
Invertase Plays a Vital Role in Regulating Wound-Induced Accumulation of Phenolic Compounds in Carrot Tissue


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatment
- (1)
- Whole: carrots with uniform color and shape and without mechanical injuries were selected, washed and air dried.
- (2)
- CK: the carrot cubes were immersed in the distilled water for 3 min and then air dried.
- (3)
- INV: the carrot cubes were immersed in the 0.5 g L−1 INV solution for 3 min and then air dried.
- (4)
- ConA: the carrot cubes were immersed in the 0.2 g L−1 ConA solution for 3 min and then air dried for 15 min.
- (5)
- IAM: the carrot cubes were immersed in the 1 mmol L−1 IAM solution for 3 min and then air dried.
- (6)
- AgNO3: the carrot cubes were immersed in the 5 mmol L−1 AgNO3 solution for 3 min and then air dried.
2.2. Total Soluble Phenolics (TSP) Contents
2.3. Antioxidant Capacity Analysis
2.4. Measurement of Soluble Sugars Content
2.5. AI and NI Activities
2.6. Gene Expression (Quantitative Real-Time PCR, qRT-PCR)
2.7. Statistical Analysis
3. Results
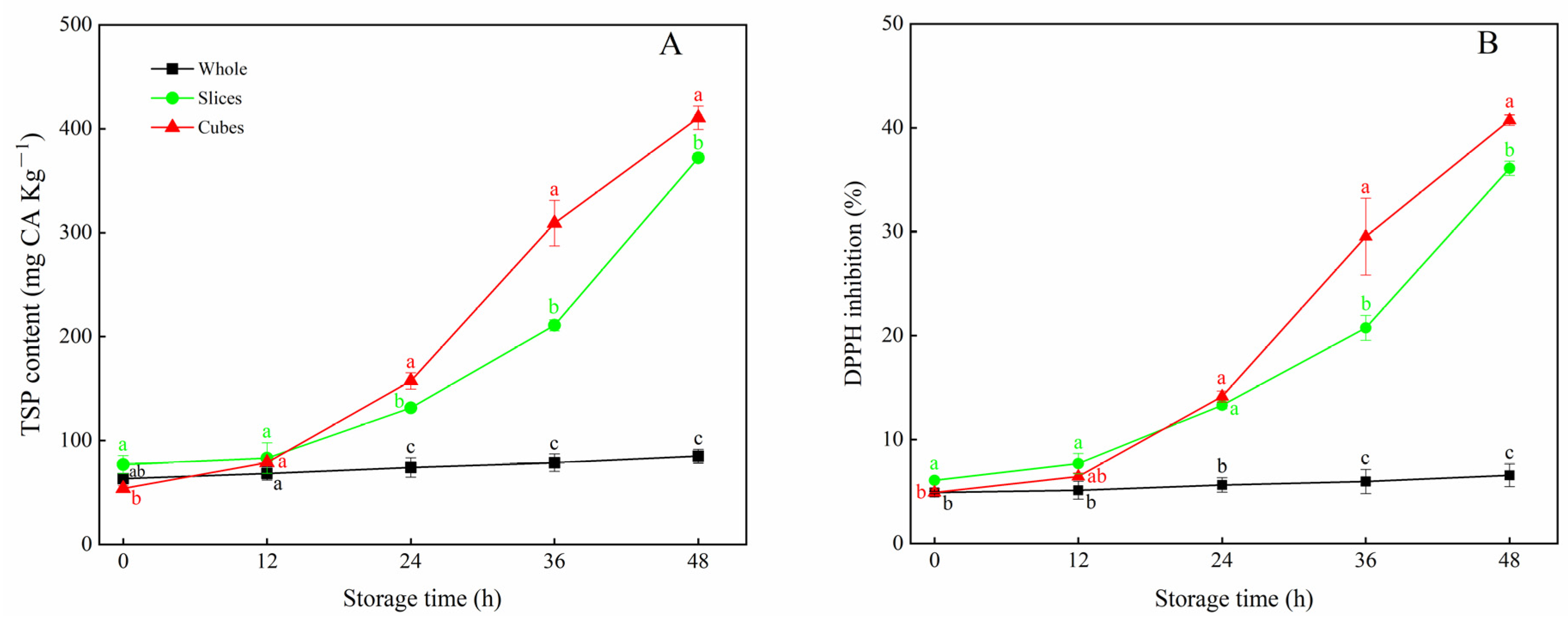
3.1. Effects of Different Cutting Styles on TSP Content and DPPH Inhibition of Carrots
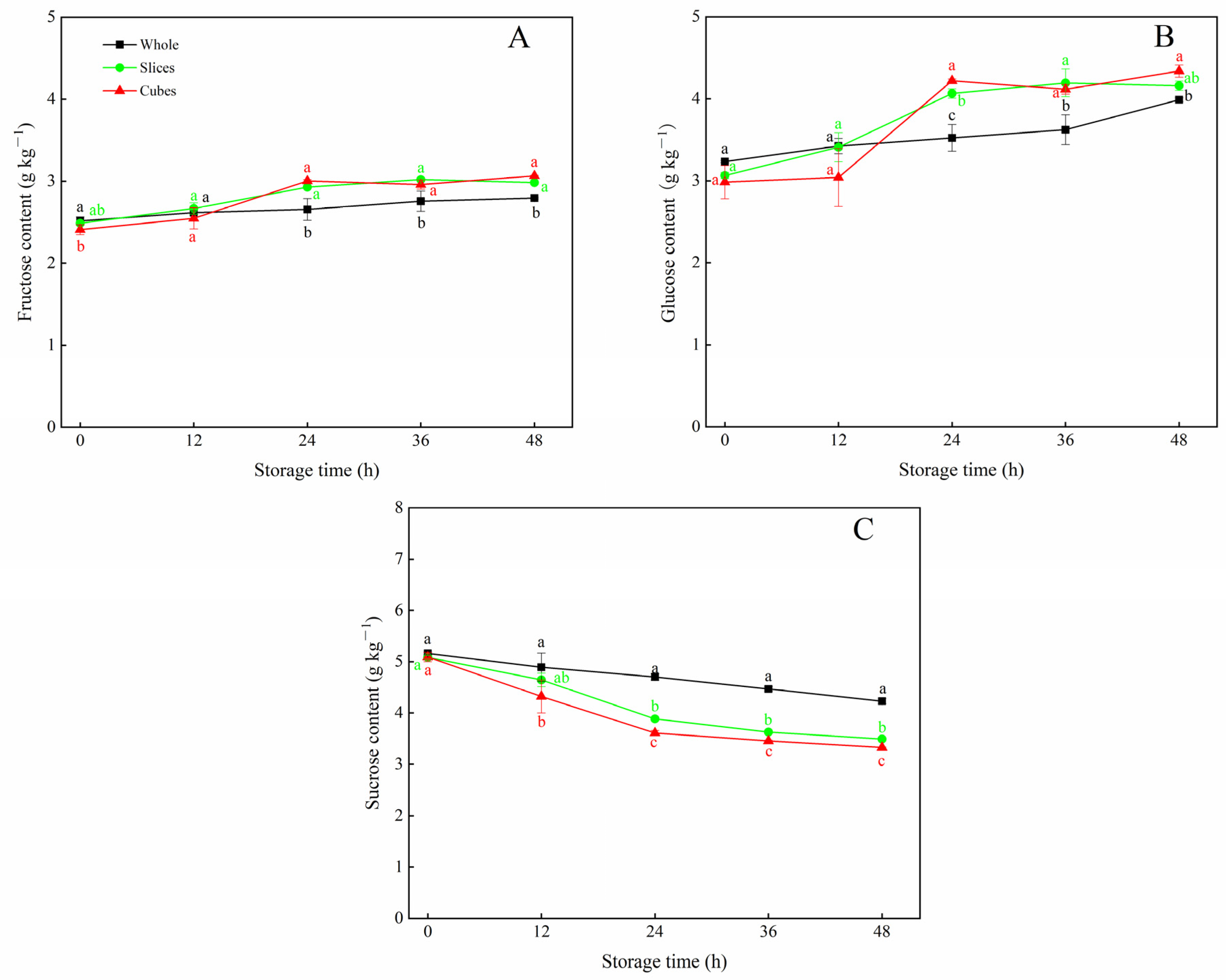
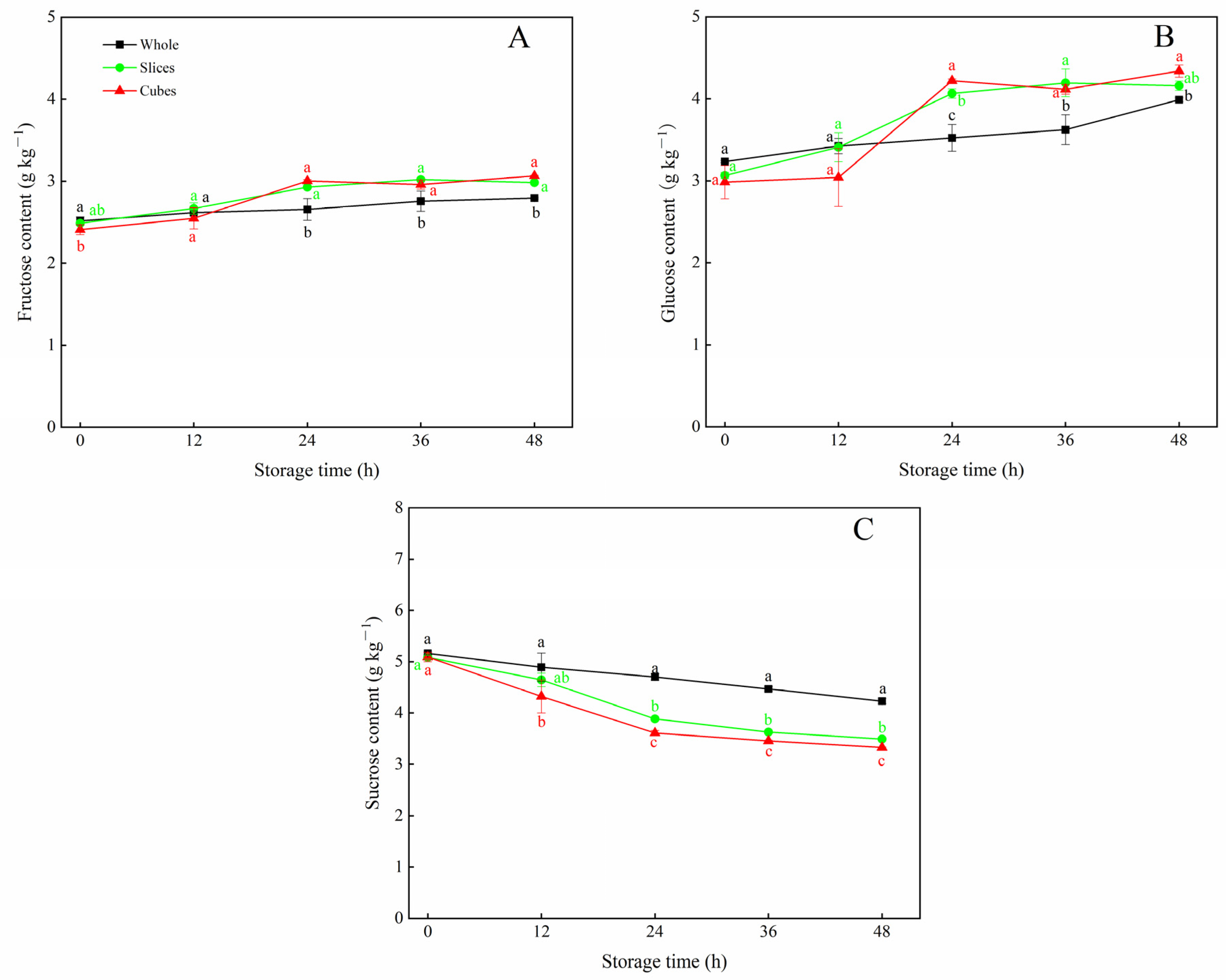
3.2. Effects of Different Cutting Styles on Soluble Sugar Contents of Carrots
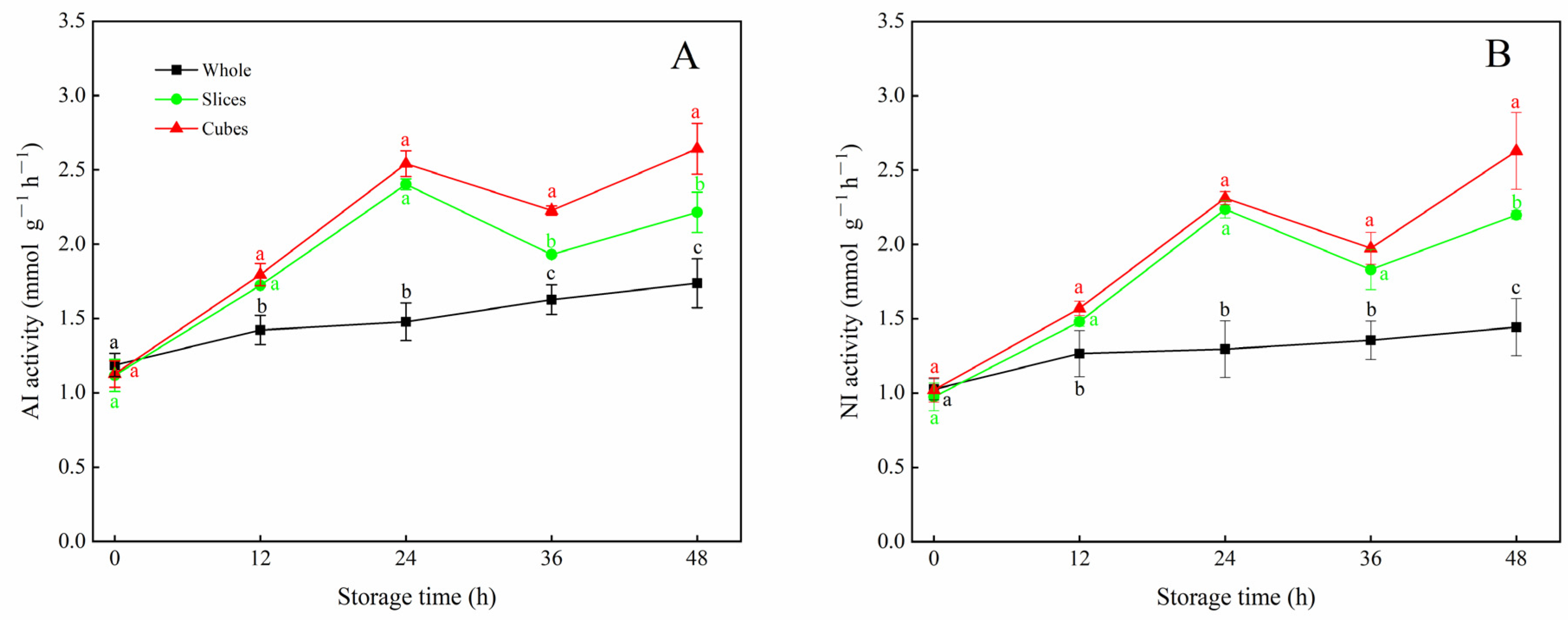
3.3. Effects of Different Cutting Styles on AI and NI Activities of Carrots
3.4. Effects of Different Cutting Styles on Relative Expression Levels of the Genes Related to INV in Carrots
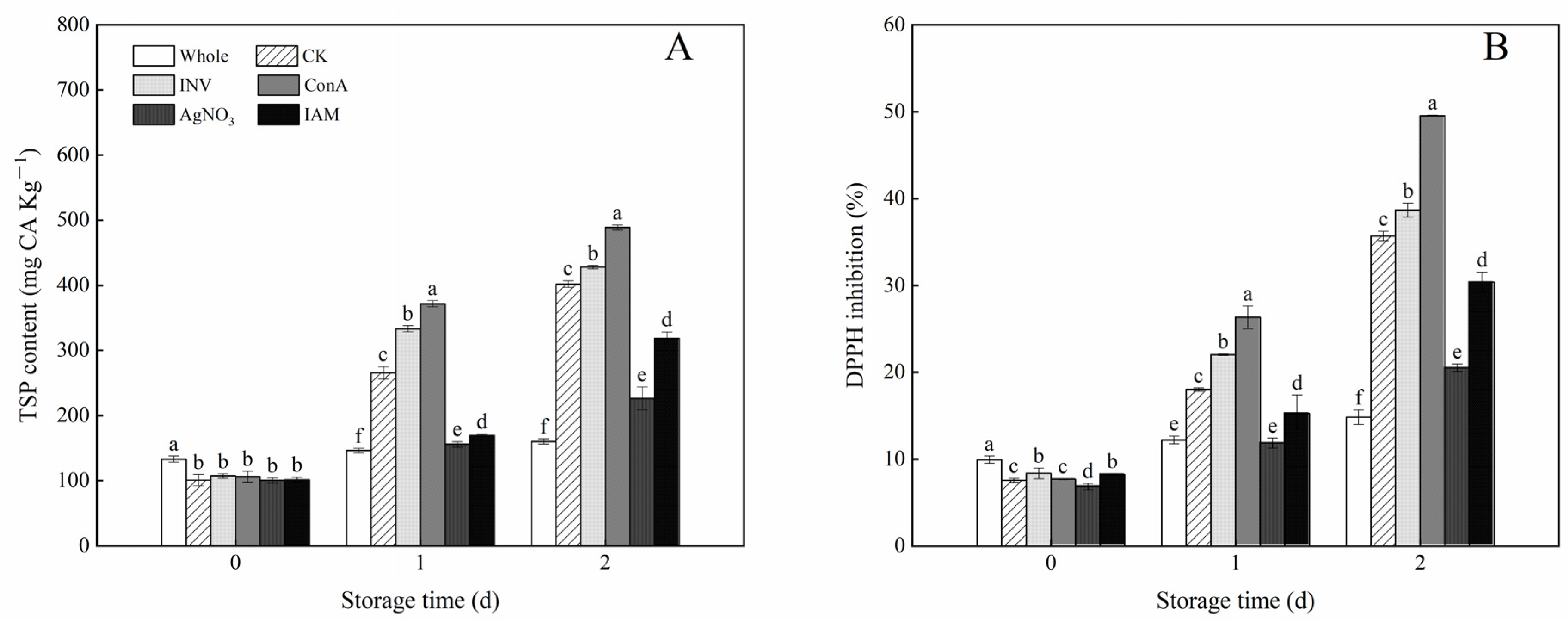
3.5. Effects of INV Activators and Inhibitors on TSP Content and DPPH Inhibition of Cubed Carrots
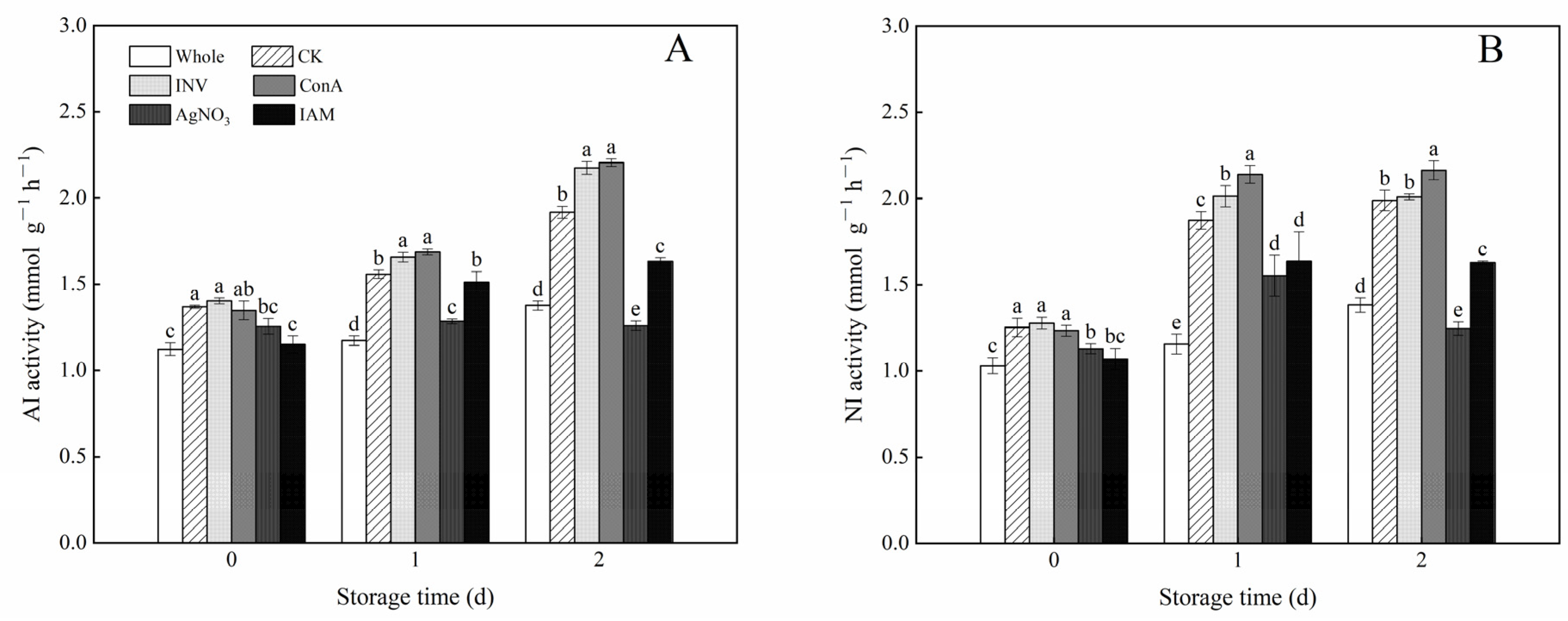
3.6. Effects of INV Activators and Inhibitors on AI and NI Activities of Cubed Carrots
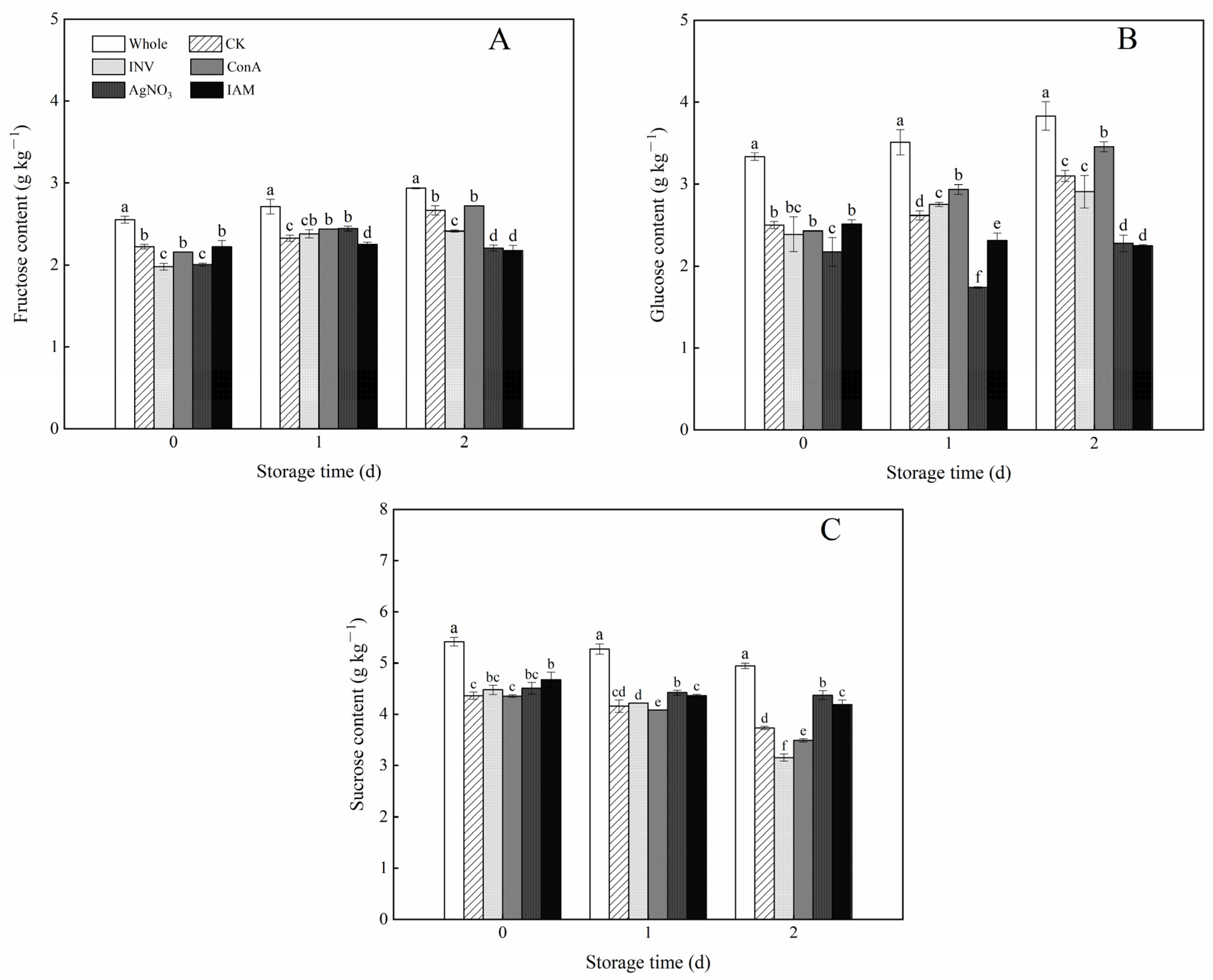
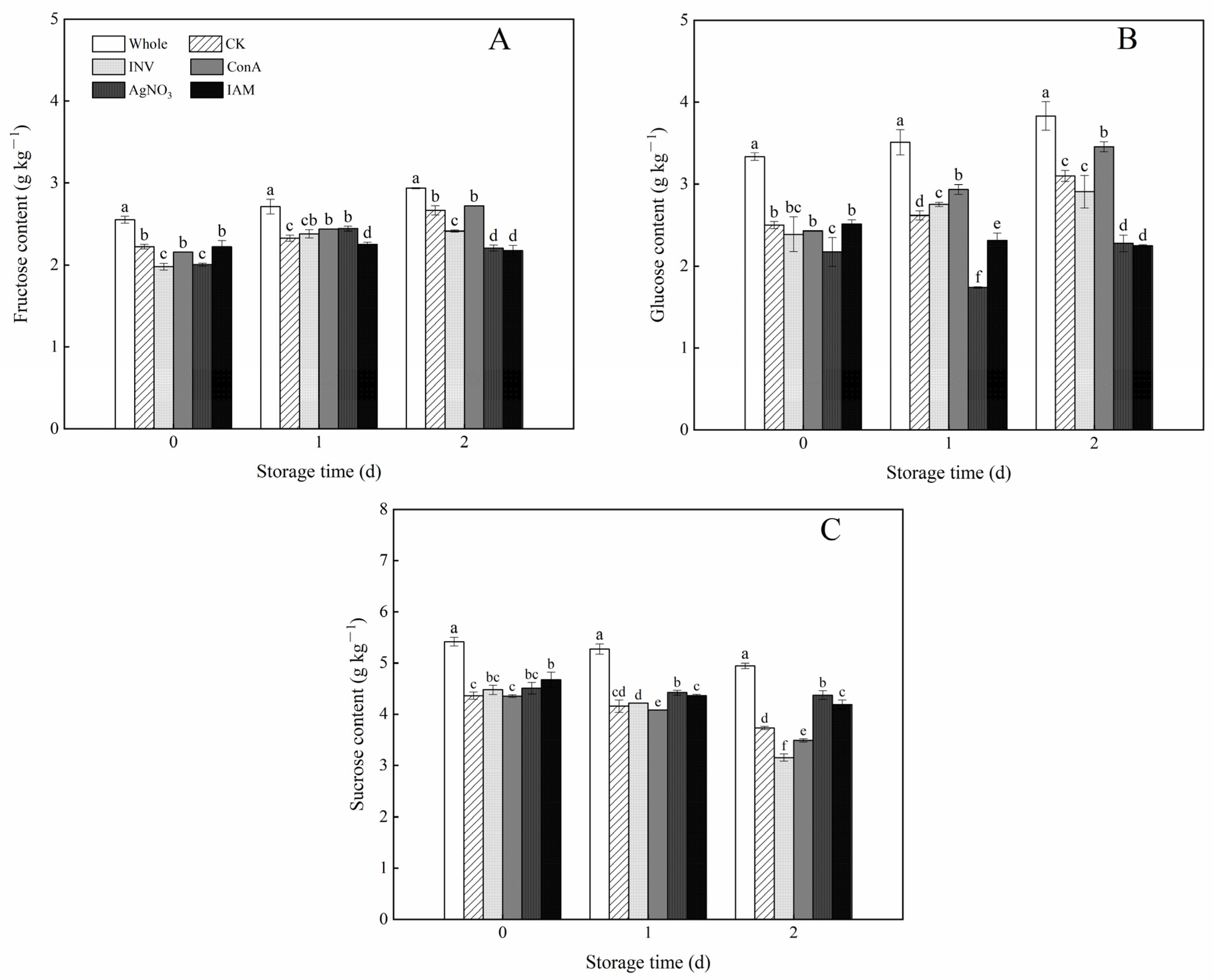
3.7. Effects of INV Activators and Inhibitors on Soluble Sugar Contents of Cubed Carrots
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lahlou, A.; Chileh-Chelh, T.; Lyashenko, S.; Rincón-Cervera, M.Á.; Rodríguez-García, I.; López-Ruiz, R.; Urrestarazu, M.; Guil-Guerrero, J.L. Arecaceae fruits: Fatty acids, phenolic compounds and in vitro antitumor activity. Food Biosci. 2022, 50, 102181. [Google Scholar] [CrossRef]
- Mizgier, P.; Kucharska, A.Z.; Sokół-Łętowska, A.; Kolniak-Ostek, J.; Kidoń, M.; Fecka, I. Characterization of phenolic compounds and antioxidant and anti-inflammatory properties of red cabbage and purple carrot extracts. J. Funct. Foods 2016, 21, 133–146. [Google Scholar] [CrossRef]
- Sen, S.; Kalita, P.; Chakraborty, R. Evaluation of hypolipidemic, antioxidant, atherogenic index and cardiac risk suppressing effects of unpolished maniki madhuri rice extract and HPLC analysis of phenolics compounds. J. Cereal Sci 2022, 108, 103581. [Google Scholar] [CrossRef]
- Viacava, F.; Santana-Gálvez, J.; Heredia-Olea, E.; Pérez-Carrillo, E.; Nair, V.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Sequential application of postharvest wounding stress and extrusion as an innovative tool to increase the concentration of free and bound phenolics in carrots. Food Chem. 2020, 307, 125551. [Google Scholar] [CrossRef]
- Li, X.A.; Li, M.L.; Ji, N.N.; Jin, P.; Zhang, J.H.; Zheng, Y.H.; Zhang, X.H.; Li, F.J. Cold plasma treatment induces phenolic accumulation and enhances antioxidant activity in fresh-cut pitaya (Hylocereus undatus) fruit. LWT-Food Sci. Technol. 2019, 115, 108447. [Google Scholar] [CrossRef]
- Guan, Y.G.; Hu, W.Z.; Jiang, A.L.; Xu, Y.P.; Zhao, M.R.; Yu, J.X.; Ji, Y.R.; Sarengaowa; Yang, X.Z.; Feng, K. The effect of cutting style on the biosynthesis of phenolics and cellular antioxidant capacity in wounded broccoli. Food Res. Int. 2020, 137, 109565. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.Z.; Guan, Y.G.; Ji, Y.R.; Yang, X.Z. Effect of cutting styles on quality, antioxidant activity, membrane lipid peroxidation, and browning in fresh-cut potatoes. Food Biosci. 2021, 44, 101435. [Google Scholar] [CrossRef]
- Surjadinata, B.B.; Cisneros-Zevallos, L. Biosynthesis of phenolic antioxidants in carrot tissue increases with wounding intensity. Food Chem. 2012, 134, 615–624. [Google Scholar] [CrossRef]
- Formica-Oliveira, A.C.; Martínez-Hernández, G.B.; Díaz-López, V.; Artés, F.; Artés-Hernández, F. Effects of UV-B and UV-C combination on phenolic compounds biosynthesis in fresh-cut carrots. Postharvest Biol. Technol. 2017, 127, 99–104. [Google Scholar] [CrossRef]
- Heredia, J.B.; Cisneros-Zevallos, L. The effect of exogenous ethylene and methyl jasmonate on pal activity, phenolic profiles and antioxidant capacity of carrots (Daucus carota) under different wounding intensities. Postharvest Biol. Technol. 2019, 51, 242–249. [Google Scholar] [CrossRef]
- Jacobo-Velázquez, D.A.; Martinez-Hernandez, G.B.; Rodriguez, S.; Cao, C.M.; Cisneros-Zevallos, L. Plants as biofactories: Physiological role of reactive oxygen species on the accumulation of phenolic antioxidants in carrot tissue under wounding and hyperoxia stress. J. Agric. Food Chem. 2011, 59, 6583–6593. [Google Scholar] [CrossRef] [PubMed]
- Becerra-Moreno, A.; Redondo-Gil, M.; Benavides, J.; Nair, V.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Combined effect of water loss and wounding stress on gene activation of metabolic pathways associated with phenolic biosynthesis in carrot. Front Plant Sci. 2015, 6, 837. [Google Scholar] [CrossRef] [PubMed]
- Becerra-Moreno, A.; Benavides, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Plants as biofactories: Glyphosate-induced production of shikimic acid and phenolic antioxidants in wounded carrot tissue. J. Agric. Food Chem. 2012, 60, 11378–11386. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Li, J.; Jin, P.; Li, X.A.; Wang, L.; Zheng, Y.H. The effect of temperature on phenolic content in wounded carrots. Food Chem. 2017, 215, 116–123. [Google Scholar] [CrossRef]
- Jacobo-Velázquez, D.A.; Cisneros-Zevallos, L. An alternative use of horticultural crops: Stressed plants as biofactories of bioactive phenolic compounds. Agriculture 2012, 2, 259–271. [Google Scholar] [CrossRef]
- Jacobo-Velázquez, D.A.; González-Agüero, M.; Cisneros-Zevallos, L. Cross-talk between signaling pathways: The link between plant secondary metabolite production and wounding stress response. Sci. Rep. 2015, 5, 8608. [Google Scholar] [CrossRef] [PubMed]
- Surjadinata, B.B.; Jacobo-Velázquez, D.A.; Cisneros-Zevallos, L. Physiological role of reactive oxygen species, ethylene, and jasmonic acid on UV light induced phenolic biosynthesis in wounded carrot tissue. Postharvest Biol. Technol. 2021, 172, 111388. [Google Scholar] [CrossRef]
- Hahlbrock, K.; Scheel, D. Physiology and molecular biology of phenylpropanoid metabolism. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 347–369. [Google Scholar] [CrossRef]
- Umbarger, H.E. Amino acid biosynthesis and its regulation. Annu. Rev. Biochem. 1978, 47, 533–606. [Google Scholar] [CrossRef] [PubMed]
- Torres-Contreras, A.M.; Nair, V.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Effect of exogenous amylolytic enzymes on the accumulation of chlorogenic acid isomers in wounded potato tubers. J. Agric. Food Chem. 2014, 62, 7671–7675. [Google Scholar] [CrossRef]
- Li, X.A.; Li, M.L.; Wang, J.; Wang, L.; Han, C.; Jin, P.; Zheng, Y.H. Methyl jasmonate enhances wound-induced phenolic accumulation in pitaya fruit by regulating sugar content and energy status. Postharvest Biol. Technol. 2018, 137, 106–112. [Google Scholar] [CrossRef]
- Han, C.; Jin, P.; Li, M.L.; Wang, L.; Zheng, Y.H. Physiological and transcriptomic analysis validates previous findings of changes in primary metabolism for the production of phenolic antioxidants in wounded carrots. J. Agric. Food Chem. 2017, 65, 7159–7167. [Google Scholar] [CrossRef] [PubMed]
- Qin, G.Z.; Zhu, Z.; Wang, W.H.; Cai, J.H.; Chen, Y.; Li, L.; Tian, S.P. A tomato vacuolar invertase inhibitor mediates sucrose metabolism and Influences fruit ripening. Plant Physiol. 2016, 172, 1596–1611. [Google Scholar] [CrossRef]
- Liu, C.C.; Huang, L.C.; Chang, C.T.; Sung, H.Y. Purification and characterization of soluble invertases from suspension-cultured bamboo (Bambusa edulis) cells. Food Chem. 2006, 96, 621–631. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Byrne, D.H. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Wang, K.; Shao, X.F.; Gong, Y.F.; Zhu, Y.; Wang, H.F.; Zhang, X.L.; Yu, D.D.; Yu, F.; Qiu, Z.Y.; Lu, H. The metabolism of soluble carbohydrates related to chilling injury in peach fruit exposed to cold stress. Postharvest Biol. Technol. 2013, 86, 53–61. [Google Scholar] [CrossRef]
- Zhen, W.N.; Tu, Y.; Lin, Z.H.; Xu, X.X.; Fu, M.R.; Han, C. Comparative transcriptome analysis reveals the molecular mechanism of UV-B irradiation in promoting the accumulation of phenolic compounds in wounded carrot. Horticulturae 2022, 8, 896. [Google Scholar] [CrossRef]
- Li, M.L.; Li, X.A.; Han, C.; Ji, N.N.; Jin, P.; Zheng, Y.H. Physiological and metabolomic analysis of cold plasma treated fresh-cut strawberries. J. Agric. Food Chem. 2019, 67, 4043–4053. [Google Scholar] [CrossRef]
- Jacobo-Velázquez, D.A. Transformation of carrots into novel food ingredients and innovative healthy foods. Appl. Food Res. 2023, 3, 100303. [Google Scholar] [CrossRef]
- Yoshida, S. Biosynthesis and conversion of aromatic amino acids in plants. Annu. Rev. Plant Physiol. 1969, 20, 41–62. [Google Scholar] [CrossRef]
- Scalbert, A.; Johnson, I.T.; Saltmarsh, M. Polyphenols: Antioxidants and beyond. Am. J. Clin. Nutr. 2005, 81, 215S–217S. [Google Scholar] [CrossRef]
- Linask, J.; Laties, G.G. Multiphasic absorption of glucose and 3-O-methyl glucose by aged potato slices. Plant Physiol. 1973, 51, 289–294. [Google Scholar] [CrossRef]
- Dai, Z.W.; Léon, C.; Feil, R.; Lunn, J.E.; Delrot, S.; Gomeès, E. Metabolic profiling reveals coordinated switches in primary carbohydrate metabolism in grape berry (Vitis vinifera L.), a non-climacteric fleshy fruit. J. Exp. Bot. 2013, 64, 1345–1355. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, P.B.; Wu, L.; Busse, J.S.; Whitty, B.R.; Hamernik, A.J.; Jansky, S.H.; Buell, C.R.; Bethke, P.C.; Jiang, J.M. Suppression of the vacuolar invertase gene prevents cold-induced sweetening in potato. Plant Physiol. 2010, 154, 939–948. [Google Scholar] [CrossRef] [PubMed]
- Sturm, A.; Tang, G. The sucrose-cleaving enzymes of plants are crucial for development, growth and carbon partitioning. Trends Plant Sci. 1999, 4, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Roitsch, T.; González, M.C. Function and regulation of plant invertases: Sweet sensations. Trends Plant Sci. 2004, 9, 606–613. [Google Scholar] [CrossRef]
- Vargas, W.; Pontis, H.; Salerno, G. Differential expression of alkaline and neutral invertases in response to environmental stresses: Characterization of an alkaline isoform as a stress response enzyme in wheat leaves. Planta 2007, 226, 1535–1545. [Google Scholar] [CrossRef]
- Yu, L.N.; Liu, H.X.; Shao, X.F.; Yu, F.; Wei, Y.Z.; Ni, Z.M.; Xu, F.; Wang, H.F. Effects of hot air and methyl jasmonate treatment on the metabolism of soluble sugars in peach fruit during cold storage. Postharvest Biol. Technol. 2016, 113, 8–16. [Google Scholar] [CrossRef]
- Qian, W.J.; Yue, C.; Wang, Y.C.; Cao, H.L.; Li, N.N.; Wang, L.; Hao, X.Y.; Wang, X.C.; Xiao, B.; Yang, Y.J. Identification of the invertase gene family (INVs) in tea plant and their expression analysis under abiotic stress. Plant Cell Rep. 2016, 35, 2269–2283. [Google Scholar] [CrossRef] [PubMed]
- Sturm, A.; Chrispeels, M.J. cDNA cloning of carrot extracellular beta-fructosidase and its expression in response to wounding and bacterial infection. Plant Cell 1990, 2, 1107–1119. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, X.; Li, M.; Zhang, M.; Zhao, R.; Han, C. Invertase Plays a Vital Role in Regulating Wound-Induced Accumulation of Phenolic Compounds in Carrot Tissue. Horticulturae 2024, 10, 72. https://doi.org/10.3390/horticulturae10010072
Ren X, Li M, Zhang M, Zhao R, Han C. Invertase Plays a Vital Role in Regulating Wound-Induced Accumulation of Phenolic Compounds in Carrot Tissue. Horticulturae. 2024; 10(1):72. https://doi.org/10.3390/horticulturae10010072
Chicago/Turabian StyleRen, Xinle, Meilin Li, Meixue Zhang, Ruping Zhao, and Cong Han. 2024. "Invertase Plays a Vital Role in Regulating Wound-Induced Accumulation of Phenolic Compounds in Carrot Tissue" Horticulturae 10, no. 1: 72. https://doi.org/10.3390/horticulturae10010072
APA StyleRen, X., Li, M., Zhang, M., Zhao, R., & Han, C. (2024). Invertase Plays a Vital Role in Regulating Wound-Induced Accumulation of Phenolic Compounds in Carrot Tissue. Horticulturae, 10(1), 72. https://doi.org/10.3390/horticulturae10010072