


Microbial Consortia Versus Single-Strain Inoculants as Drought Stress Protectants in Potato Affected by the Form of N Supply
 , and
, and
Abstract
1. Introduction
- (i)
- to characterize the drought-protective potential of microbial strains and strain combinations with PGPM potential under controlled greenhouse conditions.
- (ii)
- to characterize under field conditions the drought-protective potential of the best- performing microbial strains and strain combinations pre-selected in the controlled experiments.
- (iii)
- to characterize the response of plants treated with drought-protective single-strain inoculants and microbial consortia to NH4+ supply, using NO3- fertilization as a control.
- (iv)
- to identify underlying modes of action.
2. Materials and Methods
2.1. Greenhouse Experiments
2.1.1. Potato Culture
2.1.2. Maintenance Culture of Inoculants
2.1.3. Preparation and Application of Inoculants
2.1.4. Setup of Pot Experiment I
2.1.5. Setup of Pot Experiment II
2.1.6. Visual Rating of Plant Performance
2.1.7. Harvest of Plant Material
2.1.8. Root Length Measurement
2.1.9. Analysis of Mineral Nutrients
2.1.10. Determination of Stress Metabolites and Hormonal Profiling
2.2. Field Trial
2.2.1. Fertilization and Irrigation
2.2.2. Microbial Inoculants, Formulation, Dosage, and Application
2.3. Experimental Design and Statistical Analysis of Greenhouse Experiments
2.4. Statistical Analysis of the Field Trial
3. Results
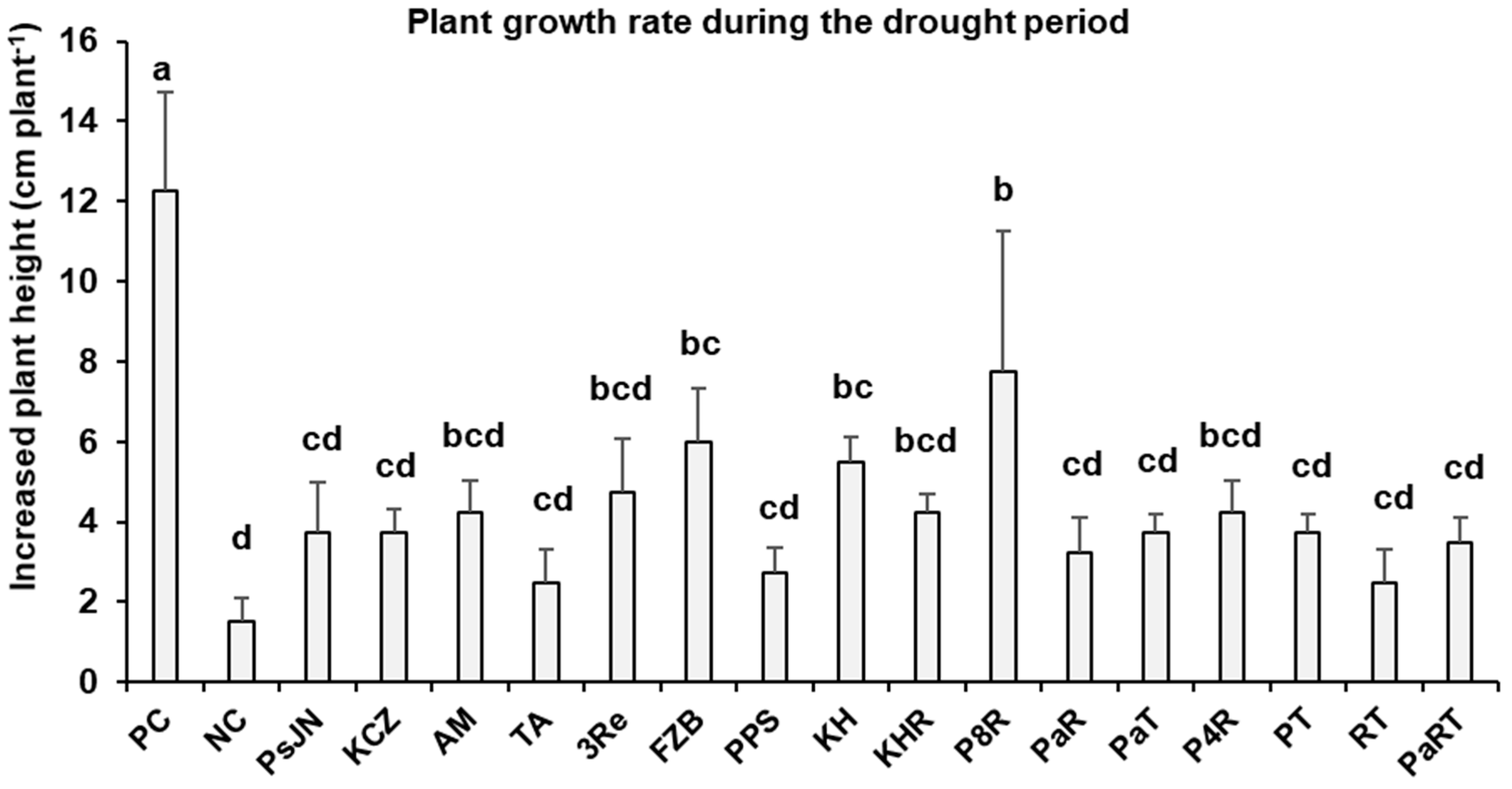
3.1. Pot Experiment I: Plant Growth Parameters
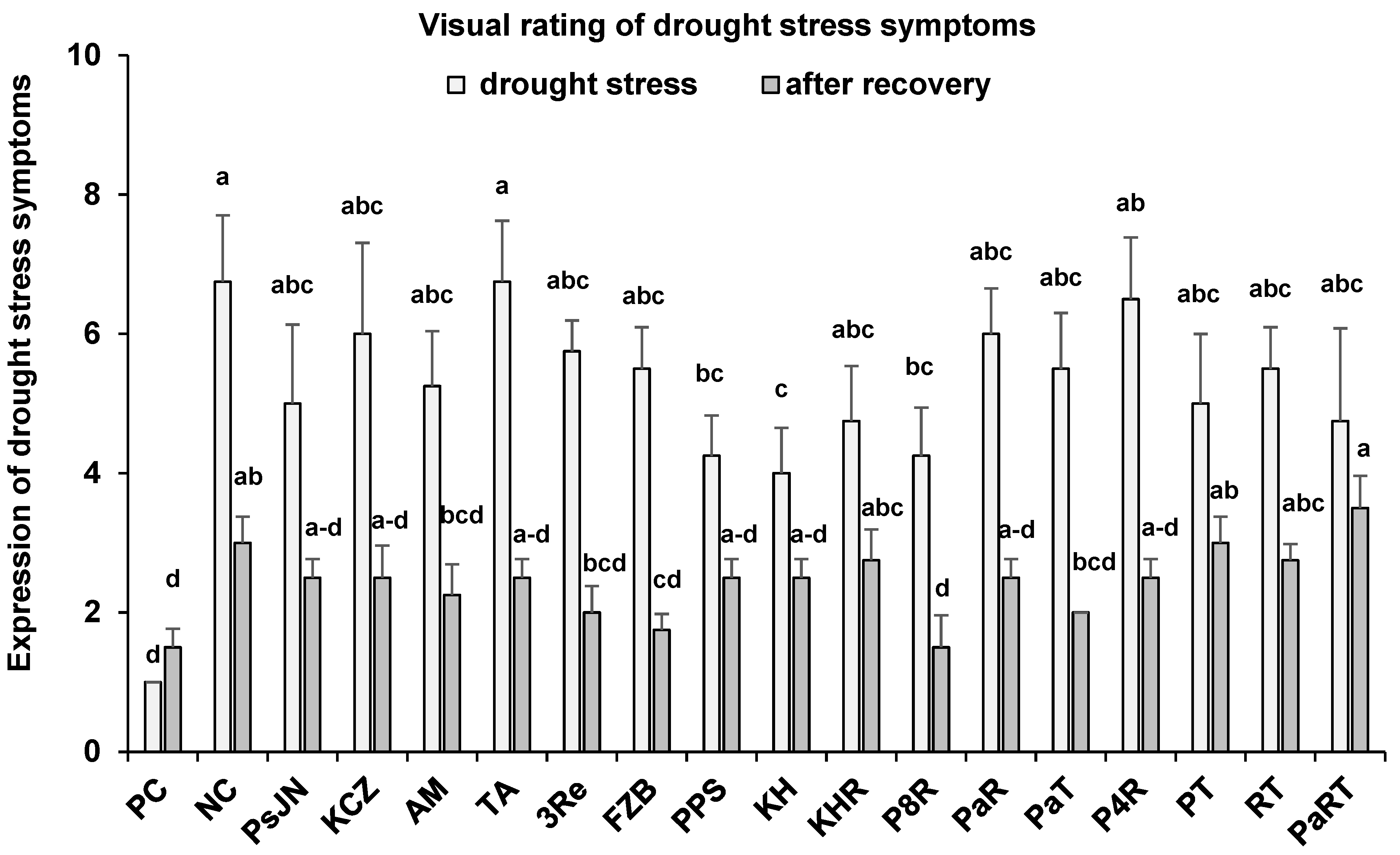
3.1.1. Visual Evaluation of Drought Stress Symptoms
3.1.2. Mineral Nutritional Status
3.2. Field Experiment
3.3. Pot Experiment II
3.3.1. Plant Growth Parameters
3.3.2. Rating of Drought Stress Symptoms
3.3.3. Plant Nutritional Status
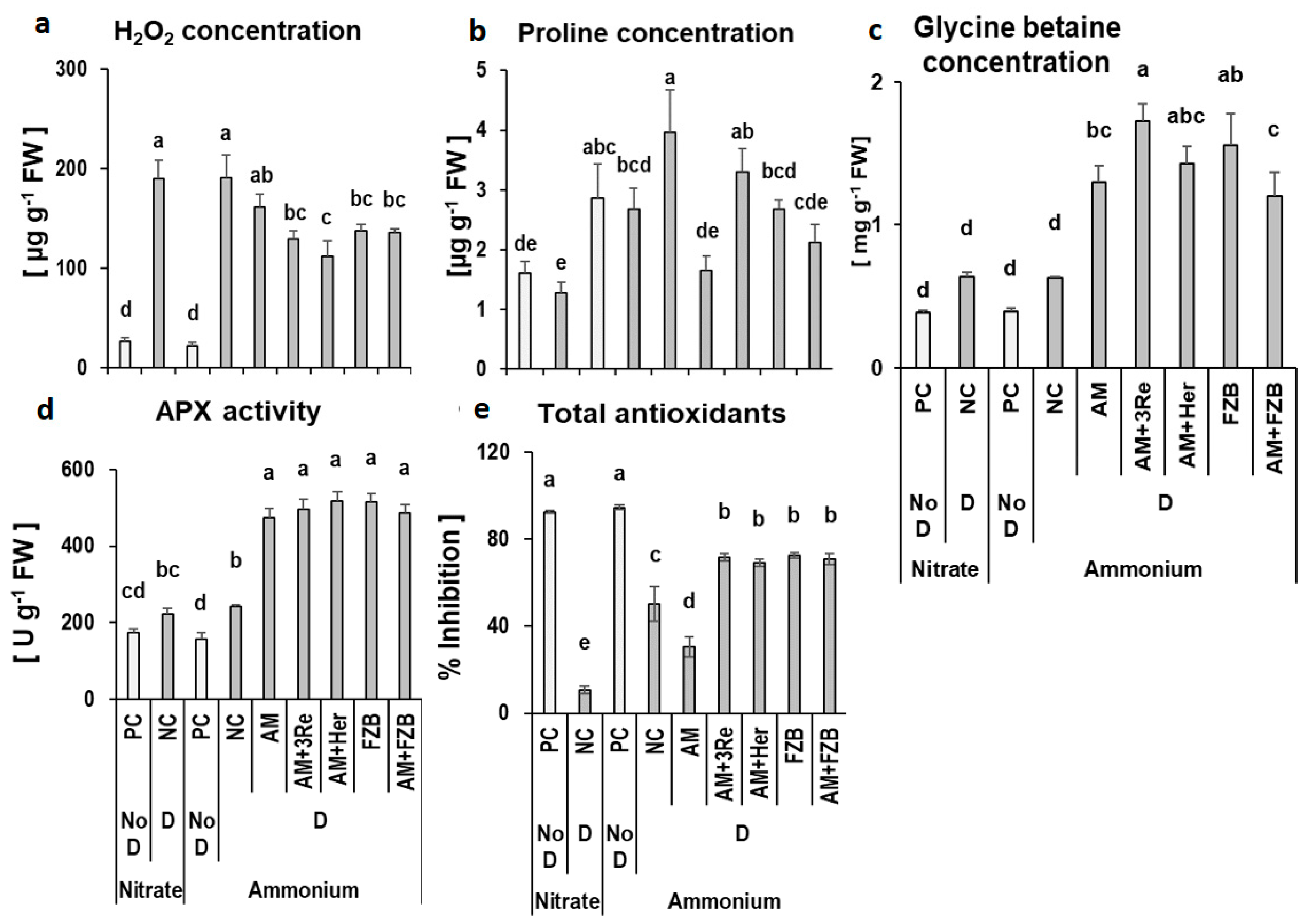
3.3.4. Stress Metabolites in Leaves
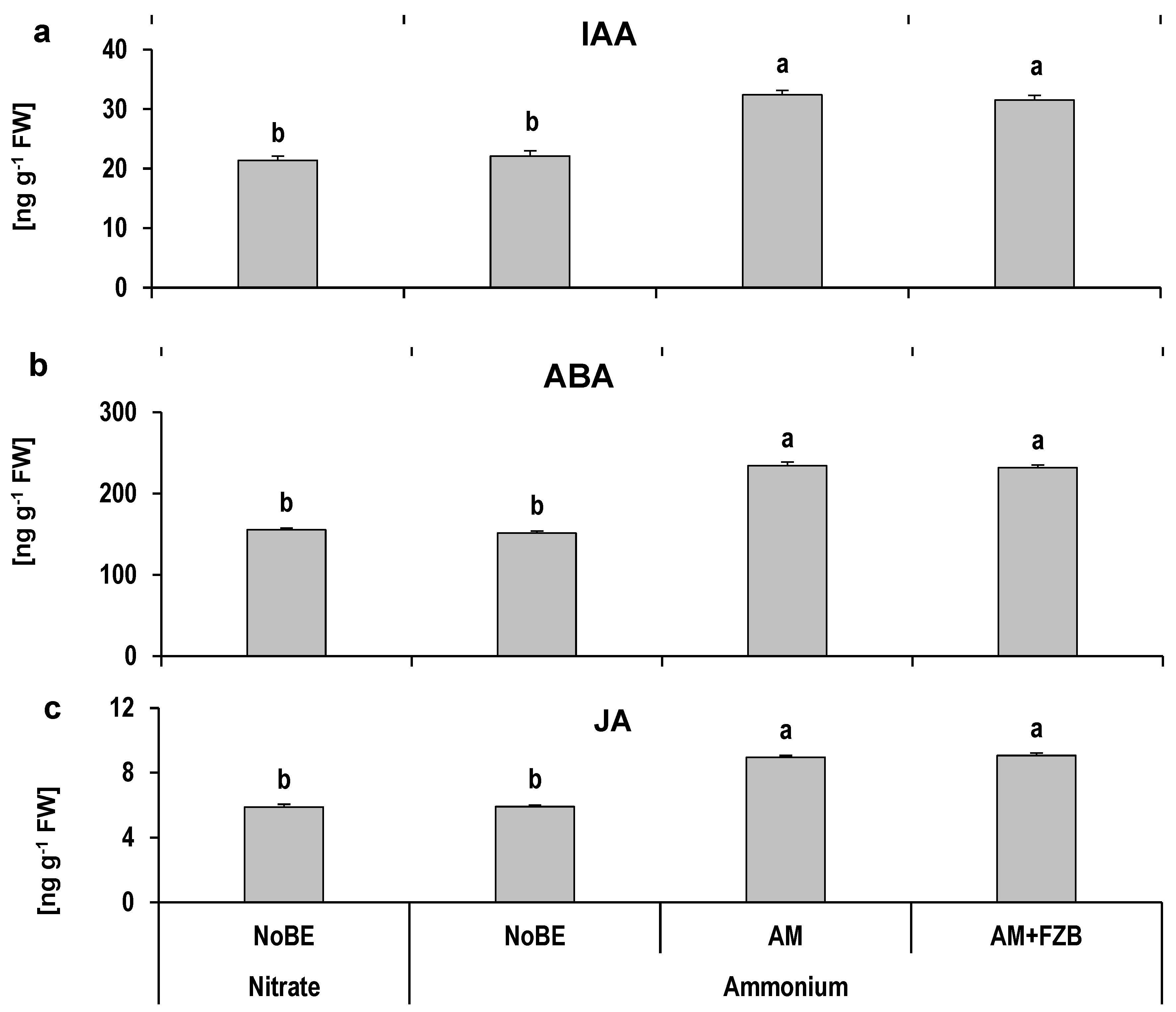
3.3.5. Hormonal Status of Drought-Affected Plants
4. Discussion
4.1. Screening for the Effects of Drought Protection Inoculants under NO3− Fertilization
4.2. Effects of Drought Protection Inoculants under NH4+ Fertilization
4.3. Effects on Plant Growth and Nutritional Status
4.4. Effects of AMF on Oxidative Stress Protection and Osmotic Adjustment
4.5. Effects of AMF on Tuberization
4.6. Effects of Bacterial Inoculants and Microbial Consortia
4.7. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAO. FAO Statistical Databases FAOSTAT. 2014. Available online: http://faostat3.fao.org/ (accessed on 20 October 2023).
- Devaux, A.; Kromann, P.; Ortiz, O. Potatoes for sustainable global food security. Potato Res. 2014, 57, 185–199. [Google Scholar] [CrossRef]
- Zarzyńska, K.; Boguszewska-Mańkowska, D.; Nosalewicz, A. Differences in size and architecture of the potato cultivars root system and their tolerance to drought stress. Plant Soil Environ. 2017, 63, 159–164. [Google Scholar] [CrossRef]
- Hopkins, B.G.; Horneck, D.A.; MacGuidwin, A.E. Improving phosphorus use efficiency through potato rhizosphere modification and extension. Am. J. Potato Res. 2014, 91, 161–174. [Google Scholar] [CrossRef]
- Koch, M.; Naumann, M.; Pawelzik, E.; Gransee, A.; Thiel, H. The importance of nutrient management for potato production Part I: Plant nutrition and yield. Potato Res. 2020, 63, 97–119. [Google Scholar] [CrossRef]
- van der Linden, C.G.; Anithakumari, A.M.; van Culemborg, M.; Visser, R.G.F. Dissecting the genetics of abiotic stress tolerance in potato. In Proceedings of the Plant & Animal Genomes XIXth Conference, San Diego, CA, USA, 15–19 January 2011. [Google Scholar]
- Neumann, G.; Römheld, V. Root-induced changes in the availability of nutrients in the rhizosphere. In Plant Roots: The Hidden Half; Marcel Dekker Inc.: New York, NY, USA, 2002; pp. 617–649. [Google Scholar]
- Aliche, E.B.; Theeuwen, T.P.; Oortwijn, M.; Visser, R.G.; van der Linden, C.G. Carbon partitioning mechanisms in potato under drought stress. Plant Physiol. Biochem. 2020, 146, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Hijmans, R.J. The effect of climate change on global potato production. Am. J. Potato Res. 2003, 80, 271–279. [Google Scholar] [CrossRef]
- Raymundo, R.; Asseng, S.; Robertson, R.; Petsakos, A.; Hoogenboom, G.; Quiroz, R.; Wolf, J. Climate change impact on global potato production. Eur. J. Agron. 2018, 100, 87–98. [Google Scholar] [CrossRef]
- Halpern, M.; Bar-Tal, A.; Ofek, M.; Minz, D.; Muller, T.; Yermiyahu, U. The use of biostimulants for enhancing nutrient uptake. Adv. Agron. 2015, 130, 141–174. [Google Scholar] [CrossRef]
- Meena, K.K.; Sorty, A.M.; Bitla, U.M.; Choudhary, K.; Gupta, P.; Pareek, A.; Singh, H.B. Abiotic stress responses and microbe-mediated mitigation in plants: The omics strategies. Front. Plant Sci. 2017, 8, 172. [Google Scholar] [CrossRef]
- Armada, E.; Portela, G.; Roldán, A.; Azcón, R. Combined use of beneficial soil microorganism and agrowaste residue to cope with plant water limitation under semiarid conditions. Geoderma 2014, 232, 640–648. [Google Scholar] [CrossRef]
- Bukovská, P.; Bonkowski, M.; Konvalinková, T.; Beskid, O.; Hujslová, M.; Püschel, D.; Jansa, J. Utilization of organic nitrogen by arbuscular mycorrhizal fungi is there a specific role for protists and ammonia oxidizers. Mycorrhiza 2018, 28, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Brookes, P.C.; Xu, J. Arbuscular mycorrhizal fungal hyphae alter soil bacterial community and enhance polychlorinated biphenyls dissipation. Front. Microbiol. 2016, 7, 939. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Fimognari, L.; de Almeida, J.; Jensen, C.N.G.; Compant, S.; Oliveira, T.; Liu, F. Effect of Bacillus paralicheniformis on soybean (Glycine max) roots colonization, nutrient uptake and water use efficiency under drought stress. J. Agron. Crop Sci. 2023, 209, 547–565. [Google Scholar] [CrossRef]
- Barea, J.M.; Toro, M.; Orozco, M.O.; Campos, E.; Azcón, R. The application of isotopic (32 P and 15 N) dilution techniques to evaluate the interactive effect of phosphate-solubilizing rhizobacteria, mycorrhizal fungi and Rhizobium to improve the agronomic efficiency of rock phosphate for legume crops. Nutr. Cycl. Agroecosyst. 2002, 63, 35–42. [Google Scholar] [CrossRef]
- Arora, N.K.; Mishra, J. Prospecting the roles of metabolites and additives in future bioformulations for sustainable agriculture. Appl. Soil Ecol. 2016, 107, 405–407. [Google Scholar] [CrossRef]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef]
- Glick, B.R.; Cheng, Z.; Czarny, J.; Duan, J. Promotion of plant growth by ACC deaminase-producing soil bacteria. In New Perspectives and Approaches in Plant Growth-Promoting Rhizobacteria Research; Springer: Berlin/Heidelberg, Germany, 2007; pp. 329–339. [Google Scholar] [CrossRef]
- Schütz, L.; Gattinger, A.; Meier, M.; Müller, A.; Boller, T.; Mäder, P.; Mathimaran, N. Improving crop yield and nutrient use efficiency via biofertilization—A global meta-analysis. Front. Plant Sci. 2018, 8, 2204. [Google Scholar] [CrossRef]
- Weinmann, M.; Neumann, G. Bio-effectors to optimize the mineral nutrition of crop plants. In Achieving Sustainable Crop Nutrion; Rengel, Z., Ed.; Burleigh Dodds Science Publishing Limited: Camebridge, UK, 2020; pp. 589–690. [Google Scholar] [CrossRef]
- Woo, S.L.; Pepe, O. Microbial consortia: Promising probiotics as plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1801. [Google Scholar] [CrossRef]
- Aguilar-Paredes, A.; Valdés, G.; Nuti, M. Ecosystem functions of microbial consortia in sustainable agriculture. Agronomy 2020, 10, 1902. [Google Scholar] [CrossRef]
- Bradáčová, K.; Florea, A.S.; Bar-Tal, A.; Minz, D.; Yermiyahu, U.; Shawahna, R.; Weinmann, M. Microbial consortia versus single-strain inoculants: An advantage in PGPM-assisted tomato production. Agronomy 2019, 9, 105. [Google Scholar] [CrossRef]
- Nkebiwe, P.M.; Weinmann, M.; Müller, T. Improving fertilizer-depot exploitation and maize growth by inoculation with plant growth-promoting bacteria: From lab to field. Chem. Biol. Technol. Agric. 2016, 3, 1–16. [Google Scholar] [CrossRef]
- Mpanga, I.K.; Gomez-Genao, N.; Moradtalab, N.; Wanke, D.; Chrobaczek, V.; Ahmed, A.; Ludewig, U. The role of N form supply for PGPM-host plant interactions in maize. J. Plant Nutr. Soil Sci. 2019, 182, 908–920. [Google Scholar] [CrossRef]
- Mpanga, I.K.; Nkebiwe, P.M.; Kuhlmann, M.; Cozzolino, V.; Piccolo, A.; Geistlinger, J.; Neumann, G. The form of N supply determines plant growth promotion by P-solubilizing microorganisms in maize. Microorganisms 2019, 7, 38. [Google Scholar] [CrossRef] [PubMed]
- Moradtalab, N.; Ahmed, A.; Geistlinger, J.; Walker, F.; Höglinger, B.; Ludewig, U.; Neumann, G. Synergisms of microbial consortia, N forms, and micronutrients alleviate oxidative damage and stimulate hormonal cold stress adaptations in maize. Front. Plant Sci. 2020, 11, 396. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.; Baumgartner, N. Unterfussdüngung bei Kartoffeln. BLW 2017, 12, 52–53. [Google Scholar]
- Öhlinger, R. Maximum Water-Holding Capacity. In Methods in Soil Biology; Schinner, F., Öhlinger, R., Kandeler, E., Margesin, R., Eds.; Springer: Heidelberg, Germany, 1996; pp. 385–386. [Google Scholar]
- De Jaeger, N.; De la Providencia, I.E.; Rouhier, H.; Declerck, S. Co-entrapment of Trichoderma harzianum and Glomus sp. within alginate beads: Impact on the arbuscular mycorrhizal fungi life cycle. J. Appl. Microbiol. 2011, 111, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Microbial Inoculants Provided by Solace Project Partners. Available online: https://www.solace-eu.net (accessed on 17 October 2018).
- Gericke, S.; Kurmies, B. Die kolorimetrische phosphorsäurebestimmung mit ammonium-vanadat-molybdat und ihre Anwendung in der Pflanzenanalyse. Z. Pflanzenernährung Düngung Bodenkd. 1952, 59, 235–247. [Google Scholar]
- Moradtalab, N.; Weinmann, M.; Walker, F.; Höglinger, B.; Ludewig, U.; Neumann, G. Silicon improves chilling tolerance during early growth of maize by effects on micronutrient homeostasis and hormonal balances. Front. Plant Sci. 2018, 9, 420. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R. Rapid assay for determination of water soluble quaternary ammonium compounds. Plant Soil 1983, 70, 303–307. [Google Scholar] [CrossRef]
- Valadez-Bustos, M.G.; Aguado-Santacruz, G.A.; Tiessen-Favier, A.; Robledo-Paz, A.; Muñoz-Orozco, A.; Rascón-Cruz, Q.; Santacruz-Varela, A. A reliable method for spectrophotometric determination of glycine betaine in cell suspension and other systems. Anal. Biochem. 2016, 498, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Boominathan, R.; Doran, P.M. Ni-induced oxidative stress in roots of the Ni hyperaccumulator, Alyssum bertolonii. New Phytol. 2002, 156, 205–215. [Google Scholar] [CrossRef] [PubMed]
- CR Team. Team RDC. R: A Language and Environment for Statistical Computing; R foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Allaire, J. RStudio: Integrated development environment for R. Boston MA 2012, 770, 165–171. [Google Scholar]
- de Mendiburu, F. Package ‘agricolae’. In R Package, Version; R foundation for Statistical Computing: Vienna, Austria, 2019; Volume 1. [Google Scholar]
- Kumar, A.; Verma, J.P. Does plant—Microbe interaction confer stress tolerance in plants: A review? Microbiol. Res. 2018, 207, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Velivelli, S.L.; Sessitsch, A.; Prestwich, B.D. The role of microbial inoculants in integrated crop management systems. Potato Res. 2014, 57, 291–309. [Google Scholar] [CrossRef]
- Overbeek, W.; Jeanne, T.; Hogue, R.; Smith, D.L. Effects of Microbial Consortia, Applied as Fertilizer Coating, on Soil and Rhizosphere Microbial Communities and Potato Yield. Front. Agron. 2021, 3, 714700. [Google Scholar] [CrossRef]
- Verbruggen, E.; van der Heijden, M.G.; Rillig, M.C.; Kiers, E.T. Mycorrhizal fungal establishment in agricultural soils: Factors determining inoculation success. New Phytol. 2013, 197, 1104–1109. [Google Scholar] [CrossRef]
- Bradáčová, K.; Kandeler, E.; Berger, N.; Ludewig, U.; Neumann, G. Microbial consortia inoculants stimulate early growth of maize depending on nitrogen and phosphorus supply. Plant Soil Environ. 2020, 66, 105–112. [Google Scholar] [CrossRef]
- Sharp, R.E.; LeNoble, M.E. ABA, ethylene and the control of shoot and root growth under water stress. J. Exp. Bot. 2002, 53, 33–37. [Google Scholar] [CrossRef]
- Mackay, A.D.; Barber, S.A. Soil moisture effects on root growth and phosphorus uptake by com. Agron. J. 1985, 77, 519–523. [Google Scholar] [CrossRef]
- Sharma, S.K.; Ramesh, A.; Johri, B.N. Isolation and characterization of plant growth-promoting Bacillus amyloliquefaciens strain sks_bnj_1 and its influence on rhizosphere soil properties and nutrition of soybean (Glycine max L. Merrill). J. Virol. Microbiol. 2013, 2013, 446006. [Google Scholar] [CrossRef]
- Thonar, C.; Lekfeldt, J.D.S.; Cozzolino, V.; Kundel, D.; Kulhánek, M.; Mosimann, C.; Mäder, P. Potential of three microbial bio-effectors to promote maize growth and nutrient acquisition from alternative phosphorous fertilizers in contrasting soils. Chem. Biol. Technol. Agric. 2017, 4, 1–16. [Google Scholar] [CrossRef]
- Yusran, Y.; Roemheld, V.; Mueller, T. Effects of Pseudomonas sp.” Proradix” and Bacillus Amyloliquefaciens FZB42 on the Establishment of AMF Infection, Nutrient Acquisition and Growth of Tomato Affected by Fusarium Oxysporum Schlecht f. sp. Radicis-lycopersici Jarvis and Shoemaker. 2009. Available online: https://escholarship.org/uc/item/8g70p0zt (accessed on 20 September 2020).
- VDLUFA (Verband Deutscher Landwirtschaftlicher Untersuchungs-und Forschungsanstalten e.V. Speyer, Germany). Standpunkt. Phosphordüngung nach Bodenuntersuchung und Pflanzenbedarf; VDLUFA Verlag: Darmsatdt, Germany, 2018. [Google Scholar]
- Gao, Y.; Jia, L.; Hu, B.; Alva, A.; Fan, M. Potato stolon and tuber growth influenced by nitrogen form. Plant Prod. Sci. 2014, 17, 138–143. [Google Scholar] [CrossRef]
- Krauss, A. Interaction of nitrogen nutrition, phytohormones, and tuberization. Potato Physiol. 1985, S, 209–230. [Google Scholar]
- Wolf, B. Diagnostic Techniques for Improving Crop Production, 1st ed.; CRC Press: Boca Raton, FL, USA, 1996. [Google Scholar]
- Jing, J.; Rui, Y.; Zhang, F.; Rengel, Z.; Shen, J. Localized application of phosphorus and ammonium improves growth of maize seedlings by stimulating root proliferation and rhizosphere acidification. Field Crops Res. 2010, 119, 355–364. [Google Scholar] [CrossRef]
- Fernández-Lizarazo, J.C.; Moreno-Fonseca, L.P. Mechanisms for tolerance to water-deficit stress in plants inoculated with arbuscular mycorrhizal fungi. A review. Agron. Colomb. 2016, 34, 179–189. [Google Scholar] [CrossRef]
- Bahadur, A.; Batool, A.; Nasir, F.; Jiang, S.; Mingsen, Q.; Zhang, Q.; Feng, H. Mechanistic insights into arbuscular mycorrhizal fungi-mediated drought stress tolerance in plants. Int. J. Mol. Sci. 2019, 20, 4199. [Google Scholar] [CrossRef] [PubMed]
- Caverzan, A.; Passaia, G.; Rosa, S.B.; Ribeiro, C.W.; Lazzarotto, F.; Margis-Pinheiro, M. Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection. Genet. Mol. Biol. 2012, 35, 1011–1019. [Google Scholar] [CrossRef]
- Rajasheker, G.; Jawahar, G.; Jalaja, N.; Kumar, S.A.; Kumari, P.H.; Punita, D.L.; Kishor, P.B.K. Role and regulation of osmolytes and ABA interaction in salt and drought stress tolerance. In Plant Signaling Molecules; Woodhead Publishing: Cambridge, UK, 2019; pp. 417–436. [Google Scholar] [CrossRef]
- Wang, Y.; Hou, Y.; Qiu, J.; Wang, H.; Wang, S.; Tang, L.; Zhang, J. Abscisic acid promotes jasmonic acid biosynthesis via a ‘SAPK10-bZIP72-AOC’pathway to synergistically inhibit seed germination in rice (Oryza sativa). New Phytol. 2020, 228, 1336–1353. [Google Scholar] [CrossRef]
- Wang, X.; Li, Q.; Xie, J.; Huang, M.; Cai, J.; Zhou, Q.; Jiang, D. Abscisic acid and jasmonic acid are involved in drought priming-induced tolerance to drought in wheat. Crop J. 2021, 9, 120–132. [Google Scholar] [CrossRef]
- Cameron, D.D.; Neal, A.L.; van Wees, S.C.; Ton, J. Mycorrhiza-induced resistance: More than the sum of its parts. Trends Plant Sci. 2013, 18, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Méndez, S.J.; Urbano-Gálvez, A.; Mahouachi, J. Mitigation of salt stress damages in Carica papaya L. seedlings through exogenous pretreatments of gibberellic acid and proline. Chil. J. Agric. Res. 2022, 82, 167–176. [Google Scholar] [CrossRef]
- Jan, S.; Abbas, N.; Ashraf, M.; Ahmad, P. Roles of potential plant hormones and transcription factors in controlling leaf senescence and drought tolerance. Protoplasma 2019, 256, 313–329. [Google Scholar] [CrossRef] [PubMed]
- Dwelle, R.B. Photosynthesis and photoassimilate partitioning. In Potato Physiology; Academic Press Inc.: Palm Bay, FL, USA, 1985; pp. 35–58. [Google Scholar]
- Dutt, S.; Manjul, A.S.; Raigond, P.; Singh, B.; Siddappa, S.; Bhardwaj, V.; Kardile, H.B. Key players associated with tuberization in potato: Potential candidates for genetic engineering. Crit. Rev. Biotechnol. 2017, 37, 942–957. [Google Scholar] [CrossRef] [PubMed]
- Kolachevskaya, O.O.; Lomin, S.N.; Arkhipov, D.V.; Romanov, G.A. Auxins in potato: Molecular aspects and emerging roles in tuber formation and stress resistance. Plant Cell Rep. 2019, 38, 681–698. [Google Scholar] [CrossRef]
- Gururani, M.A.; Upadhyaya, C.P.; Baskar, V.; Venkatesh, J.; Nookaraju, A.; Park, S.W. Plant growth-promoting rhizobacteria enhance abiotic stress tolerance in Solanum tuberosum through inducing changes in the expression of ROS-scavenging enzymes and improved photosynthetic performance. J. Plant Growth Regul. 2013, 32, 245–258. [Google Scholar] [CrossRef]
- Batool, T.; Ali, S.; Seleiman, M.F.; Naveed, N.H.; Ali, A.; Ahmed, K.; Mubushar, M. Plant growth promoting rhizobacteria alleviates drought stress in potato in response to suppressive oxidative stress and antioxidant enzymes activities. Sci. Rep. 2020, 10, 1–19. [Google Scholar] [CrossRef]
- Santoyo, G.; Guzmán-Guzmán, P.; Parra-Cota, F.I.; Santos-Villalobos, S.D.L.; Orozco-Mosqueda, M.D.C.; Glick, B.R. Plant growth stimulation by microbial consortia. Agronomy 2021, 11, 219. [Google Scholar] [CrossRef]
- Miyawaki, K.; Matsumoto-Kitano, M.; Kakimoto, T. Expression of cytokinin biosynthetic isopentenyltransferase genes in Arabidopsis: Tissue specificity and regulation by auxin, cytokinin, and nitrate. Plant J. 2004, 37, 128–138. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variants | SDW (g Plant−1) | RDW (g Plant−1) | RL (cm Plant−1) | Root/Shoot Ratio | Tuber Weight (g Plant−1) | Tuber Number Plant−1 |
|---|---|---|---|---|---|---|
| PC | 7.53 a | 0.49 a | 6149 a | 0.096 bc | 71.86 a | 3.5 a |
| NC | 5.35 bc | 0.30 c | 3347 b | 0.094 bc | 35.88 bc | 2.8 a |
| PsJN | 5.26 bc | 0.32 bc | 3059 b | 0.092 bc | 29.72 bc | 2.3 a |
| KCZ | 5.48 bc | 0.32 bc | 3380 b | 0.081 bc | 35.64 bc | 3.0 a |
| AM | 5.18 bc | 0.30 c | 3299 b | 0.096 bc | 28.09 c | 2.8 a |
| TA | 5.22 bc | 0.35 bc | 3610 ab | 0.092 bc | 38.93 bc | 3.0 a |
| 3Re27 | 6.17 b | 0.43 ab | 4425 ab | 0.120 a | 29.36 bc | 3.0 a |
| FZB | 6.02 bc | 0.40 abc | 4123 ab | 0.102 ab | 31.86 bc | 2.5 a |
| PPS | 5.76 bc | 0.37 bc | 3637 ab | 0.096 bc | 31.72 bc | 3.0 a |
| KH | 5.21 bc | 0.30 c | 2927 b | 0.083 bc | 29.51 bc | 3.0 a |
| KHR | 5.53 bc | 0.31 c | 3019 b | 0.084 bc | 32.89 bc | 3.0 a |
| P8R | 6.00 bc | 0.40 abc | 3656 ab | 0.097 abc | 26.17 c | 3.0 a |
| PaR | 5.40 bc | 0.35 bc | 3465 b | 0.090 bc | 43.79 b | 3.3 a |
| PaT | 6.16 b | 0.39 abc | 3399 b | 0.098 ab | 26.30 c | 3.0 a |
| P4R | 5.71 bc | 0.39 abc | 3746 ab | 0.102 ab | 36.61 bc | 3.3 a |
| PT | 5.05 c | 0.29 c | 2737 b | 0.087 bc | 35.29 bc | 2.8 a |
| RT | 5.27 bc | 0.34 bc | 3371 b | 0.091 bc | 34.98 bc | 3.0 a |
| PaRT | 5.49 bc | 0.30 c | 2955 b | 0.073 c | 28.12 c | 3.5 a |
| (a) Concentrations | P | K | N | Mg | Mn | Zn |
|---|---|---|---|---|---|---|
| Deficiency threshold | 2.5 mg·g−1 | 60 mg·g−1 | 30 mg·g−1 | 7.0 mg·g−1 | 30 μg·g−1 | 30 μg·g−1 |
| PC | 2.12 c-e | 44.8 h | 33.1 e | 5.72 bc | 38.3 c | 104.2 a |
| NC | 1.87 e | 54.3 fg | 43.6 cd | 6.03 abc | 56.7 ab | 93.3 a |
| PsJN | 2.38 a–e | 61.0 abc | 49.3 ab | 6.42 ab | 59.2 ab | 100.8 a |
| KCZ | 2.26 a–e | 55.4 d–g | 45.6 bcd | 6.28 abc | 54.6 ab | 67.2 a |
| AM | 2.50 a–d | 60.8 abc | 46.0 bc | 6.18 abc | 58.9 ab | 73.5 a |
| TA | 2.22 b–e | 55.3 efg | 43.7 cd | 6.55 ab | 57.8 ab | 94.2 a |
| 3Re | 2.65 abc | 57.4 b–g | 45.9 bc | 6.51 ab | 53.3 b | 77.0 a |
| FZB | 2.49 a–d | 56.3 c–g | 45.3 bcd | 5.53 c | 54.4 ab | 82.9 a |
| PPS | 2.78 a | 61.4 ab | 48.7 ab | 6.35 abc | 53.8 ab | 82.6 a |
| KH | 2.70 ab | 62.9 a | 51.0 a | 6.57 ab | 56.2ab | 81.4 a |
| KHR | 2.45 a–d | 59.9 a–e | 49.2 ab | 6.35 abc | 56.0 ab | 86.1 a |
| P&R | 2.51 a–d | 59.4 a–e | 45.9 bc | 5.89 bc | 51.8 b | 73.8 a |
| PaR | 2.00 de | 57.7 b–g | 45.5 bcd | 6.87 a | 58.8 ab | 86.9 a |
| PaT | 2.78 a | 59.3 a–f | 47.0 abc | 6.20 abc | 56.9 ab | 91.9 a |
| P4R | 2.57 a–d | 54.1 g | 45.8 bc | 6.22 abc | 54.3 ab | 84.7 a |
| PT | 2.54 a–d | 60.4 a–d | 48.5 ab | 6.85 a | 59.5 ab | 68.4 a |
| RT | 2.68 abc | 63.7 a* | 41.3 d | 6.58 ab | 57.9 ab | 64.7 a |
| PaRT | 2.06 de | 56.7 b–g | 46.8 abc | 6.47 ab | 62.2 a | 84.2 a |
| (b) Contents | Macronutrients (mg·plant−1) | Micronutrients (μg·plant−1) | ||||
| P | K | N | Mg | Mn | Zn | |
| PC | 15.95 ab | 337.0 a–d | 249.2 a–f | 43.02 a | 288.3 a | 780.7 a |
| NC | 10.18 c | 291.4 cd | 232.6 def | 32.00 c | 300.9 a | 513.6 ab |
| PsJN | 12.67 abc | 321.1 a–d | 257.7 a–e | 33.61 bc | 313.1 a | 522.5 ab |
| KCZ | 12.31 abc | 303.1 bcd | 248.0 a–f | 34.43 bc | 298.9 a | 373.8 b |
| AM | 12.97 abc | 312.9 a–d | 235.8 c–f | 31.70 c | 301.4 a | 367.4 b |
| TA | 11.59 bc | 286.1 d | 225.3 ef | 34.11 bc | 302.3 a | 502.7 b |
| 3Re | 16.95 a* | 353.4 ab* | 281.0 ab | 39.09 ab | 323.6 a | 465.1 b |
| FZB | 15.11 abc | 338.8 a–d | 272.4 abc | 32.62 bc | 325.4 a | 503.7 b |
| PPS | 16.23 ab* | 351.5 abc | 278.9 ab | 36.59 abc | 308.6 a | 485.0 b |
| KH | 14.34 abc | 327.4 a–d | 264.8 a–e | 33.97 bc | 292.3 a | 430.0 b |
| KHR | 13.58 abc | 329.9 a–d | 270.5 a–d | 35.15 bc | 307.0 a | 482.6 b |
| P8R | 14.91 abc | 353.6 ab | 272.4 abc | 35.43 bc | 312.1 a | 453.8 ab |
| PaR | 10.73 c | 311.5 a–d | 245.2 b–f | 37.12 abc | 318.5 a | 459.9 b |
| PaT | 17.05 a | 363.8 a | 287.2 a | 38.06 abc | 350.8 a | 558.5 ab |
| P4R | 14.68 abc | 308.8 a–d | 261.4 a–e | 35.54 bc | 309.3 a | 479.6 b |
| PT | 12.66 abc | 302.7 bcd | 243.4 b–f | 34.79 bc | 302.3 a | 332.8 b |
| RT | 13.97 abc | 335.0 a–d | 216.0 f | 34.73 bc | 305.7 a | 342.1 b |
| PaRT | 11.33 bc | 312.1 a–d | 257.4 a–e | 35.63 bc | 340.7 a | 467.3 b |
| N Form | Treatments | SDW (g·Plant−1) | RDW (g·Plant−1) | RL (cm·Plant−1) | Tuber Weight (g·Plant−1) | Tuber Number Plant−1 | ITW (g) |
|---|---|---|---|---|---|---|---|
| NO3− | Well-watered | 13.92 ± 0.68 a | 0.64 ± 0.05 ab | 6086 ± 254 c | 141.19 ± 4.77 a | 3.0 ± 1.0 a | 68.0 ± 18.3 a |
| Drought stress | 11.74 ± 0.26 bc | 0.66 ± 0.04 a | 6543 ± 281 c | 3.23 ± 2.02 d | 1.8 ± 0.6 ab | 2.9 ± 2.1 b | |
| NH4+ | Well-watered | 13.69 ± 1.04 ab | 0.51 ± 0.05 b | 4721 ± 287 d | 135.61 ± 5.61 a | 1.8 ± 0.2 ab | 83.3 ± 17.9 a |
| Drought stress | 8.71 ± 0.66 f | 0.55 ± 0.06 ab | 6336 ± 349 c | 20.48 ± 5.53 bc | 1.2 ± 0.2 b | 19.4 ± 6.1 b | |
| AM | 9.37 ± 0.55 df | 0.53 ± 0.03 ab | 6931 ± 158 bc | 33.97 ± 1.38 b | 1.4 ± 0.2 b | 27.2 ± 4.4 b | |
| AM+3Re2-7 | 9.15 ± 0.81 ef | 0.55 ± 0.01 ab | 6708 ± 146 c | 17.13 ± 5.83 cd | 1.6 ± 0.4 ab | 12.8 ± 5.5 b | |
| AM+Her | 9.36 ± 0.99 def | 0.53 ± 0.07 ab | 6419 ± 424 c | 13,63 ± 5.22 cd | 1.2 ± 0.5 b | 13.1 ± 5.5 b | |
| FZB | 11.11 ± 0.93 cde | 0.57 ± 0.05 ab | 6296 ± 366 c | 13.52 ± 6.55 cd | 1.4 ± 0.5 b | 12.4 ± 6.9 b | |
| AM+FZB | 11.30 ± 0.38 cd | 0.65 ± 0.03 a | 7850 ± 351 a | 10.28 ± 5.22 cd | 0.8 ± 0.2 b | 10.3 ± 5.2 b |
| NO3WW | NO3D | NH4WW | NH4D | NH4D +AM | NH4D +P&R | NH4D +H&R | NH4D +FZB | NH4D +AM+FZB | |
|---|---|---|---|---|---|---|---|---|---|
| Total [g] | 233.5 a | 95.6 c | 227.1 a | 87.5 c | 106.9 b | 90.0 c | 85.5 c | 94.5 c | 95.9 c |
| Relative biomass distribution % | |||||||||
| Shoot | 36.7 | 87.4 | 37.9 | 68.8 | 62.1 | 73.2 | 76.3 | 78.2 | 80.5 |
| Tuber | 60.4 | 3.0 | 59.7 | 23.4 | 31.8 | 18.8 | 15.7 | 14.3 | 10.7 |
| Roots | 2.8 | 9.6 | 2.4 | 7.8 | 6.1 | 8.0 | 8.0 | 7.5 | 8.8 |
| (A) | Nutrient Concentrations | P | K | N | Mg | Ca | Mn | Fe | Zn | Cu |
|---|---|---|---|---|---|---|---|---|---|---|
| Deficiency Threshold | 2.5 mg·g−1 | 60 mg·g−1 | 30 mg·g−1 | 7.0 mg·g−1 | 15 mg·g−1 | 30 μg·g−1 | 40 μg·g−1 | 30 μg·g−1 | 6 μg·g−1 | |
| NO3− | Well-watered | 1.19 ± 0.07 f | 24.0 ± 0.7 c | 13.1 ± 0.4 c | 2.88 ± 0.19 cd | 20.3 ± 0.8 a | 29.2 ± 0.5 bcd | 172.4 ± 18.8 a | 56.7 ± 4.6 ab | 5.85 ± 0.9 b |
| Drought stress | 2.19 ± 0.09 a | 39.6 ± 1.7 ab | 30.5 ± 1.2 a | 3.31 ± 0.16 abc | 18.3 ± 0.8 abc | 32.4 ± 0.8 ab | 175.7 ± 16.7 a | 48.9 ± 2.5 ab | 9.29 ± 0.9 a | |
| NH4+ | Well-watered | 1.32 ± 0.13 ef | 21.6 ± 0.5 c | 12.5 ± 0.5 c | 2.71 ± 0.11 d | 17.2 ± 0.9 bc | 26.1 ± 1.2 d | 187.8 ± 22.8 a | 60.1 ± 7.2 a | 7.15 ± 1.1 ab |
| Drought stress | 1.81 ± 0.08 cd | 41.4 ± 1.7 a | 30.8 ± 1.3 a | 3.48 ± 0.14 ab | 19.2 ± 1.0 ab | 32.5 ± 1.4 ab | 171.6 ± 21.5 a | 53.5 ± 3.2 ab | 8.73 ± 0.7 ab | |
| AM | 1.54 ± 0.05 de | 38.2 ± 0.6 ab | 28.7 ± 0.9 ab | 3.59 ± 0.09 ab | 20.1 ± 0.4 a | 33.2 ± 0.4 a | 168.1 ± 15.5 a | 57.5 ± 5.2 ab | 7.72 ± 0.7 ab | |
| AM+3Re2-7 | 1.89 ± 0.06 bc | 40.6 ± 1.9 a | 31.1 ± 1.4 a | 3.58 ± 0.19 ab | 19.5 ± 1.3 ab | 32.6 ± 1.7 ab | 162.6 ± 14.7 a | 57.8 ± 5.4 ab | 9.57 ± 2.2 a | |
| AM+Her | 2.11 ± 0.15 ab | 40.4 ± 2.4 a | 31.2 ± 1.5 a | 3.66 ± 0.22 a | 19.2 ± 1.5 ab | 30.5 ± 1.0 abc | 156.2 ± 23.9 a | 54.4 ± 5.7 ab | 9.00 ± 1.1 a | |
| FZB | 1.87 ± 0.12 bc | 35.8 ± 0.5 b | 26.8 ± 1.0 b | 2.98 ± 0.16 cd | 16.0 ± 1.2 c | 29.1 ± 1.8 bcd | 162.3 ± 13.4 a | 50.5 ± 2.4 ab | 8.56 ± 0.8 ab | |
| AM+FZB | 1.92 ± 0.10 abc | 36.0 ± 0.7 b | 27.2 ± 0.7 b | 3.16 ± 0.08 bcd | 16.1 ± 0.7 c | 28.8 ± 1.5 cd | 146.6 ± 11.6 a | 46.7 ± 2.5 b | 8.89 ± 0.6 ab | |
| (B) | Nutrient contents | Macronutrients (mg·plant−1) | Micronutrients(μg·plant−1) | |||||||
| P | K | N | Mg | Ca | Mn | Fe | Zn | Cu | ||
| NO3− | Well-watered | 16.5 ± 1.1 bcd | 333.7 ± 14.2 cd | 181.9 ± 6.6 d | 39.6 ± 1.4 a | 280.2 ± 6.0 a | 406 ± 19 a | 2373 ± 205 a | 792.5 ± 85.4 a | 80.7 ± 11.3 ab |
| Drought stress | 25.7 ± 0.8 a | 463.0 ± 10.9 a | 357.0 ± 6.2 a | 38.7 ± 0.9 ab | 214.4 ± 5.2 c | 380 ± 8 ab | 2065 ± 210 ab | 573.9 ± 31.2 b | 109.5 ± 12.0 a | |
| NH4+ | Well-watered | 18.3 ± 3.0 bcd | 294.6 ± 19.2 d | 170.2 ± 10.1 d | 36.7 ± 1.6 abc | 231.6 ± 5.4 b | 352.5 ± 13 bc | 2482 ± 143 a | 798.1 ± 58.4 a | 95.1 ± 11.1 ab |
| Drought stress | 15.9 ± 1.8 cd | 358.3 ± 21.1 bc | 266.6 ± 15.5 c | 30.0 ± 1.3 e | 164.3 ± 4.3 e | 280 ± 10 e | 1491 ± 206 c | 465.6 ± 41.7 b | 74.7 ± 4.9 b | |
| AM * | 14.4 ± 0.9 d | 356.5 ± 16.4 bc | 267.3 ± 7.7 c | 33.5 ± 1.2 cde | 187.5 ± 7.3 d | 311 ± 18 cde | 1593 ± 210 c | 537.5 ± 55.4 b | 72.3 ± 4.9 b | |
| AM+3Re2-7 | 17.3 ± 1.6 bcd | 366.2 ± 20.0 bc | 280.4 ± 12.9 bc | 32.1 ± 0.9 de | 174.0 ± 2.6 de | 293 ± 11 de | 1441 ± 29 c | 520.6 ± 46.3 b | 85.2 ± 16.3 ab | |
| AM+Her | 20.1 ± 3.1 abcd | 368.8 ± 21.9 bc | 286.5 ± 21.1 bc | 33.4 ± 1.9 cde | 174.1 ± 7.7 de | 282 ± 22 e | 1367 ± 60 c | 498.4 ± 48.9 b | 82.4 ± 9.1 ab | |
| FZB | 21.1 ± 2.8 abc | 396.0 ± 29.6 b | 294.3 ± 16.2 bc | 32.5 ± 1.1 de | 173.2 ± 4.1 de | 317 ± 10 cde | 1777 ± 147 bc | 554.3 ± 32.4 b | 94.6 ± 11.2 ab | |
| AM+FZB | 21.7 ± 1.2 ab | 406.3 ± 6.2 b | 307.1 ± 9.0 b | 35.6 ± 0.5 bcd | 181.0 ± 4.3 d | 324 ± 12 cd | 1642 ± 99 bc | 525.4 ± 21.4 b | 100.2 ± 6.4 ab | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mamun, A.A.; Neumann, G.; Moradtalab, N.; Ahmed, A.; Dupuis, B.; Darbon, G.; Nawaz, F.; Declerck, S.; Mai, K.; Vogt, W.; et al. Microbial Consortia Versus Single-Strain Inoculants as Drought Stress Protectants in Potato Affected by the Form of N Supply. Horticulturae 2024, 10, 102. https://doi.org/10.3390/horticulturae10010102
Mamun AA, Neumann G, Moradtalab N, Ahmed A, Dupuis B, Darbon G, Nawaz F, Declerck S, Mai K, Vogt W, et al. Microbial Consortia Versus Single-Strain Inoculants as Drought Stress Protectants in Potato Affected by the Form of N Supply. Horticulturae. 2024; 10(1):102. https://doi.org/10.3390/horticulturae10010102
Chicago/Turabian StyleMamun, Abdullah Al, Günter Neumann, Narges Moradtalab, Aneesh Ahmed, Brice Dupuis, Geoffrey Darbon, Fahim Nawaz, Stephane Declerck, Karin Mai, Wolfgang Vogt, and et al. 2024. "Microbial Consortia Versus Single-Strain Inoculants as Drought Stress Protectants in Potato Affected by the Form of N Supply" Horticulturae 10, no. 1: 102. https://doi.org/10.3390/horticulturae10010102
APA StyleMamun, A. A., Neumann, G., Moradtalab, N., Ahmed, A., Dupuis, B., Darbon, G., Nawaz, F., Declerck, S., Mai, K., Vogt, W., Ludewig, U., & Weinmann, M. (2024). Microbial Consortia Versus Single-Strain Inoculants as Drought Stress Protectants in Potato Affected by the Form of N Supply. Horticulturae, 10(1), 102. https://doi.org/10.3390/horticulturae10010102