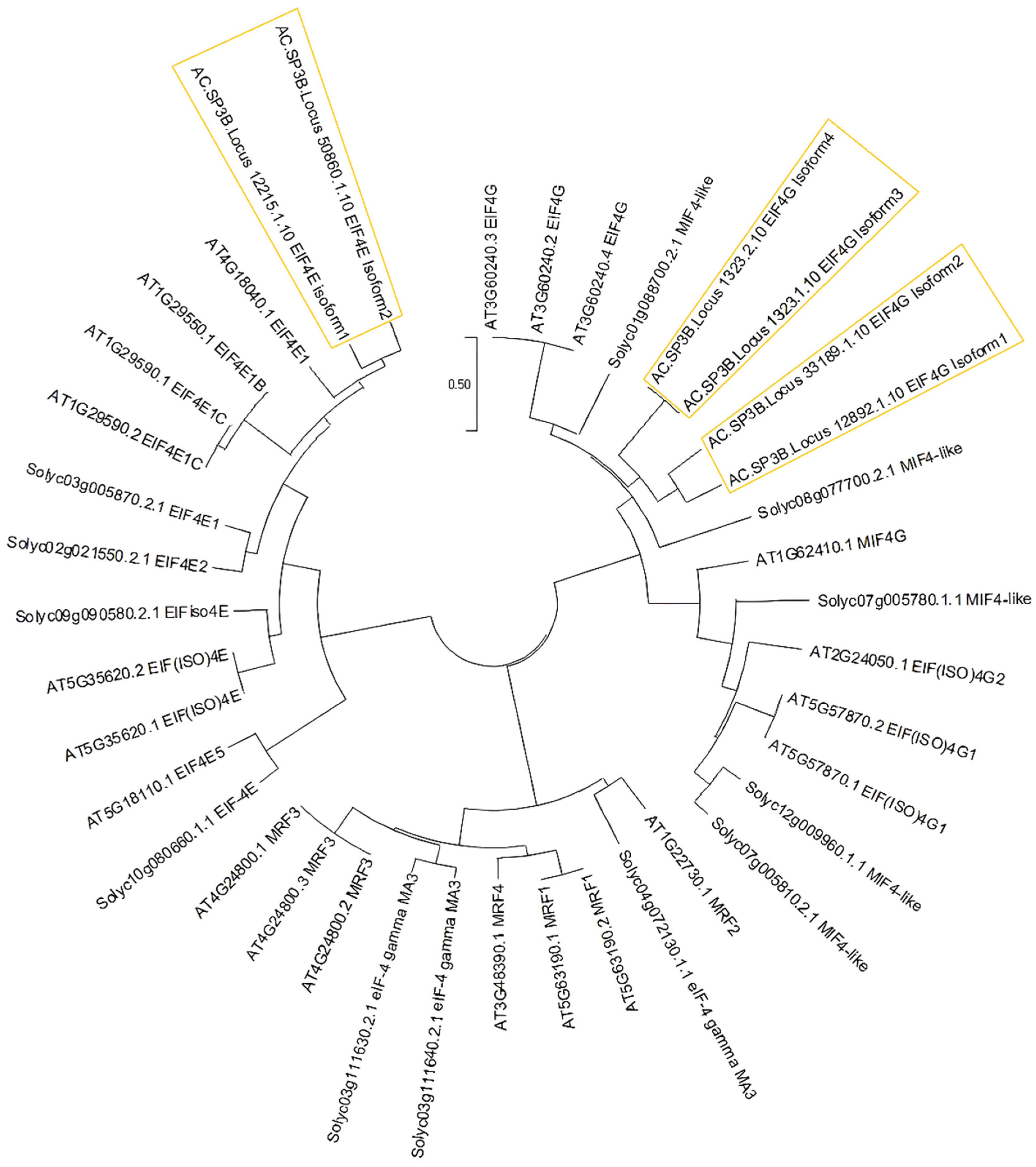
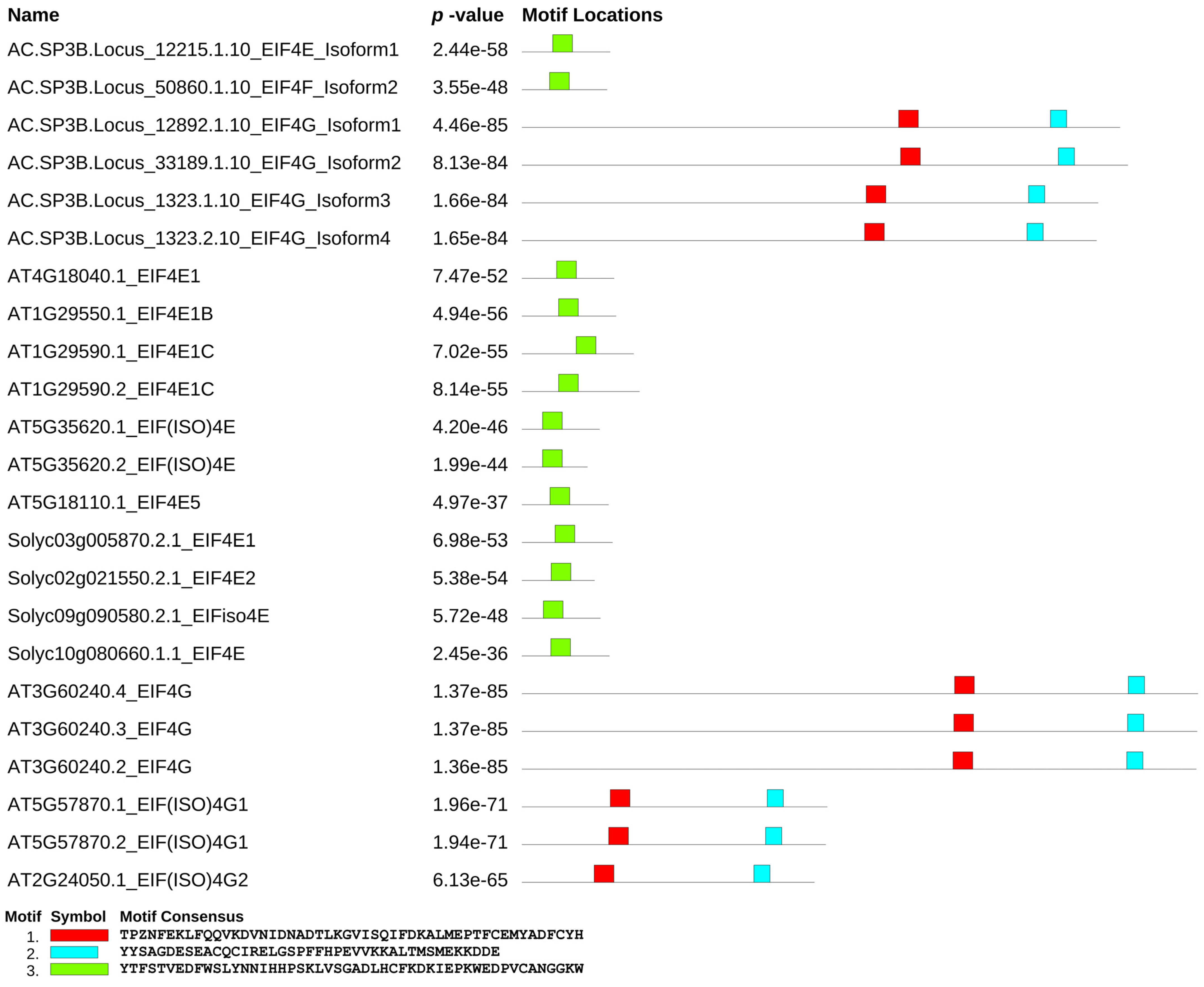
3.3. Conserved Motifs among EIF4e and EIF4g Interspecies
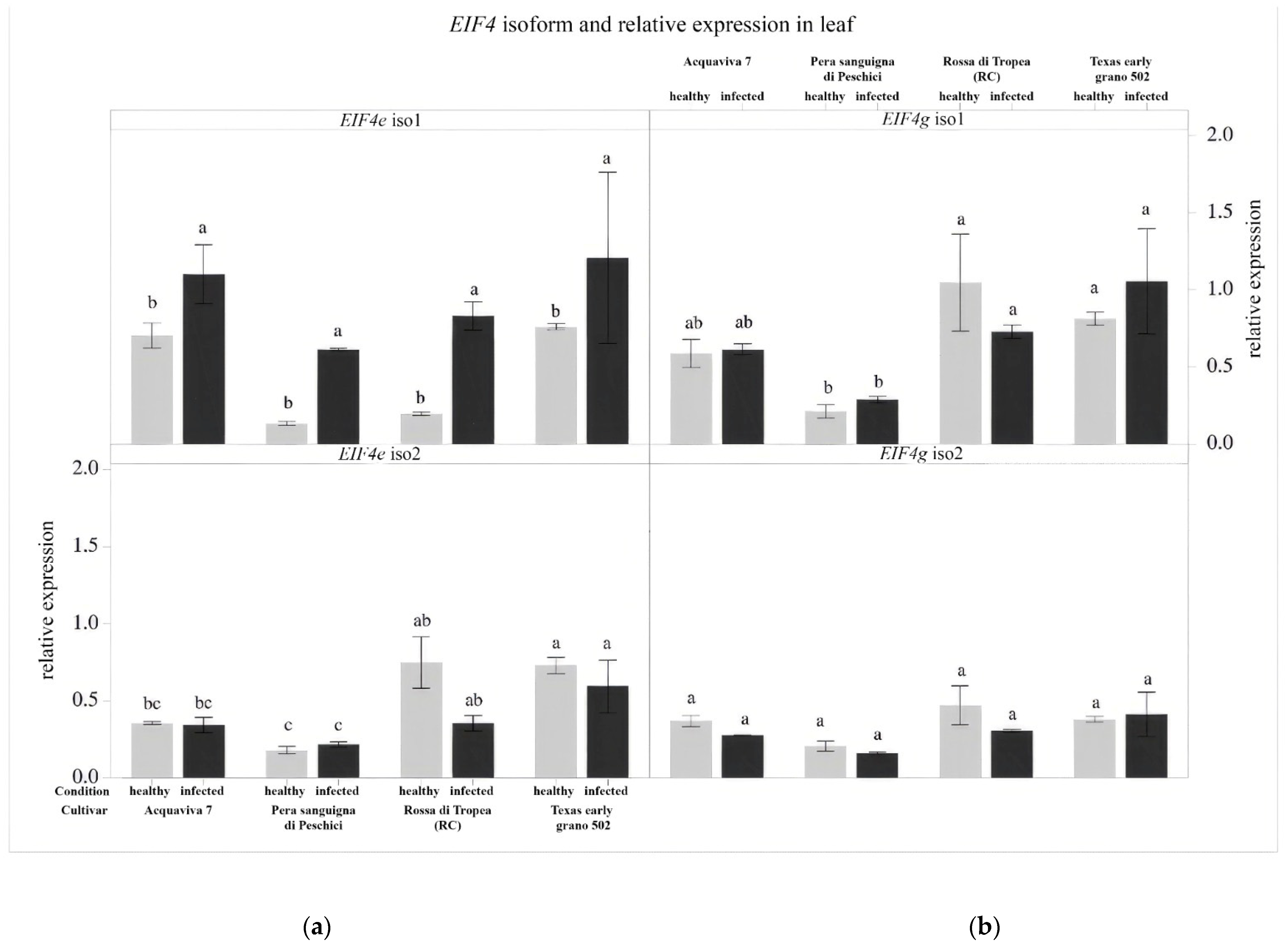
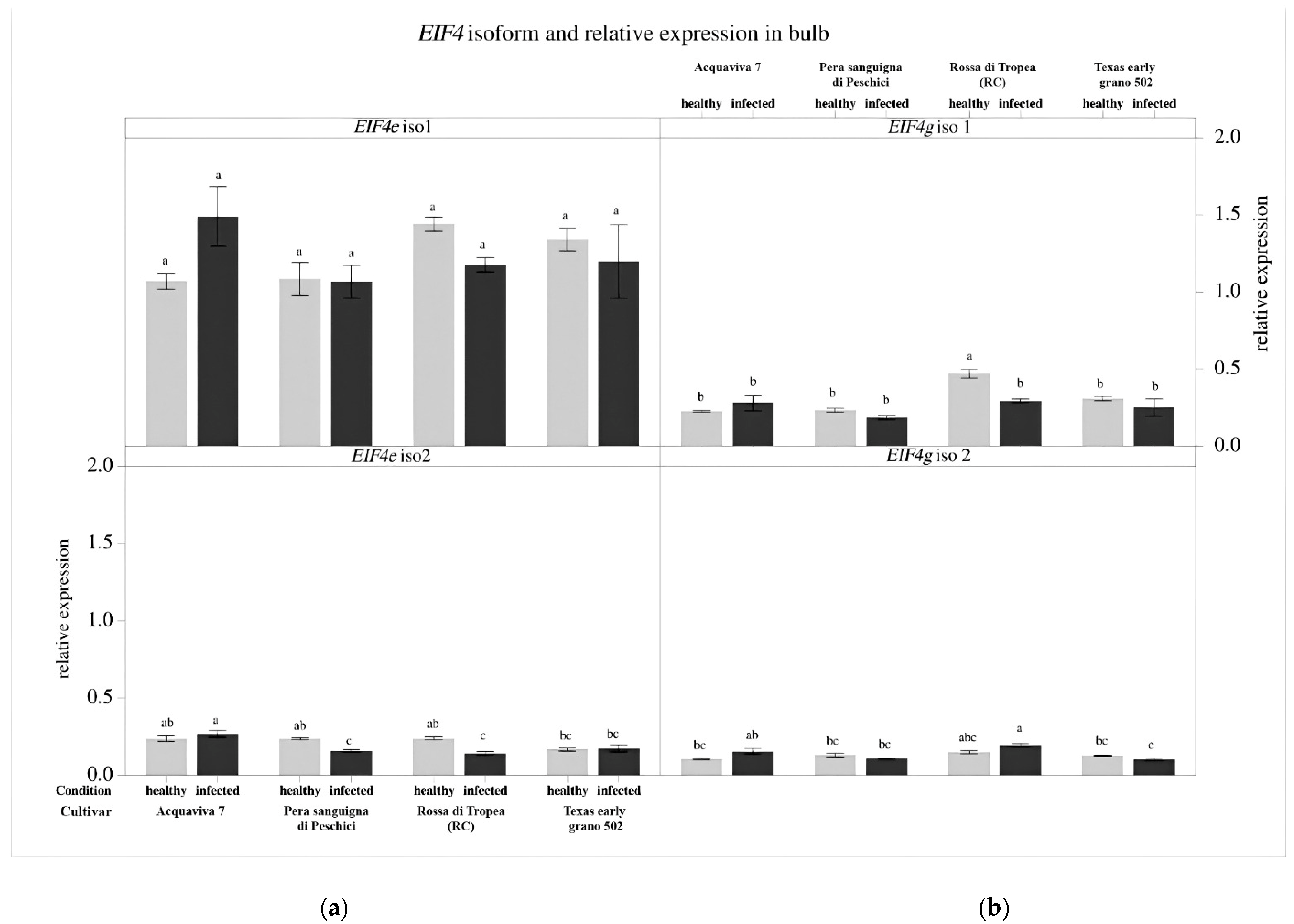
3.4. Gene Expression Analysis of the Eukaryotic Translation Initiation Factors (EIF)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, M.; Barbhai, M.D.; Hasan, M.; Dhumal, S.; Singh, S.; Pandiselvam, R.; Rais, N.; Natta, S.; Senapathy, M.; Sinha, N.; et al. Onion (Allium cepa L.) Peel: A Review on the Extraction of Bioactive Compounds, Its Antioxidant Potential, and Its Application as a Functional Food Ingredient. J. Food Sci. 2022, 87, 4289–4311. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, A.J.; Ohshima, K.; Phillips, M.J.; Gibbs, M.J. The Prehistory of Potyviruses: Their Initial Radiation Was during the Dawn of Agriculture. PLoS ONE 2008, 3, e2523. [Google Scholar] [CrossRef] [PubMed]
- Melhus, I.; Reddy, C.; Shenderson, W.J.; Vestal, E. A New Virus Disease Epidemic on Onions. Phytopathology 1929, 19, 73–77. [Google Scholar]
- Mariani, F.; Bertaccini, A. Le Virosi delle Piante Ortive; Reda edizioni per L’agricoltura: Rome, Italy, 1983; pp. 104–111. [Google Scholar]
- Parrella, G.; De Stradis, A.; Volvas, C.; Agosteo, G.E. Outbreaks of Onion Yellow Dwarf Virus (OYDV) on Onion Crops in Calabria (Southern Italy). J. Plant Pathol. 2005, 87, 302. [Google Scholar]
- Kumar, P.; Dhawan, P.; Mehra, R. Symptoms and Losses Caused by Onion Yellow Dwarf Virus and Iris Yellow Spot Virus Diseases of Onion Crop in Northern India. J. Mycol. Plant Pathol. 2012, 42, 153–160. [Google Scholar]
- Elnagar, S.; Abdel-Kader El-Sheikh, M.; Salah El-Deen Abd El-Wahab, A. Effect of Natural Infection with Onion Yellow Dwarf Virus (OYDV) on Yield of Onion and Garlic Crops in Egypt. J. Life Sci. 2011, 5, 634–638. [Google Scholar]
- Drake, C.J.; Tate, H.D.; Harris, H.M. The Relationship of Aphids to the Transmission of Yellow Dwarf of Onion. J. Econ. Entomol. 1933, 26, 841–846. [Google Scholar] [CrossRef]
- Kumar, P.; Dhawan, P.; Mehra, R. Characterization, Transmission and Host Range of Onion Yellow Dwarf Virus. Plant Dis. Res. 2011, 26, 176. [Google Scholar]
- Manglli, A.; Tomassoli, L.; Tiberini, A.; Agosteo, G.E.; Fontana, A.; Pappu, H.R.; Albanese, G. A Survey on the Infection of Onion Yellow Dwarf Virus and Iris Yellow Spot Tospovirus in Seed and Bulb Productions Systems of Onion in Calabria, Italy. Eur. J. Plant Pathol. 2020, 156, 767–778. [Google Scholar] [CrossRef]
- Ibrahim, L.M.; Award, M.A.E.; Abou-Zeid, A.A.; Gamal-Elin, A.S. Isolation and Identification of Onion Yellow Dwarf Virus in Egypt. J. Appl. Sci. 1996, 11, 184–196. [Google Scholar]
- Abd El-Wahab, A.; Elnagar, S.; Abdel Wahab, B.A.; Elnagar, S.; El-Sheikh, M. Incidence of aphid-borne onion yellow dwarf virus (OYDV) in Alliaceae crops and associated weeds in Egypt. In Proceedings of the 4th Conference on Recent Technologies in Agriculture, Giza, Egypt, 3–5 November 2009. [Google Scholar]
- Wang, A.; Krishnaswamy, S. Eukaryotic Translation Initiation Factor 4E-Mediated Recessive Resistance to Plant Viruses and Its Utility in Crop Improvement. Mol. Plant Pathol. 2012, 13, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Sanfaçon, H. Plant Translation Factors and Virus Resistance. Viruses 2015, 7, 3392–3419. [Google Scholar] [CrossRef]
- Moury, B.; Charron, C.; Janzac, B.; Simon, V.; Gallois, J.L.; Palloix, A.; Caranta, C. Evolution of Plant Eukaryotic Initiation Factor 4E (EIF4E) and Potyvirus Genome-Linked Protein (VPg): A Game of Mirrors Impacting Resistance Spectrum and Durability. Infect. Genet. Evol. 2014, 27, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Nicaise, V.; German-Retana, S.; Sanjuán, R.; Dubrana, M.P.; Mazier, M.; Maisonneuve, B.; Candresse, T.; Caranta, C.; LeGall, O. The Eukaryotic Translation Initiation Factor 4E Controls Lettuce Susceptibility to the Potyvirus Lettuce Mosaic Virus. Plant Physiol. 2003, 132, 1272–1282. [Google Scholar] [CrossRef] [PubMed]
- Michel, V.; Julio, E.; Candresse, T.; Cotucheau, J.; Decorps, C.; Volpatti, R.; Moury, B.; Glais, L.; Jacquot, E.; de Borne, F.D.; et al. A Complex EIF4E Locus Impacts the Durability of va Resistance to Potato Virus Y in Tobacco. Mol. Plant Pathol. 2019, 20, 1051–1066. [Google Scholar] [CrossRef] [PubMed]
- Ling, K.S.; Harris, K.R.; Meyer, J.D.F.; Levi, A.; Guner, N.; Wehner, T.C.; Bendahmane, A.; Havey, M.J. Non-Synonymous Single Nucleotide Polymorphisms in the Watermelon EIF4E Gene Are Closely Associated with Resistance to Zucchini Yellow Mosaic Virus. Theor. Appl. Genet. 2009, 120, 191–200. [Google Scholar] [CrossRef]
- Zlobin, N.; Taranov, V. Plant EIF4E Isoforms as Factors of Susceptibility and Resistance to Potyviruses. Front. Plant Sci. 2023, 14, 1041868. [Google Scholar] [CrossRef]
- Taglienti, A.; Tiberini, A.; Ciampa, A.; Piscopo, A.; Zappia, A.; Tomassoli, L.; Poiana, M.; Dell’Abate, M.T. Metabolites Response to Onion Yellow Dwarf Virus (OYDV) Infection in ‘Rossa Di Tropea’ Onion during Storage: A 1H HR-MAS NMR Study. J. Sci. Food Agric. 2020, 100, 3418–3427. [Google Scholar] [CrossRef]
- Tiberini, A.; Mangano, R.; Micali, G.; Leo, G.; Manglli, A.; Tomassoli, L.; Albanese, G. Onion Yellow Dwarf Virus ∆∆Ct-Based Relative Quantification Obtained by Using Real-Time Polymerase Chain Reaction in ‘Rossa Di Tropea’ Onion. Eur. J. Plant Pathol. 2019, 153, 251–264. [Google Scholar] [CrossRef]
- Clark, M.F.; Adams, A.N. Characteristics of the Microplate Method of Enzyme-Linked Immunosorbent Assay for the Detection of Plant Viruses. J. Gen. Virol. 1977, 34, 475–483. [Google Scholar] [CrossRef]
- Baldwin, S.; Pither-Joyce, M.; Wright, K.; Chen, L.; McCallum, J. Development of Robust Genomic Simple Sequence Repeat Markers for Estimation of Genetic Diversity within and among Bulb Onion (Allium cepa L.) Populations. Mol. Breed. 2012, 30, 1401–1411. [Google Scholar] [CrossRef]
- McCallum, J.; Thomson, S.; Pither-Joyce, M.; Kenel, F.; Clarke, A.; Havey, M.J. Genetic Diversity Analysis and Single-Nucleotide Polymorphism Marker Development in Cultivated Bulb Onion Based on Expressed Sequence Tag–Simple Sequence Repeat Markers. J. Am. Soc. Hortic. Sci. 2008, 133, 810–818. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenALEx 6.5: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research-an Update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Nei, M.; Li, W.-H. Mathematical Model for Studying Genetic Variation in Terms of Restriction Endonucleases. Mol. Evol./Mitochondrial DNA/Nucleotide Divers. 1979, 76, 5269–5273. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME Suite: Tools for Motif Discovery and Searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, H.; Zhan, Z.; Liu, B.; Chen, Z.; Liang, Y. Transcriptome Analysis of Sucrose Metabolism during Bulb Swelling and Development in Onion (Allium cepa L.). Front. Plant Sci. 2016, 7, 1425. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Poque, S.; Wu, H.W.; Huang, C.H.; Cheng, H.W.; Hu, W.C.; Yang, J.Y.; Wang, D.; Yeh, S.D. Potyviral Gene-Silencing Suppressor HCPro Interacts with Salicylic Acid (SA)-Binding Protein 3 to Weaken SA-Mediated Defense Responses. Mol. Plant-Microbe Interact. 2018, 31, 86–100. [Google Scholar] [CrossRef]
- Nyalugwe, E.P.; Barbetti, M.J.; Clode, P.L.; Jones, R.A.C. Programmed Cell Death Pathways Induced by Early Plant-Virus Infection Are Determined by Isolate Virulence and Stage of Infection. Plant Pathol. 2016, 65, 1518–1528. [Google Scholar] [CrossRef]
- Taglienti, A.; Dell’Abate, M.T.; Ciampa, A.; Tomassoli, L.; Albanese, G.; Sironi, L.; Tiberini, A. Study on Ultra-Structural Effects Caused by Onion Yellow Dwarf Virus Infection in ‘Rossa Di Tropea’ Onion Bulb by Means of Magnetic Resonance Imaging. Sci. Hortic. 2020, 271, 109486. [Google Scholar] [CrossRef]
- Murray, R.R.; Emblow, M.S.M.; Hetherington, A.M.; Foster, G.D. Plant Virus Infections Control Stomatal Development. Sci. Rep. 2016, 6, 34507. [Google Scholar] [CrossRef] [PubMed]
- Liu, G. Food Losses and Food Waste in China: A First Estimate; OECD Publishing: Paris, France, 2014. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Ntatsi, G.; Ferreira, I.C.F.R. Long-Term Storage of Onion and the Factors That Affect Its Quality: A Critical Review. Food Rev. Int. 2017, 33, 62–83. [Google Scholar] [CrossRef]
- Gubb, I.R.; MacTavish, H.S. Onion Pre- and Post-Harvest Considerations. In Allium Crop Science: Recent Advances; Rabinowitch, H.D., Currah, L., Eds.; CABI Publishing: Wallingford, UK, 2002; ISBN 9780851995106. [Google Scholar]
- Ricciardi, L.; Mazzeo, R.; Marcotrigiano, A.R.; Rainaldi, G.; Iovieno, P.; Zonno, V.; Pavan, S.; Lotti, C. Assessment of Genetic Diversity of the “Acquaviva Red Onion” (Allium cepa L.) Apulian Landrace. Plants 2020, 9, 260. [Google Scholar] [CrossRef] [PubMed]
- Raj, A.C.; Sharangi, A.B.; Das, A.; Pramanik, K.; Upadhyay, T.K.; Almutairi, M.; Khan, M.I.; Ahmad, I.; Kausar, M.A.; Saeed, M. Assessing the Genetic Divergence of Onion (Allium cepa L.) through Morpho-Physiological and Molecular Markers. Sustainability 2022, 14, 1131. [Google Scholar] [CrossRef]
- Joshi, B.; Lee, K.; Maeder, D.L.; Jagus, R. Phylogenetic Analysis of EIF4E-Family Members. BMC Evol. Biol. 2005, 5, 48. [Google Scholar] [CrossRef]
- Patrick, R.M.; Lee, J.C.H.; Teetsel, J.R.J.; Yang, S.H.; Choy, G.S.; Browning, K.S. Discovery and Characterization of Conserved Binding of EIF4E 1 (CBE1), a Eukaryotic Translation Initiation Factor 4E– Binding Plant Protein. J. Biol. Chem. 2018, 293, 17240–17247. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accessions | Germplasm Collection | Origin |
|---|---|---|
| Rossa di Tropea (Reggio Calabria-RC) | University of Reggio Calabria | Calabria |
| Giarratana | Private nursery | Sicilia |
| Bianca di Castrovillari | ARSAC–Castrovillari (Cosenza-CS) | Calabria |
| Pera Sanguigna di Peschici | University of Foggia | Puglia |
| Rossa Margherita | University of Foggia | Puglia |
| Bianca di Margherita | University of Foggia | Puglia |
| Rosa o Dorata di Monteleone | University of Foggia | Puglia |
| Acquaviva (1–12 *) | University of Bari | Puglia |
| Ramata di Montoro | Coop.Rama | Campania |
| Dorata d’Ozieri | Genebank information system of the IPK Gatersleben | Sardegna |
| Dorata di Voghera | University of Pavia | Lombardia |
| Rossa di Breme | University of Pavia | Lombardia |
| Paglierina di Sermide | University of Pavia | Lombardia |
| Texas early grano 502 | Private seller | Commercial cultivar |
| Primer | Forward | Reverse | Size | Dye |
|---|---|---|---|---|
| ACM 373 | AGGTTAAGAAGTTGAATGGTCTG | AAATGGACAAGTGGCATTCA | 145–159 | FAM |
| ACM 101 | CCTTTGCTAACCAAATCCGA | CTTGTTGAGAAGGAGGACGC | 227–248 | VIC |
| ACM 235 | TGAGTCGGCACTCACCTATG | ACGCATTTTCAAA TGAAGGC | 292/304 | PET |
| ACM 446 | TCAAGAATTCTGTTGCATCTTGT | AATAAGACCGCAGAAACGAAA | 122–124 | FAM |
| ACM 449 | GTAAAGGTGTAATAGGAATGAATCG | TACAAAGAAACACACGCGCT | 133–148 | VIC |
| ACM 045 | AAAACGAAGCAACAAACAAAA | CGACGAAGGTCATAAGTAGGC | 226–275 | NED |
| ACM 138 | ACGGTTTGATGCACAAGATG | CCAACCAACAGTTGATACTGC | 242–286 | NED |
| ACM 387 | ACGCACACTATTTGGGAAGG | GAGGAATAGAGAAGGCTGCG | 151–162 | PET |
| ACM 134 | ACACACACAAGAGGGAAGGG | CACACACCCACACACATCAA | 198–212 | FAM |
| ACM 119 | TTTCAGCAACATAGTATTGCGTC | TCTTCGGGATTGGTATGGAG | 241–259 | PET |
| ACM 443 | TGGTGCTTGCTATGTTTTGC | CCCTAGGCCAAGCTTACTTGT | 154–179 | NED |
| ACM 477 | TGCAATTGGAACTTTGGTTTT | CCGTTCCTCTATTTTGCAGC | 160–165 | VIC |
| Primer | Forward | Reverse | Ta (°C) | Conc. (μmol) |
|---|---|---|---|---|
| EIF4e iso 1 | GAGGACCCTGTTTGTGCCA | GTGTGCATTTTTGGTCCAT | 59 | 0.5 |
| EIF4e iso 2 | GTCACCCAAGCAATTTAATG | GATTTTCCTCG TGAACAGTTGAC | 59 | 0.6 |
| EIF4g iso 1 | CCCTCAGTGTTGCCTTCTCC | TTCCATATCCACGCTGAGGC | 59 | 0.2 |
| EIF4g iso 2 | TGGCAGGAGAGAAGGAAGGT | GGTCCTCGTGTCAGTCTGTT | 59 | 0.5 |
| Actin | CTGGGATGACATGGAGAAGATT | GTTAAGTGGAGCCTCCGT | 59 | 0.1 |
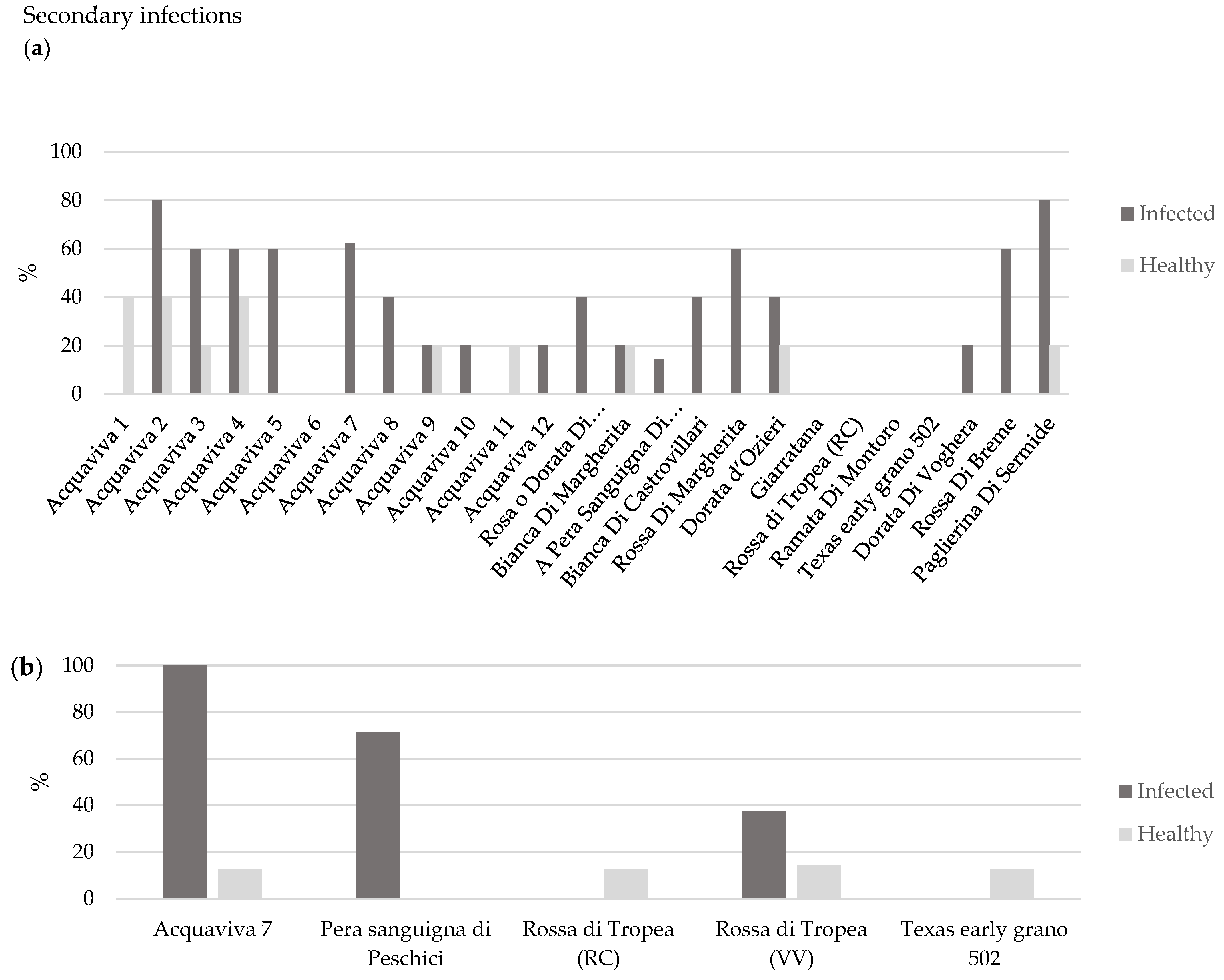
| First trial Accessions | Total Symptoms Score | OYDV-Positive Plants (n) | Infection Rating (IR %) | Plant Infection Index (PII) | PII (%) § |
|---|---|---|---|---|---|
| Acquaviva 1 | 46 | 11 | 50 | 4.18 | 43.9 |
| Acquaviva 2 | 89 | 12 | 55 | 7.42 | 77.9 |
| Acquaviva 3 | 70 | 14 | 64 | 5 | 52.5 |
| Acquaviva 4 | 107 | 14 | 64 | 7.64 | 80.2 |
| Acquaviva 5 | 104 | 16 | 73 | 6.5 | 68.2 |
| Acquaviva 6 | 55 | 14 | 64 | 3.93 | 41.2 |
| Acquaviva 7 * | 143 | 15 | 68 | 9.53 | 100 |
| Acquaviva 8 | 88 | 14 | 64 | 6.29 | 66 |
| Acquaviva 9 | 66 | 12 | 55 | 5.5 | 57.7 |
| Acquaviva 10 | 78 | 14 | 64 | 5.57 | 58.5 |
| Acquaviva 11 | 135 | 16 | 73 | 8.44 | 88.5 |
| Acquaviva 12 | 72 | 13 | 59 | 5.54 | 58.1 |
| Rosa o Dorata di Monteleone | 50 | 16 | 73 | 3.13 | 32.8 |
| Bianca di Margherita | 20 | 6 | 27 | 3.33 | 35 |
| Pera Sanguigna di Peschici * | 112 | 12 | 55 | 9.33 | 97.9 |
| Bianca di Castrovillari | 74 | 14 | 64 | 5.29 | 55.5 |
| Rossa Margherita | 99 | 13 | 59 | 7.62 | 79.9 |
| Dorata d’Ozieri | 99 | 17 | 77 | 5.82 | 61.1 |
| Giarratana | 44 | 9 | 41 | 4.89 | 51.3 |
| Rossa di Tropea (RC) * | 1 | 8 | 36 | 0.13 | 1.3 |
| Ramata di Montoro | 74 | 13 | 59 | 5.69 | 59.7 |
| Texas early grano 502 * | 8 | 13 | 59 | 0.62 | 6.7 |
| Dorata di Voghera | 90 | 13 | 59 | 6.92 | 72.6 |
| Rossa di Breme | 38 | 8 | 36 | 4.75 | 49.8 |
| Paglierina di Sermide | 68 | 14 | 64 | 4.86 | 51 |
| Second trial accessions | |||||
| Acquaviva 7 | 22 | 11 | 50 | 2 | 56 |
| Pera Sanguigna di Peschici | 50 | 14 | 64 | 3.57 | 100 |
| Rossa di Tropea (RC) | 1 | 9 | 41 | 0.11 | 3.1 |
| Texas early grano 502 | 0 | 6 | 27 | 0 | 0 |
| Rossa di Tropea (VV) | 1 | 8 | 36 | 0.13 | 3.5 |
| Locus | Na 1 | Ne 2 | I 3 | Ho 4 | He 5 | PIC 6 | F 7 |
|---|---|---|---|---|---|---|---|
| ACM373 | 13 | 9.184 | 2.378 | 0.533 | 0.922 | 0.882 | 0.401 |
| ACM101 | 7 | 3.409 | 1.455 | 0.667 | 0.731 | 0.659 | 0.057 |
| ACM235 | 5 | 2.980 | 1.284 | 0.067 | 0.687 | 0.618 | 0.900 |
| ACM446 | 5 | 3.913 | 1.460 | 0.800 | 0.770 | 0.701 | −0.075 |
| ACM449 | 7 | 4.245 | 1.620 | 0.800 | 0.791 | 0.731 | −0.047 |
| ACM045 | 7 | 4.412 | 1.681 | 0.733 | 0.800 | 0.743 | 0.052 |
| ACM138 | 12 | 6.429 | 2.115 | 0.867 | 0.874 | 0.827 | −0.026 |
| ACM387 | 10 | 4.945 | 1.871 | 0.667 | 0.825 | 0.772 | 0.164 |
| ACM134 | 9 | 5.172 | 1.875 | 0.800 | 0.834 | 0.783 | 0.008 |
| ACM119 | 7 | 3.846 | 1.602 | 0.200 | 0.766 | 0.710 | 0.730 |
| ACM443 | 9 | 4.592 | 1.802 | 0.533 | 0.809 | 0.756 | 0.318 |
| ACM477 | 8 | 5.921 | 1.892 | 0.800 | 0.860 | 0.809 | 0.037 |
| Mean | 9.25 | 4.921 | 1.753 | 0.622 | 0.805 | 0.749 | 0.209 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corrado, C.L.; Micali, G.; Mauceri, A.; Bertin, S.; Sunseri, F.; Abenavoli, M.R.; Tiberini, A. Study on Italian Onion Cultivars/Ecotypes towards Onion Yellow Dwarf Virus (OYDV) Infection. Horticulturae 2024, 10, 73. https://doi.org/10.3390/horticulturae10010073
Corrado CL, Micali G, Mauceri A, Bertin S, Sunseri F, Abenavoli MR, Tiberini A. Study on Italian Onion Cultivars/Ecotypes towards Onion Yellow Dwarf Virus (OYDV) Infection. Horticulturae. 2024; 10(1):73. https://doi.org/10.3390/horticulturae10010073
Chicago/Turabian StyleCorrado, Carla Libia, Giuseppe Micali, Antonio Mauceri, Sabrina Bertin, Francesco Sunseri, Maria Rosa Abenavoli, and Antonio Tiberini. 2024. "Study on Italian Onion Cultivars/Ecotypes towards Onion Yellow Dwarf Virus (OYDV) Infection" Horticulturae 10, no. 1: 73. https://doi.org/10.3390/horticulturae10010073
APA StyleCorrado, C. L., Micali, G., Mauceri, A., Bertin, S., Sunseri, F., Abenavoli, M. R., & Tiberini, A. (2024). Study on Italian Onion Cultivars/Ecotypes towards Onion Yellow Dwarf Virus (OYDV) Infection. Horticulturae, 10(1), 73. https://doi.org/10.3390/horticulturae10010073