

Exploitation of Cocoa Pod Residues for the Production of Antioxidants, Polyhydroxyalkanoates, and Ethanol

 , ,
, ,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Feedstocks and Chemicals
2.2. Chemical Analyses of CPH and CBS
2.3. Phenolic Extraction of Cocoa Pods and Shells
2.4. Quantification of the Total Phenolic Compounds (TPC)
2.5. Phenolic Compounds Identification
2.6. Antioxidant Activity
2.7. Determination of the Oxidative Stability
2.8. Pretreatments and Enzymatic Hydrolysis
2.9. Microbial Strains
2.10. PHAs Production by C. necator DSM 545 from Alkaline Pretreated CPH Hydrolyzate (Ahs-CPH)
2.11. Cell Dry Matter and PHAs Analysis
2.12. Bioethanol Production by S. cerevisiae Fm17 from Ahs-CPH
2.13. Statistical Analysis
3. Results and Discussion
3.1. Chemical Composition of Cocoa Pods Husk
3.2. Quantification and Identification of Total Phenolic Compounds (TPC) and Antioxidant Activity
3.3. Oxidative Stability
3.4. Pretreatment of Cocoa Residues
3.4.1. Hydrogen Peroxide (H2O2) Pretreatment of CPH
3.4.2. Alkaline (NaOH) Pretreatment of CPH
3.5. PHAs and Bioethanol Production from Alkaline Pretreated Saccharified CPH (Ahs-CPH)
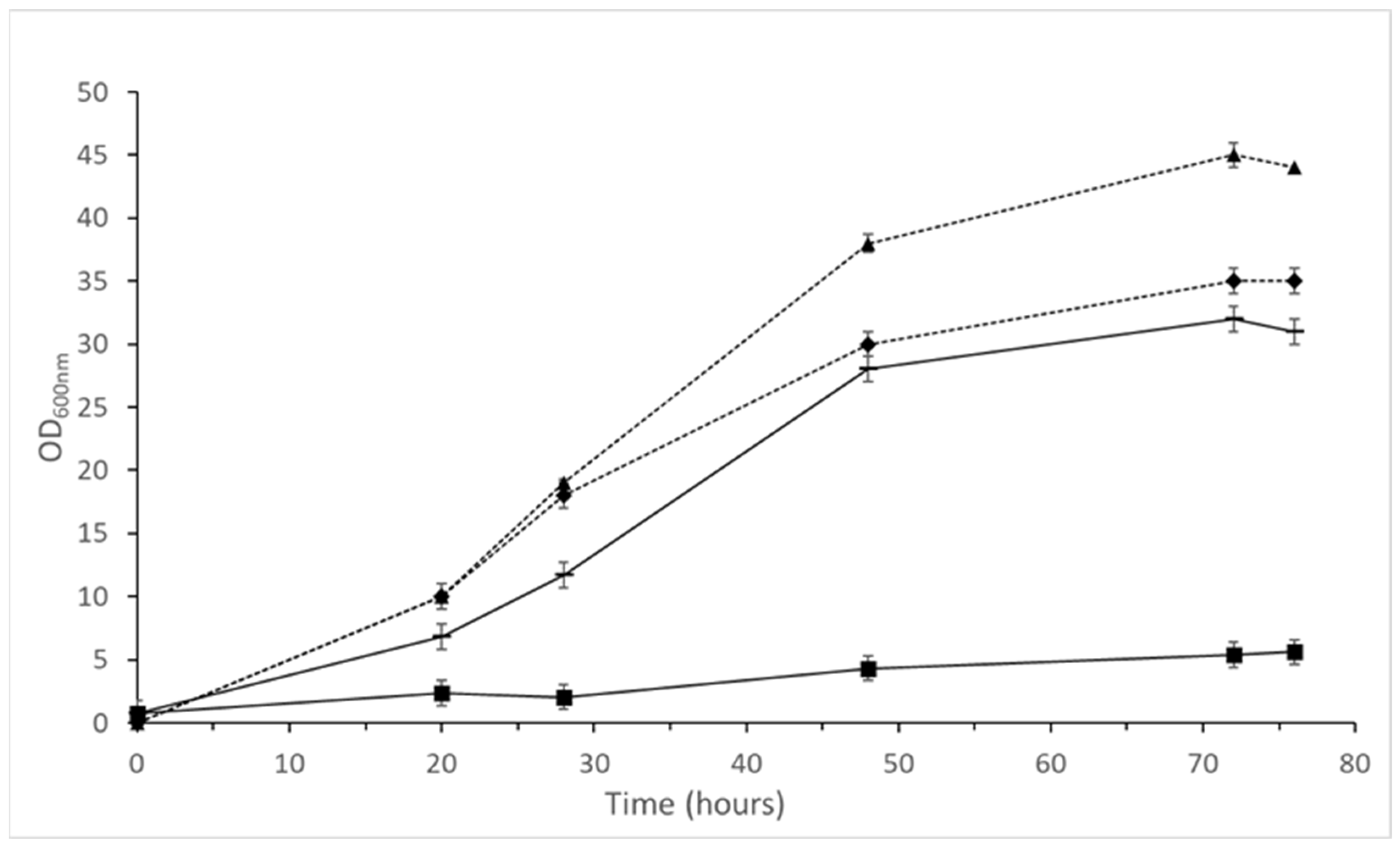
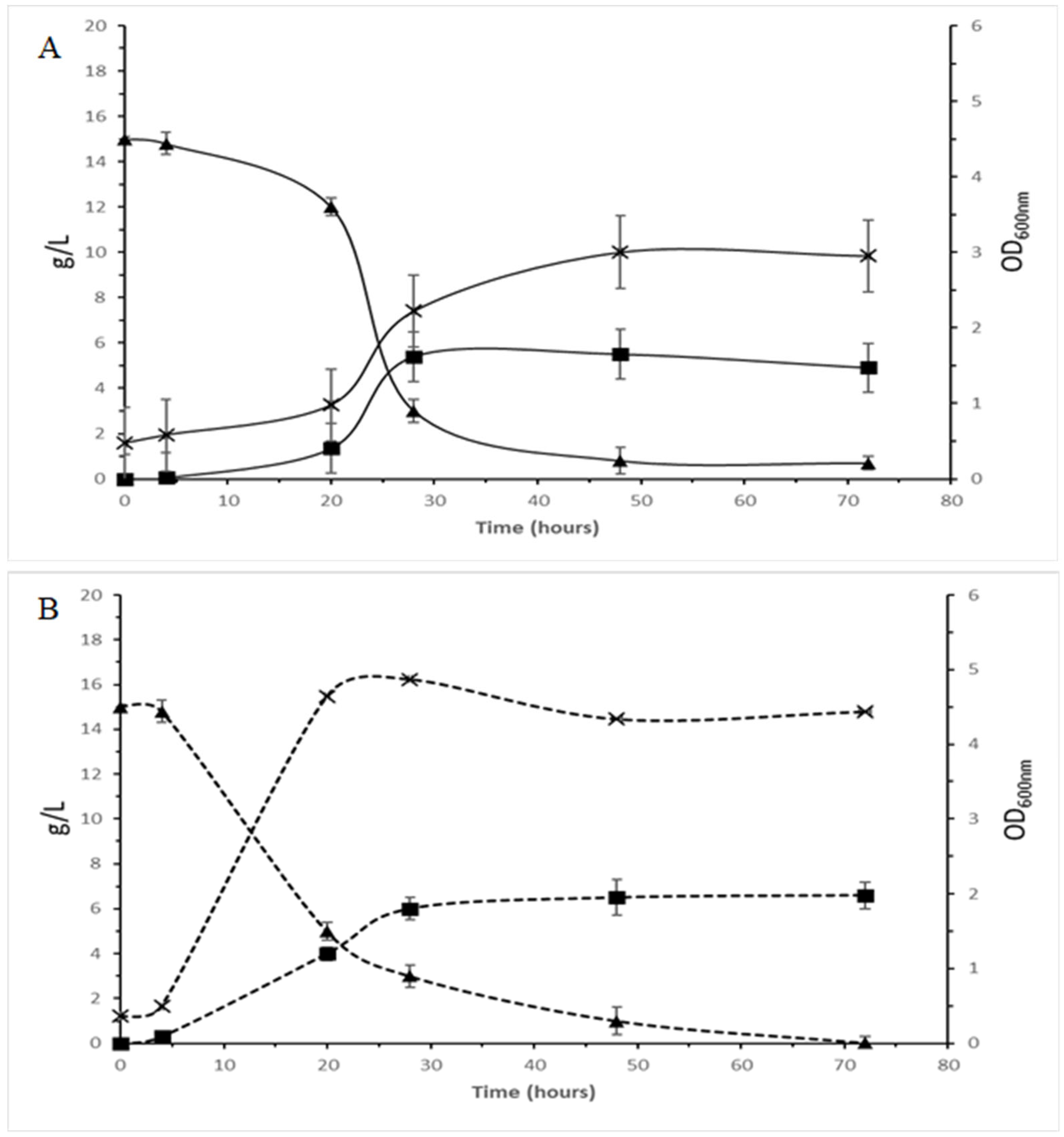
3.5.1. PHAs
3.5.2. Bioethanol
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Soares, T.; Oliveira, M.B. Cocoa By-Products: Characterization of Bioactive Compounds and Beneficial Health Effects. Molecules 2022, 27, 1625. [Google Scholar] [CrossRef] [PubMed]
- ICCO. ICCO Cocoa Year 2021/22. Q. Bull. Cocoa Stat. 2022, XLVIII, 1. [Google Scholar]
- Daymond, A.; Bekele, F. Cacao. In Cash Crops; Springer: Cham, Switzerland, 2022; pp. 23–53. [Google Scholar]
- Afoakwa, E.O.; Paterson, A.; Fowler, M.; Ryan, A. Flavor Formation and Character in Cocoa and Chocolate: A Critical Review. Crit. Rev. Food Sci. Nutr. 2008, 48, 840–857. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Rodriguez-Garcia, J.; Van Damme, I.; Westwood, N.J.; Shaw, L.; Robinson, J.S.; Warren, G.; Chatzifragkou, A.; McQueen Mason, S.; Gomez, L.; et al. Valorisation Strategies for Cocoa Pod Husk and Its Fractions. Curr. Opin. Green Sustain. Chem. 2018, 14, 80–88. [Google Scholar] [CrossRef]
- Beckett, S. The Science of Chocolate, 3rd ed.; RSC Publishing: Cambridge, UK, 2001; ISBN 9780854049707. [Google Scholar]
- Vásquez, Z.S.; de Carvalho Neto, D.P.; Pereira, G.V.M.; Vandenberghe, L.P.S.; de Oliveira, P.Z.; Tiburcio, P.B.; Rogez, H.L.G.; Góes Neto, A.; Soccol, C.R. Biotechnological Approaches for Cocoa Waste Management: A Review. Waste Manag. 2019, 90, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Vriesmann, L.C.; de Mello Castanho Amboni, R.D.; de Oliveira Petkowicz, C.L. Cacao Pod Husks (Theobroma cacao L.): Composition and Hot-Water-Soluble Pectins. Ind. Crops Prod. 2011, 34, 1173–1181. [Google Scholar] [CrossRef]
- Oddoye, E.O.K.; Agyente-Badu, C.K.; Gyedu-Akoto, E. Cocoa and Its By-Products: Identification and Utilization. In Chocolate in Health and Nutrition. Nutrition and Health; Humana Press: Totowa, NJ, USA, 2013; Volume 7, pp. 23–37. [Google Scholar] [CrossRef]
- Acosta, N.; De Vrieze, J.; Sandoval, V.; Sinche, D.; Wierinck, I.; Rabaey, K. Cocoa Residues as Viable Biomass for Renewable Energy Production through Anaerobic Digestion. Bioresour. Technol. 2018, 265, 568–572. [Google Scholar] [CrossRef] [PubMed]
- Antwi, E.; Engler, N.; Narra, S.; Schüch, A.; Nelles, M. Environmental Effect of Cocoa Pods Disposal in 3 West African Countries. In 13th Rostock Bioenergy Forum Proceedings; Rostock University: Rostock, Germany, 2019; 463p. [Google Scholar]
- Hougni, D.-G.J.M.; Schut, A.G.T.; Woittiez, L.S.; Vanlauwe, B.; Giller, K.E. How Nutrient Rich Are Decaying Cocoa Pod Husks? The Kinetics of Nutrient Leaching. Plant Soil 2021, 463, 155–170. [Google Scholar] [CrossRef]
- Kley Valladares-Diestra, K.; Porto de Souza Vandenberghe, L.; Ricardo Soccol, C. A Biorefinery Approach for Pectin Extraction and Second-Generation Bioethanol Production from Cocoa Pod Husk. Bioresour. Technol. 2022, 346, 126635. [Google Scholar] [CrossRef]
- Muñoz-Almagro, N.; Valadez-Carmona, L.; Mendiola, J.A.; Ibáñez, E.; Villamiel, M. Structural Characterisation of Pectin Obtained from Cacao Pod Husk. Comparison of Conventional and Subcritical Water Extraction. Carbohydr. Polym. 2019, 217, 69–78. [Google Scholar] [CrossRef]
- Porto de Souza Vandenberghe, L.; Kley Valladares-Diestra, K.; Amaro Bittencourt, G.; Fátima Murawski de Mello, A.; Sarmiento Vásquez, Z.; Zwiercheczewski de Oliveira, P.; Vinícius de Melo Pereira, G.; Ricardo Soccol, C. Added-Value Biomolecules’ Production from Cocoa Pod Husks: A Review. Bioresour. Technol. 2022, 344, 126252. [Google Scholar] [CrossRef] [PubMed]
- United Nations The 17 Goals. Available online: https://sdgs.un.org/goals (accessed on 1 August 2023).
- Yadav, A.; Sharma, V.; Tsai, M.L.; Chen, C.W.; Sun, P.P.; Nargotra, P.; Wang, J.X.; Dong, C. Di Development of Lignocellulosic Biorefineries for the Sustainable Production of Biofuels: Towards Circular Bioeconomy. Bioresour. Technol. 2023, 381, 129145. [Google Scholar] [CrossRef] [PubMed]
- Mujtaba, M.; Fernandes Fraceto, L.; Fazeli, M.; Mukherjee, S.; Savassa, S.M.; Araujo de Medeiros, G.; do Espírito Santo Pereira, A.; Mancini, S.D.; Lipponen, J.; Vilaplana, F. Lignocellulosic Biomass from Agricultural Waste to the Circular Economy: A Review with Focus on Biofuels, Biocomposites and Bioplastics. J. Clean. Prod. 2023, 402, 136815. [Google Scholar] [CrossRef]
- Sánchez, M.; Laca, A.; Laca, A.; Díaz, M. Cocoa Bean Shell: A By-Product with High Potential for Nutritional and Biotechnological Applications. Antioxidants 2023, 12, 1028. [Google Scholar] [CrossRef] [PubMed]
- Cinar, Z.Ö.; Atanassova, M.; Tumer, T.B.; Caruso, G.; Antika, G.; Sharma, S.; Sharifi-Rad, J.; Pezzani, R. Cocoa and Cocoa Bean Shells Role in Human Health: An Updated Review. J. Food Compos. Anal. 2021, 103, 104115. [Google Scholar] [CrossRef]
- Koller, M.; Mukherjee, A. A New Wave of Industrialization of PHA Biopolyesters. Bioengineering 2022, 9, 74. [Google Scholar] [CrossRef] [PubMed]
- Cadariu, A.I.; Cocan, I.; Negrea, M.; Alexa, E.; Obistioiu, D.; Hotea, I.; Radulov, I.; Poiana, M.A. Exploring the Potential of Tomato Processing Byproduct as a Natural Antioxidant in Reformulated Nitrite-Free Sausages. Sustainability 2022, 14, 11802. [Google Scholar] [CrossRef]
- Atiwesh, G.; Mikhael, A.; Parrish, C.C.; Banoub, J.; Le, T.A.T. Environmental Impact of Bioplastic Use: A Review. Heliyon 2021, 7, e07918. [Google Scholar] [CrossRef]
- Berezina, N. Novel Approach for Productivity Enhancement of Polyhydroxyalkanoates (PHA) Production by Cupriavidus necator DSM 545. New Biotechnol. 2013, 30, 192–195. [Google Scholar] [CrossRef]
- Available online: https://www.european-bioplastics.org/market/ (accessed on 1 August 2023).
- Favaro, L.; Basaglia, M.; Casella, S. Improving Polyhydroxyalkanoate Production from Inexpensive Carbon Sources by Genetic Approaches: A Review. Biofuels Bioprod. Biorefining 2019, 13, 208–227. [Google Scholar] [CrossRef]
- Favaro, L.; Basaglia, M.; Rodriguez, J.E.G.; Morelli, A.; Ibraheem, O.; Pizzocchero, V.; Casella, S. Bacterial Production of PHAs from Lipid-Rich by-Products. Appl. Food Biotechnol. 2019, 6, 45–52. [Google Scholar] [CrossRef]
- Yukesh Kannah, R.; Dinesh Kumar, M.; Kavitha, S.; Rajesh Banu, J.; Kumar Tyagi, V.; Rajaguru, P.; Kumar, G. Production and Recovery of Polyhydroxyalkanoates (PHA) from Waste Streams—A Review. Bioresour. Technol. 2022, 366, 128203. [Google Scholar] [CrossRef] [PubMed]
- International Energy Agency. World Energy Outlook 2010; Birol, F., Ed.; International Energy Agency: Paris, France, 2010; Available online: https://iea.blob.core.windows.net/assets/1b090169-1c58-4f5d-9451-ee838f6f00e5/weo2010.pdf (accessed on 1 August 2023).
- Sánchez, M.; Laca, A.; Laca, A.; Díaz, M. Cocoa Bean Shell as Promising Feedstock for the Production of Poly(3-Hydroxybutyrate) (PHB). Appl. Sci. 2023, 13, 975. [Google Scholar] [CrossRef]
- Samah, O.A.; Sias, S.; Hua, Y.G.; Hussin, N.N. Production of Ethanol from Cocoa Pod Hydrolysate. J. Math. Fundam. Sci. 2011, 43, 87–94. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, P.; Mihaylova, D.; Marangon, C.M.; Grigoletto, L.; Lante, A. Impact of Sample Pretreatment and Extraction Methods on the Bioactive Compounds of Sugar Beet (Beta vulgaris L.) Leaves. Molecules 2022, 27, 8110. [Google Scholar] [CrossRef] [PubMed]
- Amare, E.; Grigoletto, L.; Corich, V.; Giacomini, A.; Lante, A. Fatty Acid Profile, Lipid Quality and Squalene Content of Teff (Eragrostis teff (Zucc.) Trotter) and Amaranth (Amaranthus caudatus L.) Varieties from Ethiopia. Appl. Sci. 2021, 11, 3590. [Google Scholar] [CrossRef]
- Cisneros-Yupanqui, M.; Chalova, V.I.; Kalaydzhiev, H.R.; Mihaylova, D.; Krastanov, A.I.; Lante, A. Preliminary Characterisation of Wastes Generated from the Rapeseed and Sunflower Protein Isolation Process and Their Valorisation in Delaying Oil Oxidation. Food Bioprocess Technol. 2021, 1, 1962–1971. [Google Scholar] [CrossRef]
- Stratil, P.; Kubáň, V.; Fojtová, J. Comparison of the Phenolic Content and Total Antioxidant Activity in Wines as Determined by Spectrophotometric Methods. Czech J. Food Sci. 2008, 26, 242–253. [Google Scholar] [CrossRef]
- Tinello, F.; Lante, A.; Bernardi, M.; Cappiello, F.; Galgano, F.; Caruso, M.C.; Favati, F. Comparison of OXITEST and RANCIMAT Methods to Evaluate the Oxidative Stability in Frying Oils. Eur. Food Res. Technol. 2018, 244, 747–755. [Google Scholar] [CrossRef]
- Hernández-Mendoza, A.G.; Saldaña-Trinidad, S.; Martínez-Hernández, S.; Pérez-Sariñana, B.Y.; Láinez, M. Optimization of Alkaline Pretreatment and Enzymatic Hydrolysis of Cocoa Pod Husk (Theobroma cacao L.) for Ethanol Production. Biomass Bioenergy 2021, 154, 106268. [Google Scholar] [CrossRef]
- Brojanigo, S.; Gronchi, N.; Cazzorla, T.; Wong, T.S.; Basaglia, M.; Favaro, L.; Casella, S. Engineering Cupriavidus Necator DSM 545 for the One-Step Conversion of Starchy Waste into Polyhydroxyalkanoates. Bioresour. Technol. 2022, 347, 126383. [Google Scholar] [CrossRef] [PubMed]
- Cagnin, L.; Gronchi, N.; Basaglia, M.; Favaro, L.; Casella, S. Selection of Superior Yeast Strains for the Fermentation of Lignocellulosic Steam-Exploded Residues. Front. Microbiol. 2021, 12, 756032. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, B.A.; Lomaliza, K.; Chavarie, C.; Dube, B.; Bataille, P.; Ramsay, J.A. Production of Poly-(Beta-Hydroxybutyric-Co-Beta-Hydroxyvaleric) Acids. Appl. Environ. Microbiol. 1990, 56, 2093. [Google Scholar] [CrossRef] [PubMed]
- Braunegg, G.; Sonnleitner, B.; Lafferty, R.M. A Rapid Gas Chromatographic Method for the Determination of Poly-b-Hydroxybutyric Acid in Microbial Biomass. Eur. J. Appl. Microbiol. Biotechnol. 1978, 6, 29–37. [Google Scholar] [CrossRef]
- Rodríguez Gamero, J.E.; Favaro, L.; Pizzocchero, V.; Lomolino, G.; Basaglia, M.; Casella, S. Nuclease Expression in Efficient Polyhydroxyalkanoates-Producing Bacteria Could Yield Cost Reduction during Downstream Processing. Bioresour. Technol. 2018, 261, 176–181. [Google Scholar] [CrossRef]
- Cagnin, L.; Favaro, L.; Gronchi, N.; Rose, S.H.; Basaglia, M.; van Zyl, W.H.; Casella, S. Comparing Laboratory and Industrial Yeast Platforms for the Direct Conversion of Cellobiose into Ethanol under Simulated Industrial Conditions. FEMS Yeast Res. 2019, 19, foz018. [Google Scholar] [CrossRef]
- Mendoza-Meneses, C.J.; Feregrino-Pérez, A.A.; Gutiérrez-Antonio, C. Potential Use of Industrial Cocoa Waste in Biofuel Production. J. Chem. 2021, 2021, 3388067. [Google Scholar] [CrossRef]
- Coimbra, M.C.; Jorge, N. Proximate Composition of Guariroba (Syagrus oleracea), Jerivá (Syagrus romanzoffiana) and Macaúba (Acrocomia aculeata) Palm Fruits. Food Res. Int. 2011, 44, 2139–2142. [Google Scholar] [CrossRef]
- Martínez, R.; Torres, P.; Meneses, M.A.; Figueroa, J.G.; Pérez-Álvarez, J.A.; Viuda-Martos, M. Chemical, Technological and in Vitro Antioxidant Properties of Cocoa (Theobroma cacao L.) Co-Products. Food Res. Int. 2012, 49, 39–45. [Google Scholar] [CrossRef]
- Campione, A.; Pauselli, M.; Natalello, A.; Valenti, B.; Pomente, C.; Avondo, M.; Luciano, G.; Caccamo, M.; Morbidini, L. Inclusion of Cocoa By-Product in the Diet of Dairy Sheep: Effect on the Fatty Acid Profile of Ruminal Content and on the Composition of Milk and Cheese. Animal 2021, 15, 100243. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Barreto, J.F.; Larrea, F.; Pinos, M.C.; Benalcázar, J.; Oña, D.; Andino, C.; Viteri, D.A.; Leon, M.; Almeida-Streitwieser, D. Chemical Pretreatments on Residual Cocoa Pod Shell Biomass for Bioethanol Production. Bionatura 2021, 6, 1490–1500. [Google Scholar] [CrossRef]
- Botella-Martínez, C.; Lucas-Gonzalez, R.; Ballester-Costa, C.; Pérez-álvarez, J.Á.; Fernández-López, J.; Delgado-Ospina, J.; Chaves-López, C.; Viuda-Martos, M. Ghanaian Cocoa (Theobroma cacao L.) Bean Shells Coproducts: Effect of Particle Size on Chemical Composition, Bioactive Compound Content and Antioxidant Activity. Agronomy 2021, 11, 401. [Google Scholar] [CrossRef]
- Rojo-Poveda, O.; Barbosa-Pereira, L.; Mateus-Reguengo, L.; Bertolino, M.; Stévigny, C.; Zeppa, G. Effects of Particle Size and Extraction Methods on Cocoa Bean Shell Functional Beverage. Nutrients 2019, 11, 867. [Google Scholar] [CrossRef] [PubMed]
- Donkoh, A.; Atuahene, C.C.; Wilson, B.N.; Adomako, D. Chemical Composition of Cocoa Pod Husk and Its Effect on Growth and Food Efficiency in Broiler Chicks. Anim. Feed Sci. Technol. 1991, 35, 161–169. [Google Scholar] [CrossRef]
- Serra Bonvehí, J.; Ventura Coll, F. Protein Quality Assessment in Cocoa Husk. Food Res. Int. 1999, 32, 201–208. [Google Scholar] [CrossRef]
- Lessa, O.A.; dos Santos Reis, N.; Leite, S.G.F.; Gutarra, M.L.E.; Souza, A.O.; Gualberto, S.A.; de Oliveira, J.R.; Aguiar-Oliveira, E.; Franco, M. Effect of the Solid State Fermentation of Cocoa Shell on the Secondary Metabolites, Antioxidant Activity, and Fatty Acids. Food Sci. Biotechnol. 2018, 27, 107–113. [Google Scholar] [CrossRef]
- Soares, I.D.; Okiyama, D.C.G.; da Costa Rodrigues, C.E. Simultaneous Green Extraction of Fat and Bioactive Compounds of Cocoa Shell and Protein Fraction Functionalities Evaluation. Food Res. Int. 2020, 137, 109622. [Google Scholar] [CrossRef]
- Adjin-Tetteh, M.; Asiedu, N.; Dodoo-Arhin, D.; Karam, A.; Amaniampong, P.N. Thermochemical Conversion and Characterization of Cocoa Pod Husks a Potential Agricultural Waste from Ghana. Ind. Crops Prod. 2018, 119, 304–312. [Google Scholar] [CrossRef]
- Adewole, E.; Ajiboye, B.; Ojo, B.; Ogunmodede, O.; Oso, O. Characterization of Cocoa (Theobroma cocoa) Pod. J. Chem. Inf. Model. 2013, 4. [Google Scholar]
- Abdul Karim, A.; Azlan, A.; Ismail, A.; Hashim, P.; Abd Gani, S.S.; Zainudin, B.H.; Abdullah, N.A. Phenolic Composition, Antioxidant, Anti-Wrinkles and Tyrosinase Inhibitory Activities of Cocoa Pod Extract. BMC Complement. Altern. Med. 2014, 14, 381. [Google Scholar] [CrossRef] [PubMed]
- Dewi, S.R.; Stevens, L.A.; Pearson, A.E.; Ferrari, R.; Irvine, D.J.; Binner, E.R. Investigating the Role of Solvent Type and Microwave Selective Heating on the Extraction of Phenolic Compounds from Cacao (Theobroma cacao L.) Pod Husk. Food Bioprod. Process. 2022, 134, 210–222. [Google Scholar] [CrossRef]
- Rachmawaty; Mu’Nisa, A.; Hasri; Pagarra, H.; Hartati; Maulana, Z. Active Compounds Extraction of Cocoa Pod Husk (Thebroma cacao L.) and Potential as Fungicides. J. Phys. Conf. Ser. 2018, 1028, 012013. [Google Scholar] [CrossRef]
- Sotelo, L.; Alvis, A.; Arrázola, G. Evaluation of Epicatechin, Theobromine and Caffeine in Cacao Husks (Theobroma cacao L.), Determination of the Antioxidant Capacity. Rev. Colomb. Cienc. Hortícolas 2015, 9, 124–134. [Google Scholar] [CrossRef]
- Valadez-Carmona, L.; Plazola-Jacinto, C.P.; Hernández-Ortega, M.; Hernández-Navarro, M.D.; Villarreal, F.; Necoechea-Mondragón, H.; Ortiz-Moreno, A.; Ceballos-Reyes, G. Effects of Microwaves, Hot Air and Freeze-Drying on the Phenolic Compounds, Antioxidant Capacity, Enzyme Activity and Microstructure of Cacao Pod Husks (Theobroma cacao L.). Innov. Food Sci. Emerg. Technol. 2017, 41, 378–386. [Google Scholar] [CrossRef]
- Bortolini, C.; Patrone, V.; Puglisi, E.; Morelli, L. Detailed Analyses of the Bacterial Populations in Processed Cocoa Beans of Different Geographic Origin, Subject to Varied Fermentation Conditions. Int. J. Food Microbiol. 2016, 236, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Campos-Vega, R.; Nieto-Figueroa, K.H.; Oomah, B.D. Cocoa (Theobroma cacao L.) Pod Husk: Renewable Source of Bioactive Compounds. Trends Food Sci. Technol. 2018, 81, 172–184. [Google Scholar] [CrossRef]
- Boungo Teboukeu, G.; Tonfack Djikeng, F.; Klang, M.J.; Houketchang Ndomou, S.; Karuna, M.S.L.; Womeni, H.M. Polyphenol Antioxidants from Cocoa Pods: Extraction Optimization, Effect of the Optimized Extract, and Storage Time on the Stability of Palm Olein during Thermoxidation. J. Food Process. Preserv. 2018, 42, e13592. [Google Scholar] [CrossRef]
- Lante, A.; Nardi, T.; Zocca, F.; Giacomini, A.; Corich, V. Evaluation of Red Chicory Extract as a Natural Antioxidant by Pure Lipid Oxidation and Yeast Oxidative Stress Response as Model Systems. J. Agric. Food Chem. 2011, 59, 5318–5324. [Google Scholar] [CrossRef]
- Cisneros-Yupanqui, M.; Zagotto, A.; Alberton, A.; Lante, A.; Zagotto, G.; Ribaudo, G.; Rizzi, C. Study of the Phenolic Profile of a Grape Pomace Powder and Its Impact on Delaying Corn Oil Oxidation. Nat. Prod. Res. 2020, 36, 455–459. [Google Scholar] [CrossRef]
- Mensah, M.; Yaw Asiedu, N.; Abunde Neba, F.; Nana Amaniampong, P.; Boakye, P.; Addo, A. Modeling, Optimization and Kinetic Analysis of the Hydrolysis Process of Waste Cocoa Pod Husk to Reducing Sugars. SN Appl. Sci. 2020, 2, 1160. [Google Scholar] [CrossRef]
- García, Y.C.; Morales, J.J.; Montalvo, P.A.J.; Figueroa, L.V. Pretratamiento e Hidrólisis Enzimática de La Cascarilla de Arroz. Rev. Soc. Quím. Perú 2019, 85, 476–488. [Google Scholar] [CrossRef]
- Benalcazar Bassante, J.C. Evaluación de Diferentes Pretratamientos Químicos a la Biomasa de la Cáscara de Cacao Para Procesos de Fermentación Alcohólica; Universidad San Francisco de Quito: Quito, Ecuador, 2018. [Google Scholar]
- Sarmiento-Vásquez, Z.; Vandenberghe, L.; Rodrigues, C.; Tanobe, V.O.A.; Marín, O.; de Melo Pereira, G.V.; Ghislain Rogez, H.L.; Góes-Neto, A.; Soccol, C.R. Cocoa Pod Husk Valorization: Alkaline-Enzymatic Pre-Treatment for Propionic Acid Production. Cellulose 2021, 28, 4009–4024. [Google Scholar] [CrossRef]
- Shet, V.B.; Sanil, N.; Bhat, M.; Naik, M.; Mascarenhas, L.N.; Goveas, L.C.; Rao, C.V.; Ujwal, P.; Sandesh, K.; Aparna, A. Acid Hydrolysis Optimization of Cocoa Pod Shell Using Response Surface Methodology Approach toward Ethanol Production. Agric. Nat. Resour. 2018, 52, 581–587. [Google Scholar] [CrossRef]
- Gutiérrez-Macías, P.; Montañez-Barragán, B.; Barragán-Huerta, B.E. A Review of Agro-Food Waste Transformation into Feedstock for Use in Fermentation. Fresenius Environ. Bull. 2015, 24, 3703–3716. [Google Scholar]
- Brojanigo, S.; Parro, E.; Cazzorla, T.; Favaro, L.; Basaglia, M.; Casella, S. Conversion of Starchy Waste Streams into Polyhydroxyalkanoates Using Cupriavidus Necator DSM 545. Polymers 2020, 12, 1496. [Google Scholar] [CrossRef] [PubMed]
- Brodin, M.; Vallejos, M.; Opedal, M.T.; Area, M.C.; Chinga-Carrasco, G. Lignocellulosics as Sustainable Resources for Production of Bioplastics—A Review. J. Clean. Prod. 2017, 162, 646–664. [Google Scholar] [CrossRef]
- Annamalai, N.; Sivakumar, N. Production of Polyhydroxybutyrate from Wheat Bran Hydrolysate Using Ralstonia Eutropha through Microbial Fermentation. J. Biotechnol. 2016, 237, 13–17. [Google Scholar] [CrossRef]
- de Souza, L.; Manasa, Y.; Shivakumar, S. Bioconversion of Lignocellulosic Substrates for the Production of Polyhydroxyalkanoates. Biocatal. Agric. Biotechnol. 2020, 28, 101754. [Google Scholar] [CrossRef]
- Silva, L.F.; Taciro, M.K.; Michelin Ramos, M.E.; Carter, J.M.; Pradella, J.G.C.; Gomez, J.G.C. Poly-3-Hydroxybutyrate (P3HB) Production by Bacteria from Xylose, Glucose and Sugarcane Bagasse Hydrolysate. J. Ind. Microbiol. Biotechnol. 2004, 31, 245–254. [Google Scholar] [CrossRef]
- Yogaswara, R.R.; Billah, M.; Saputro, E.A.; Erliyanti, N.K. A Kinetic Study in Fermentation of Cocoa Pod Husk Using Zymomonas mobilis. In Proceedings of the IOP Conference Series: Materials Science and Engineering; IOP Publishing: Bristol, UK, 2021. [Google Scholar]
- Favaro, L.; Jansen, T.; van Zyl, W.H. Exploring Industrial and Natural Saccharomyces cerevisiae Strains for the Bio-Based Economy from Biomass: The Case of Bioethanol. Crit. Rev. Biotechnol. 2019, 39, 800–816. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CPH | CBS | |
|---|---|---|
| Cellulose | 22.32 | 12.30 |
| Hemicellulose | 10.10 | 10.07 |
| Acid Detergent Lignin (ADL) | 21.15 | 14.80 |
| Neutral Detergent Fiber (NDF) | 53.85 | 37.91 |
| Acid Detergent Fiber (ADF) | 43.75 | 27.85 |
| Acid Insoluble Ash (AIA) | 0.28 | 0.66 |
| Protein | 4.79 | 17.98 |
| Ashes | 10.41 | 7.28 |
| Lipids | 0.35 | 16.24 |
| Residue | TPC (mg GA/g) | Antioxidant Activity (mg TE/g) |
|---|---|---|
| CPH | 10.08 ± 1.40 a | 9.93 ± 0.38 b |
| CBS | 13.04 ± 0.10 a | 16.24 ± 0.61 a |
| Compound | CBS | CPH |
|---|---|---|
| Pyrogallol | 164.47 | 72.57 |
| Syringic acid | 8.59 | 0.77 |
| p-hydroxybenzoic acid | - | 2.47 |
| Vanillic acid | - | 7.21 |
| Sample | IT (hours) | AAI (mg TE/g) |
|---|---|---|
| CPH | 9.46 ± 0.04 a | 1.48 |
| CPS | 8.45 ± 0.10 b | 1.32 |
| Control (soybean oil) | 6.40 ± 0.06 c | 1.00 |
| Medium | PHAs (% CDM) |
|---|---|
| Ahs-CPH | 51.30 ± 2.83 |
| MM+ Ahs-CPH | 58.60 ± 4.95 |
| MM+ glucose 15 g/L | 72.00 ± 1.00 |
| MM + glucose 20 g/L | 74.60 ± 0.28 |
| Parameter | Growth on Glucose | Growth on Ahs-CPH |
|---|---|---|
| Sugars concentration (g/L) | 15.00 ± 0.08 | 15.15 ± 0.85 |
| Consumed sugars (%) | 100.0 | 95.3 |
| Highest Ethanol (g/L) | 6.60 ± 0.60 | 5.50 ± 0.30 |
| YEtOH/S | 0.44 | 0.38 |
| % theoretical yield | 86 | 74 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos, L.H.; Cisneros-Yupanqui, M.; Santisteban Soto, D.V.; Lante, A.; Favaro, L.; Casella, S.; Basaglia, M. Exploitation of Cocoa Pod Residues for the Production of Antioxidants, Polyhydroxyalkanoates, and Ethanol. Fermentation 2023, 9, 843. https://doi.org/10.3390/fermentation9090843
Ramos LH, Cisneros-Yupanqui M, Santisteban Soto DV, Lante A, Favaro L, Casella S, Basaglia M. Exploitation of Cocoa Pod Residues for the Production of Antioxidants, Polyhydroxyalkanoates, and Ethanol. Fermentation. 2023; 9(9):843. https://doi.org/10.3390/fermentation9090843
Chicago/Turabian StyleRamos, Licelander Hennessey, Miluska Cisneros-Yupanqui, Diana Vanessa Santisteban Soto, Anna Lante, Lorenzo Favaro, Sergio Casella, and Marina Basaglia. 2023. "Exploitation of Cocoa Pod Residues for the Production of Antioxidants, Polyhydroxyalkanoates, and Ethanol" Fermentation 9, no. 9: 843. https://doi.org/10.3390/fermentation9090843
APA StyleRamos, L. H., Cisneros-Yupanqui, M., Santisteban Soto, D. V., Lante, A., Favaro, L., Casella, S., & Basaglia, M. (2023). Exploitation of Cocoa Pod Residues for the Production of Antioxidants, Polyhydroxyalkanoates, and Ethanol. Fermentation, 9(9), 843. https://doi.org/10.3390/fermentation9090843