
Exploring the Fermentation-Driven Functionalities of Lactobacillaceae-Originated Probiotics in Preventive Measures of Alzheimer’s Disease: A Review


 , ,
, ,  and
and 
Abstract
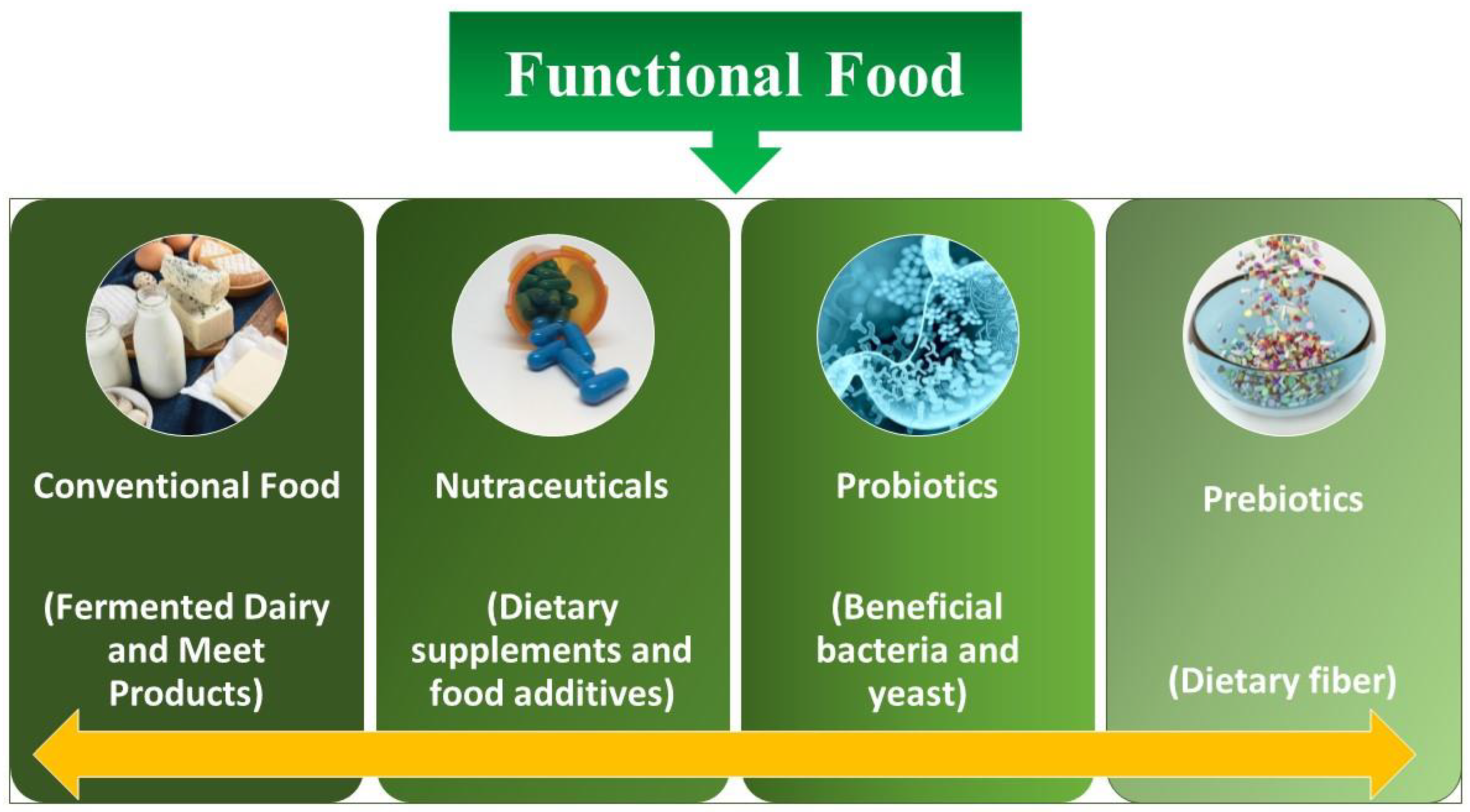
:1. Introduction
Role of Psychobiotics in the Improving Mental Health
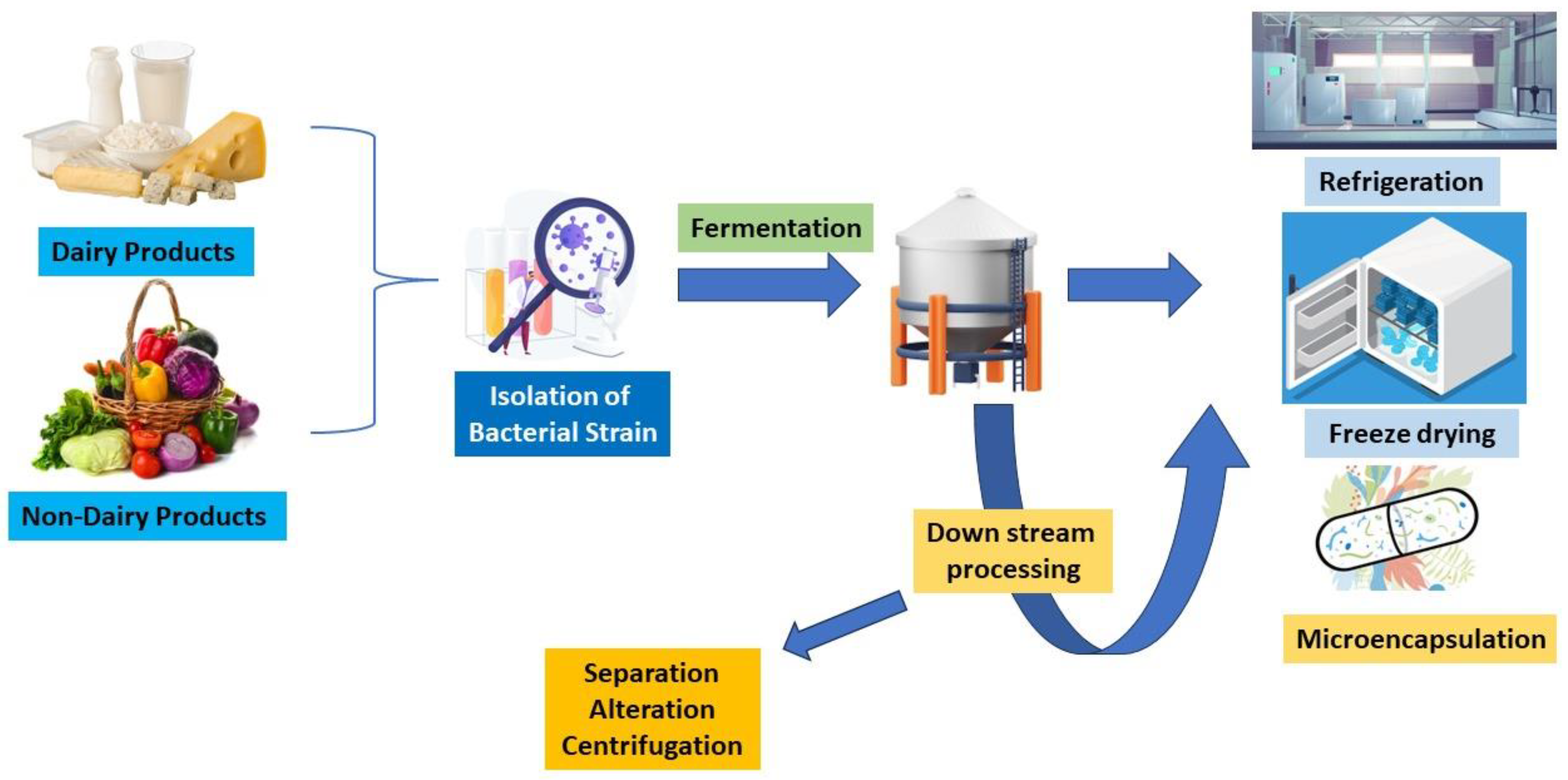
2. Production of Probiotics from Dairy and Non-Dairy Products
3. Comparison of Probiotics Originated from Lactobacilli and Other Strains
4. The Effects of Fermentation on Probiotic-Generating Microbiota
4.1. Increased Viability
4.2. Metabolic Activity
4.3. Increased Availability of Nutrients
4.4. Bioactive Molecule Synthesis
5. Importance of Probiotics for Mental Health
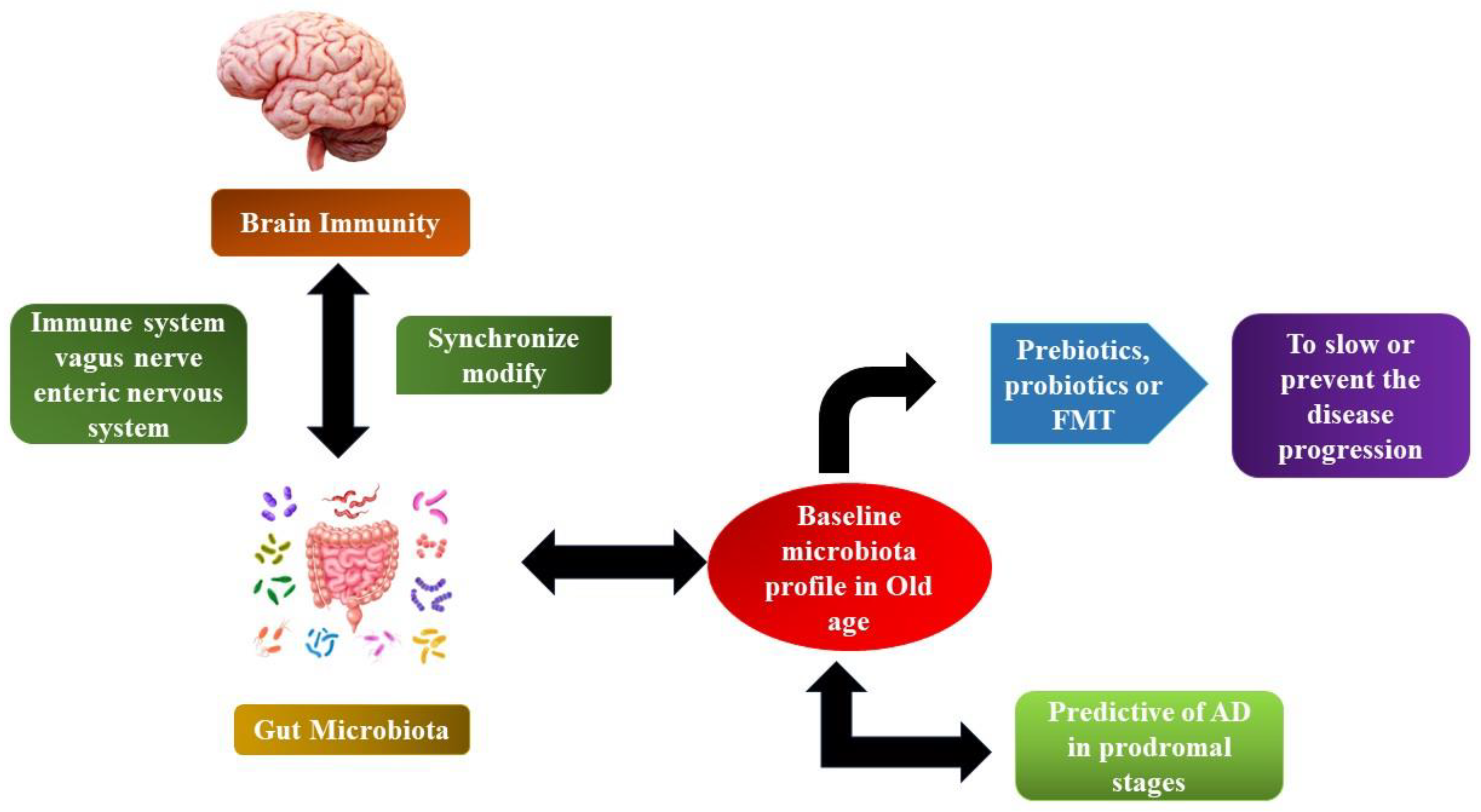
6. Aetiology of AD
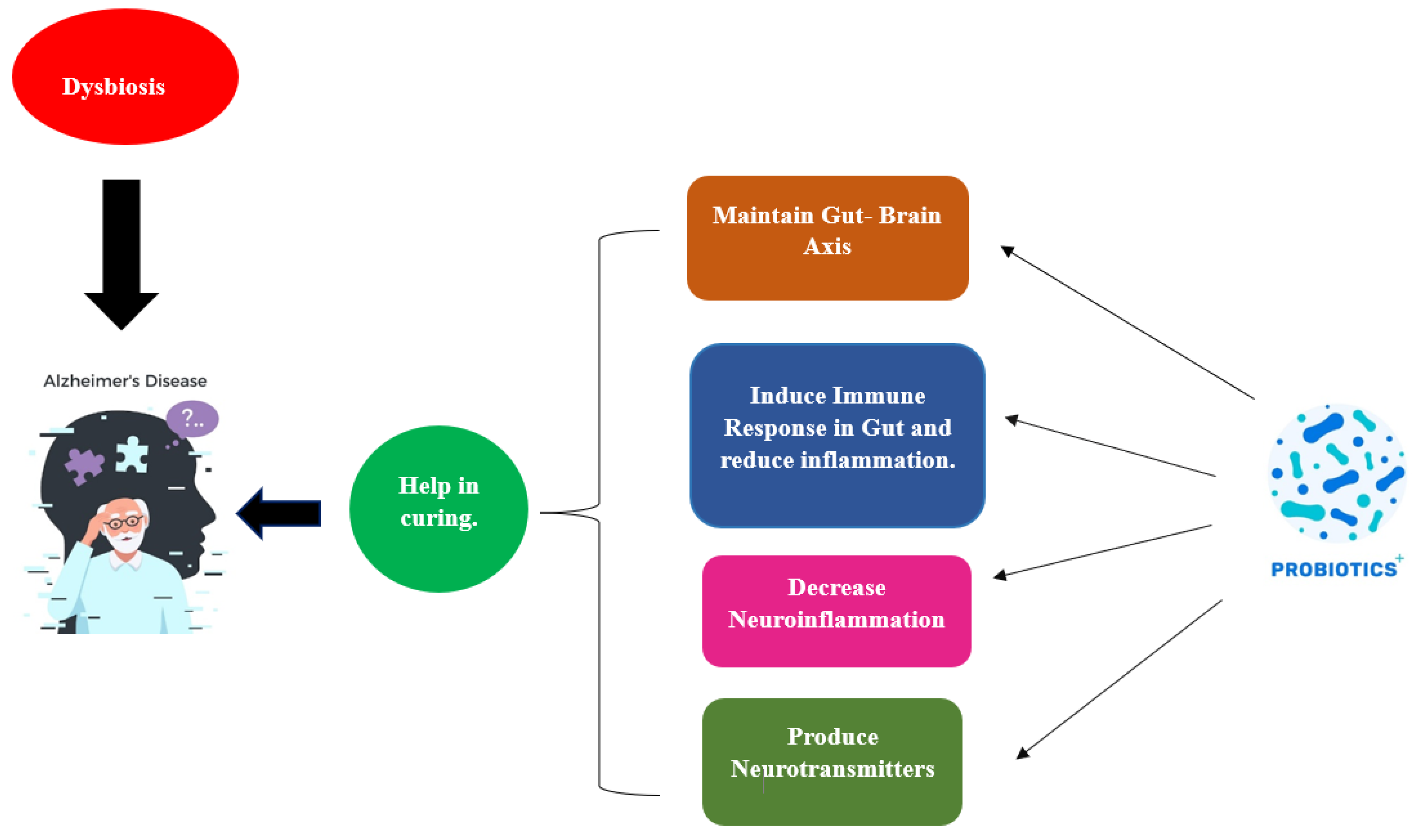
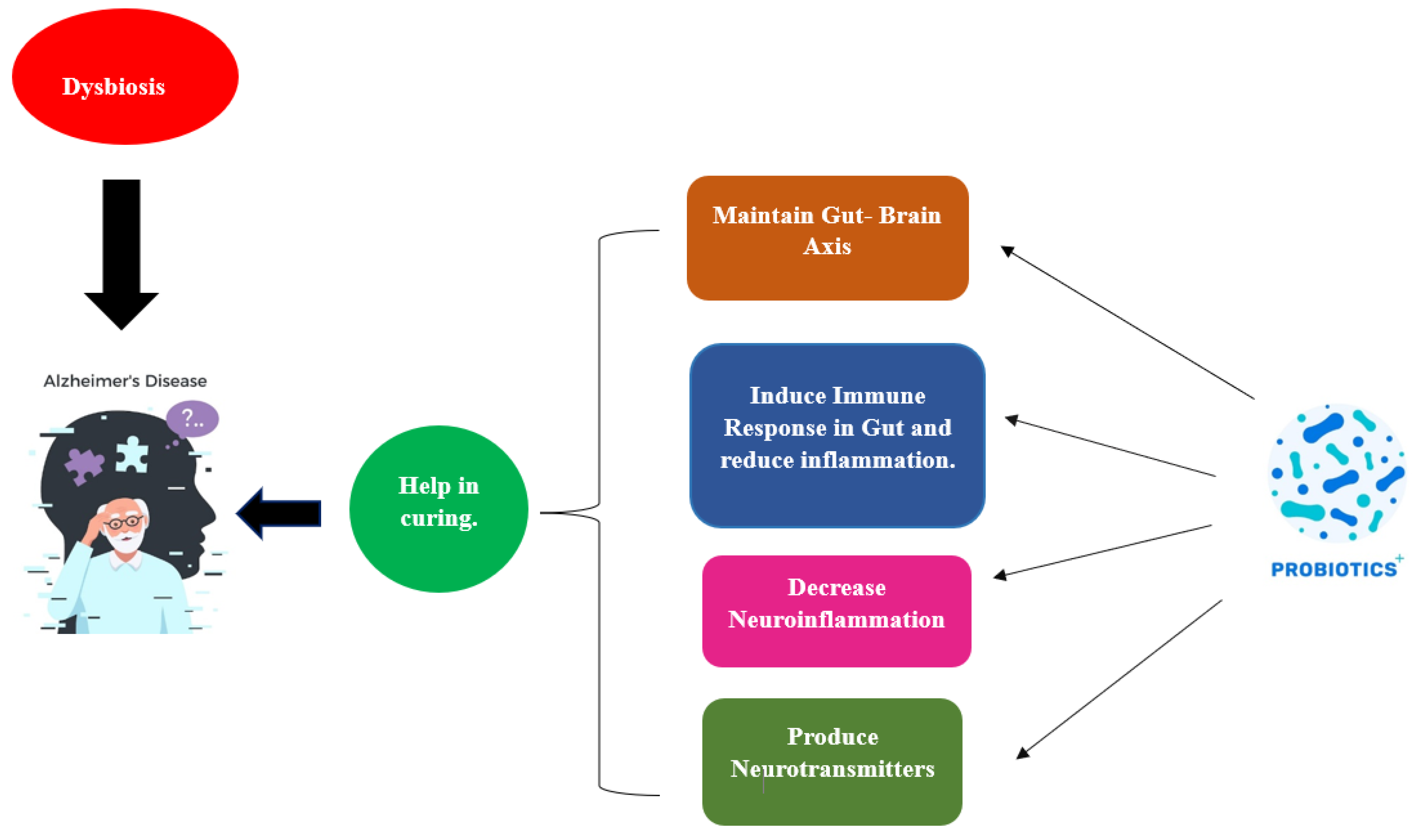
7. Significance of Probiotics in Curing AD
8. Probiotics Safety Considerations
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Monteiro, S.S.; Almeida, R.L.; Santos, N.C.; Pereira, E.M.; Silva, A.P.; Oliveira, H.M.L.; Pasquali, M.A.B. New Functional Foods with Cactus Components: Sustainable Perspectives and Future Trends. Foods 2023, 12, 2494. [Google Scholar] [CrossRef] [PubMed]
- Wieërs, G.; Belkhir, L.; Enaud, R.; Leclercq, S.; Philippart de Foy, J.M.; Dequenne, I.; de Timary, P.; Cani, P.D. How Probiotics Affect the Microbiota. Front. Cell. Infect. Microbiol. 2019, 9, 454. [Google Scholar] [CrossRef]
- Jäger, R.; Mohr, A.E.; Carpenter, K.C.; Kerksick, C.M.; Purpura, M.; Moussa, A.; Townsend, J.R.; Lamprecht, M.; West, N.P.; Black, K.; et al. International Society of Sports Nutrition Position Stand: Probiotics. J. Int. Soc. Sports Nutr. 2019, 16, 62. [Google Scholar] [CrossRef]
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Vergès, M.-C.C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Corral, G.H.; et al. Microbiome Definition Re-visited: Old Concepts and New Challenges. Microbiome 2020, 8, 103. [Google Scholar] [CrossRef] [PubMed]
- Widyastuti, Y.; Febrisiantosa, A.; Tidona, F. Health-Promoting Properties of Lactobacilli in Fermented Dairy Products. Front. Microbiol. 2021, 12, 673890. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Santamarina, A.; Gonzalez, E.G.; Lamas, A.; Mondragon, A.D.C.; Regal, P.; Miranda, J.M. Probiotics as a Possible Strategy for the Prevention and Treatment of Allergies. A Narrative Review. Foods 2021, 10, 701. [Google Scholar] [CrossRef]
- Shaikh, S.D.; Sun, N.; Canakis, A.; Park, W.Y.; Weber, H.C. Irritable Bowel Syndrome and the Gut Microbiome: A Comprehensive Review. J. Clin. Med. 2023, 12, 2558. [Google Scholar] [CrossRef]
- Lenoir-Wijnkoop, I.; Merenstein, D.; Korchagina, D.; Broholm, C.; Sanders, M.E.; Tancredi, D. Probiotics Reduce Health Care Cost and Societal Impact of Flu-Like Respiratory Tract Infections in the USA: An Economic Modeling Study. Front. Pharmacol. 2019, 10, 980. [Google Scholar] [CrossRef]
- Barta, D.G.; Cornea-Cipcigan, M.; Margaoan, R.; Vodnar, D.C. Biotechnological Processes Simulating the Natural Fermentation Process of Bee Bread and Therapeutic Properties—An Overview. Front. Nutr. 2022, 9, 871896. [Google Scholar] [CrossRef]
- Madabushi, J.S.; Khurana, P.; Gupta, N.; Gupta, M. Gut Biome and Mental Health: Do Probiotics Work? Cureus 2023, 15, e40293. [Google Scholar] [CrossRef]
- Chudzik, A.; Orzyłowska, A.; Rola, R.; Stanisz, G.J. Probiotics, Prebiotics and Postbiotics on Mitigation of Depression Symptoms: Modulation of the Brain–Gut–Microbiome Axis. Biomolecules 2021, 11, 1000. [Google Scholar] [CrossRef] [PubMed]
- Reis, D.J.; Ilardi, S.S.; Punt, S.E.W. The Anxiolytic Effect of Probiotics: A Systematic Review and Meta-analysis of the Clinical and Preclinical Literature. PLoS ONE 2018, 13, e0199041. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Lee, I.S.; Braun, C.; Enck, P. Effect of Probiotics on Central Nervous System Functions in Animals and Humans: A Systematic Review. J. Neurogastroenterol. Motil. 2016, 22, 589–605. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A.; Knight, R.; Mazmanian, S.K.; Cryan, J.F.; Tillisch, K. Gut Microbes and the Brain: Paradigm Shift in Neuroscience. J. Neurosci. 2014, 34, 15490–15496. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Li, X.; Zhang, G.; Sadiq, F.A.; Simal-Gandara, J.; Xiao, J.; Sang, Y. Probiotics in the Dairy Industry—Advances and Opportunities. Compr. Rev. Food Sci. Food Saf. 2021, 20, 3937–3982. [Google Scholar] [CrossRef]
- Arora, K.; Green, M.; Prakash, S. The Microbiome and Alzheimer’s Disease: Potential and Limitations of Prebiotic, Synbiotic, and Probiotic Formulations. Front. Bioeng. Biotechnol. 2020, 8, 537847. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, S. Yogurt Production. In Methods in Molecular Biology; Humana Press: New York, NY, USA, 2019; pp. 45–54. [Google Scholar] [CrossRef]
- Chen, W.; Narbad, A. Lactic Acid Bacteria in Foodborne Hazards Reduction; Springer: Singapore, 2018. [Google Scholar]
- Du, Y.; Xu, W.; Wu, T.; Li, H.; Hu, X.; Chen, J. Enhancement of Growth, Survival, Immunity and Disease Resistance in Litopenaeus Vannamei, by the Probiotic, Lactobacillus plantarum Ep-M17. Fish Shellfish Immunol. 2022, 129, 36–51. [Google Scholar] [CrossRef]
- Basso, M.; Johnstone, N.; Knytl, P.; Nauta, A.; Groeneveld, A.; Cohen Kadosh, K. A Systematic Review of Psychobiotic Interventions in Children and Adolescents to Enhance Cognitive Functioning and Emotional Behavior. Nutrients 2022, 14, 614. [Google Scholar] [CrossRef]
- Vasiliu, O. The Current State of Research for Psychobiotics Use in the Management of Psychiatric Disorders–A Systematic Literature Review. Front. Psychiatry 2023, 14, 1074736. [Google Scholar] [CrossRef]
- Zhu, R.; Fang, Y.; Li, H.; Liu, Y.; Wei, J.; Zhang, S.; Wang, L.; Fan, R.; Wang, L.; Li, S.; et al. Psychobiotic Lactobacillus plantarum JYLP-326 Relieves Anxiety, Depression, and Insomnia Symptoms in Test Anxious College via Modulating the Gut Microbiota and Its Metabolism. Front. Immunol. 2023, 14, 1158137. [Google Scholar] [CrossRef]
- Roy, S.; Banerjee, S.; Bhowmick, P.; Choudhury, L. Psychobiotics: Deciphering Its Role in Neuropsychiatry. World J. Bio. Pharm. Health Sci. 2023, 13, 457–464. [Google Scholar] [CrossRef]
- Sharma, R.; Gupta, D.; Mehrotra, R.; Mago, P. Psychobiotics: The Next-Generation Probiotics for the Brain. Curr. Microbiol. 2021, 78, 449–463. [Google Scholar] [CrossRef]
- Cheng, L.H.; Liu, Y.W.; Wu, C.C.; Wang, S.; Tsai, Y.C. Psychobiotics in Mental Health, Neurodegenerative and Neurodevelopmental Disorders. J. Food Drug Anal. 2019, 27, 632–648. [Google Scholar] [CrossRef]
- Kelesidis, T.; Pothoulakis, C. Efficacy and Safety of the Probiotic Saccharomyces boulardii for the Prevention and Therapy of Gastrointestinal Disorders. Ther. Adv. Gastroenterol. 2012, 5, 111–125. [Google Scholar] [CrossRef] [PubMed]
- da Silva Vale, A.; Venturim, B.C.; da Silva Rocha, A.R.F.; Martin, J.G.P.; Maske, B.L.; Balla, G.; De Dea Lindner, J.; Soccol, C.R.; de Melo Pereira, G.V. Exploring Microbial Diversity of Non-Dairy Fermented Beverages with a Focus on Functional Probiotic Microorganisms. Fermentation 2023, 9, 496. [Google Scholar] [CrossRef]
- Stavropoulou, E.; Bezirtzoglou, E. Probiotics in Medicine: A Long Debate. Front. Immunol. 2020, 11, 2192. [Google Scholar] [CrossRef]
- Farahmand, N.; Ouoba, L.I.I.; Naghizadeh Raeisi, S.; Sutherland, J.; Ghoddusi, H.B. Probiotic Lactobacilli in Fermented Dairy Products: Selective Detection, Enumeration and Identification Scheme. Microorganisms 2021, 9, 1600. [Google Scholar] [CrossRef]
- Owusu-Kwarteng, J.; Akabanda, F.; Agyei, D.; Jespersen, L. Microbial Safety of Milk Production and Fermented Dairy Products in Africa. Microorganisms 2020, 8, 752. [Google Scholar] [CrossRef]
- Dahiya, D.; Nigam, P.S. Nutrition and Health Through the Use of Probiotic Strains in Fermentation to Produce Non-dairy Functional Beverage Products Supporting Gut Microbiota. Foods 2022, 11, 2760. [Google Scholar] [CrossRef]
- Küçükgöz, K.; Trząskowska, M. Nondairy Probiotic Products: Functional Foods That Require More Attention. Nutrients 2022, 14, 753. [Google Scholar] [CrossRef]
- Aprea, G.; Del Matto, I.; Tucci, P.; Marino, L.; Scattolini, S.; Rossi, F. In Vivo Functional Properties of Dairy Bacteria. Microorganisms 2023, 11, 1787. [Google Scholar] [CrossRef] [PubMed]
- Karami, S.; Roayaei, M.; Hamzavi, H.; Bahmani, M.; Hassanzad-Azar, H.; Leila, M.; Rafieian-Kopaei, M. Isolation and Identification of Probiotic Lactobacillus from Local Dairy and Evaluating Their Antagonistic Effect on Pathogens. Int. J. Pharm. Investig. 2017, 7, 137–141. [Google Scholar] [CrossRef]
- Aspri, M.; Papademas, P.; Tsaltas, D. Review on Non-dairy Probiotics and Their Use in Non-dairy Based Products. Fermentation 2020, 6, 30. [Google Scholar] [CrossRef]
- Rasika, D.M.D.; Vidanarachchi, J.K.; Luiz, S.F.; Azeredo, D.R.P.; Cruz, A.G.; Ranadheera, C.S. Probiotic Delivery Through Non-dairy Plant-Based Food Matrices. Agriculture 2021, 11, 599. [Google Scholar] [CrossRef]
- Ballini, A.; Charitos, I.A.; Cantore, S.; Topi, S.; Bottalico, L.; Santacroce, L. About Functional Foods: The Probiotics and Prebiotics State of Art. Antibiotics 2023, 12, 635. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey, M.P.; Jones Taggart, H.J.; Strap, J.L.; Edun, G.; Green-Johnson, J.M. Milk Fermented with Lactobacillus rhamnosus R0011 Induces a Regulatory Cytokine Profile in LPS-Challenged U937 and THP-1 Macrophages. Curr. Res. Food Sci. 2020, 3, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Chen, H.; Qiao, Y.; Liu, G.; Leng, C.; Zhang, Y.; Lv, X.; Feng, Z. The Nutrient Requirements of Lactobacillus rhamnosus GG and Their Application to Fermented Milk. J. Dairy Sci. 2019, 102, 5971–5978. [Google Scholar] [CrossRef]
- Syiemlieh, I.; Morya, S. Dairy and Non-dairy Based Probiotics: A Review. J. Pharm. Innov. J. 2022, 11, 2956–2964. [Google Scholar] [CrossRef]
- Klindt-Toldam, S.; Larsen, S.K.; Saaby, L.; Olsen, L.R.; Svenstrup, G.; Müllertz, A.; Knøchel, S.; Heimdal, H.; Nielsen, D.S.; Zielińska, D. Survival of Lactobacillus acidophilus NCFM® and Bifidobacterium lactis HN019 Encapsulated in Chocolate During In Vitro Simulated Passage of the Upper Gastrointestinal Tract. LWT 2016, 74, 404–410. [Google Scholar] [CrossRef]
- Ambrogi, V.; Bottacini, F.; Cao, L.; Kuipers, B.; Schoterman, M.; van Sinderen, D. Galacto-Oligosaccharides as Infant Prebiotics: Production, Application, Bioactive Activities and Future Perspectives, Application. Crit. Rev. Food Sci. Nutr. 2023, 63, 753–766. [Google Scholar] [CrossRef]
- Alcántara, C.; Perez, M.; Huedo, P.; Altadill, T.; Espadaler-Mazo, J.; Arqués, J.L.; Zúñiga, M.; Monedero, V. Study of the Biosynthesis and Functionality of Polyphosphate in Bifidobacterium longum KABP042. Sci. Rep. 2023, 13, 11076. [Google Scholar] [CrossRef] [PubMed]
- Ansari, F.; Alian Samakkhah, S.; Bahadori, A.; Jafari, S.M.; Ziaee, M.; Khodayari, M.T.; Pourjafar, H. Health-Promoting Properties of Saccharomyces cerevisiae var. boulardii as a Probiotic; Characteristics, Isolation, and Applications in Dairy Products. Crit. Rev. Food Sci. Nutr. 2023, 63, 457–485. [Google Scholar] [CrossRef]
- dos Santos, D.C.; da Oliveira Filho, J.G.; Andretta, J.R.; Silva, F.G.; Egea, M.B. Challenges in Maintaining the Probiotic Potential in Alcoholic Beverage Development. Food Biosci. 2023, 52, 102485. [Google Scholar] [CrossRef]
- A, A.J.; Suresh, A. Oral Microbial Shift Induced by Probiotic Bacillus Coagualans along with Its Clinical Perspectives. J. Oral Biol. Craniofac. Res. 2023, 13, 398–402. [Google Scholar] [CrossRef]
- Zhao, N.; Yu, T.; Yan, F. Probiotic Role and Application of Thermophilic Bacillus as Novel Food Materials. Trends Food Sci. Technol. 2023, 138, 1–15. [Google Scholar] [CrossRef]
- Mileriene, J.; Aksomaitiene, J.; Kondrotiene, K.; Asledottir, T.; Vegarud, G.E.; Serniene, L.; Malakauskas, M. Whole-Genome Sequence of Lactococcus lactis Subsp. lactis LL16 Confirms Safety, Probiotic Potential, and Reveals Functional Traits. Microorganisms 2023, 11, 1034. [Google Scholar] [CrossRef]
- Saeed, A.; Yasmin, A.; Baig, M.; Khan, K.; Heyat, M.B.B.; Akhtar, F.; Batool, Z.; Kazmi, A.; Wahab, A.; Shahid, M.; et al. Isolation and Characterization of Lactobacillus crispatus, Lactococcus lactis, and Carnobacterium divergens as Potential Probiotic Bacteria from Fermented Black and Green Olives (Olea europaea): An Exploratory Study. BioMed Res. Int. 2023, 2023, 8726320. [Google Scholar] [CrossRef]
- Martinović, A.; Chittaro, M.; Mora, D.; Arioli, S. The Ability of Streptococcus thermophilus BT01 to Modulate Urease Activity in Healthy Subjects’ Fecal Samples Depends on the Biomass Production Process. Mol. Nutr. Food Res. 2023, 67, e2200529. [Google Scholar] [CrossRef]
- Lavelle, K.; McDonnell, B.; Fitzgerald, G.; van Sinderen, D.; Mahony, J. Bacteriophage-Host Interactions in Streptococcus thermophilus and Their Impact on Co-evolutionary Processes. FEMS Microbiol. Rev. 2023, 47, fuad032. [Google Scholar] [CrossRef]
- Çam, G.; Akın, N.; Konak Göktepe, Ç.; Demirci, T. Pea (Pisum sativum L.) Pod Powder as a Potential Enhancer of Probiotic Enterococcus faecium M74 in Ice Cream and Its Physicochemical, Structural, and Sensory Effects. J. Sci. Food Agric. 2023, 103, 3184–3193. [Google Scholar] [CrossRef]
- Aljohani, A.B.; Al-Hejin, A.M.; Shori, A.B. Bacteriocins as Promising Antimicrobial Peptides, Definition, Classification, and Their Potential Applications in Cheeses. Food Sci. Technol. 2023, 43, e118021. [Google Scholar] [CrossRef]
- Ratajczak, A.E.; Zawada, A.; Rychter, A.M.; Dobrowolska, A.; Krela-Kaźmierczak, I. Milk and Dairy Products: Good or Bad for Human Bone? Practical Dietary Recommendations for the Prevention and Management of Osteoporosis. Nutrients 2021, 13, 1329. [Google Scholar] [CrossRef]
- Vanga, S.K.; Raghavan, V. How Well Do Plant Based Alternatives Fare Nutritionally Compared to Cow’s Milk? J. Food Sci. Technol. 2018, 55, 10–20. [Google Scholar] [CrossRef]
- Hever, J.; Cronise, R.J. Plant-Based Nutrition for Healthcare Professionals: Implementing Diet as a Primary Modality in the Prevention and Treatment of Chronic Disease. J. Geriatr. Cardiol. JGC 2017, 14, 355–368. [Google Scholar] [CrossRef]
- Şanlıbaba, P. Fermented Nondairy Functional Foods Based on Probiotics. Ital. J. Food Sci. 2023, 35, 91–105. [Google Scholar] [CrossRef]
- Rizzoli, R.; Biver, E. Are Probiotics the New Calcium and Vitamin D for Bone Health? Curr. Osteoporos. Rep. 2020, 18, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Chelladhurai, K.; Ayyash, M.; Turner, M.S.; Kamal-Eldin, A. Lactobacillus helveticus: Health Effects, Current Applications, and Future Trends in Dairy Fermentation. Trends Food Sci. Technol. 2023, 136, 159–168. [Google Scholar] [CrossRef]
- Kulkarni, R.R.; Gaghan, C.; Gorrell, K.; Sharif, S.; Taha-Abdelaziz, K. Probiotics as Alternatives to Antibiotics for the Prevention and Control of Necrotic Enteritis in Chickens. Pathogens 2022, 11, 692. [Google Scholar] [CrossRef]
- Bernardeau, M.; Vernoux, J.P. Overview of Differences Between Microbial Feed Additives and Probiotics for Food Regarding Regulation, Growth Promotion Effects and Health Properties and Consequences for Extrapolation of Farm Animal Results to Humans. Clin. Microbiol. Infect. 2013, 19, 321–330. [Google Scholar] [CrossRef]
- Cai, J.; Rimal, B.; Jiang, C.; Chiang, J.Y.L.; Patterson, A.D. Bile Acid Metabolism and Signaling, the Microbiota, and Metabolic Disease. Pharmacol. Ther. 2022, 237, 108238. [Google Scholar] [CrossRef]
- Collins, S.L.; Stine, J.G.; Bisanz, J.E.; Okafor, C.D.; Patterson, A.D. Bile Acids and the Gut Microbiota: Metabolic Interactions and Impacts on Disease. Nat. Rev. Microbiol. 2023, 21, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Kavitake, D.; Tiwari, S.; Shah, I.A.; Devi, P.B.; Delattre, C.; Reddy, G.B.; Shetty, P.H. Antipathogenic Potentials of Exopolysaccharides Produced by Lactic Acid Bacteria and Their Food and Health Applications. Food Control 2023, 152, 109850. [Google Scholar] [CrossRef]
- Langa, S.; Peirotén, Á.; Curiel, J.A.; de la Bastida, A.R.; Landete, J.M. Isoflavone Metabolism by Lactic Acid Bacteria and Its Application in the Development of Fermented Soy Food with Beneficial Effects on Human Health. Foods 2023, 12, 1293. [Google Scholar] [CrossRef]
- Vlasova, A.N.; Kandasamy, S.; Chattha, K.S.; Rajashekara, G.; Saif, L.J. Comparison of Probiotic Lactobacilli and Bifidobacteria Effects, Immune Responses and Rotavirus Vaccines and Infection in Different Host Species. Vet. Immunol. Immunopathol. 2016, 172, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Pahumunto, N.; Dahlen, G.; Teanpaisan, R. Evaluation of Potential Probiotic Properties of Lactobacillus and Bacillus Strains Derived from Various Sources for Their Potential Use in Swine Feeding. Probiotics Antimicrob. Proteins 2023, 15, 479–490. [Google Scholar] [CrossRef]
- Coimbra-Gomes, J.; Reis, P.J.M.; Tavares, T.G.; Faria, M.A.; Malcata, F.X.; Macedo, A.C. Evaluating the Probiotic Potential of Lactic Acid Bacteria Implicated in Natural Fermentation of Table Olives, cv. Cobrançosa. Molecules 2023, 28, 3285. [Google Scholar] [CrossRef]
- Foongsawat, N.; Sunthornthummas, S.; Nantavisai, K.; Surachat, K.; Rangsiruji, A.; Sarawaneeyaruk, S.; Insian, K.; Sukontasing, S.; Suwannasai, N.; Pringsulaka, O. Isolation, Characterization, and Comparative Genomics of the Novel Potential Probiotics from Canine Feces. Food Sci. Anim. Resour. 2023, 43, 685–702. [Google Scholar] [CrossRef]
- Mirzabekyan, S.; Harutyunyan, N.; Manvelyan, A.; Malkhasyan, L.; Balayan, M.; Miralimova, S.; Chikindas, M.L.; Chistyakov, V.; Pepoyan, A. Fish Probiotics: Cell Surface Properties of Fish Intestinal Lactobacilli and Escherichia coli. Microorganisms 2023, 11, 595. [Google Scholar] [CrossRef]
- Haryani, Y.; Halid, N.A.; Guat, G.S.; Nor-Khaizura, M.A.R.; Hatta, M.A.M.; Sabri, S.; Radu, S.; Hasan, H. High Prevalence of Multiple Antibiotic Resistance in Fermented Food-Associated Lactic Acid Bacteria in Malaysia. Food Control 2023, 147, 109558. [Google Scholar] [CrossRef]
- Asan-Ozusaglam, M.; Gunyakti, A. Lactobacillus fermentum Strains from Human Breast Milk with Probiotic Properties and Cholesterol-Lowering Effects. Food Sci. Biotechnol. 2019, 28, 501–509. [Google Scholar] [CrossRef]
- Keresztény, T.; Libisch, B.; Orbe, S.C.; Nagy, T.; Kerényi, Z.; Kocsis, R.; Posta, K.; Papp, P.P.; Olasz, F. Isolation and Characterization of Lactic Acid Bacteria with Probiotic Attributes from Different Parts of the Gastrointestinal Tract of Free-Living Wild Boars in Hungary. Probiotics Antimicrob. Proteins 2023, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Moiseenko, K.V.; Begunova, A.V.; Savinova, O.S.; Glazunova, O.A.; Rozhkova, I.V.; Fedorova, T.V. Biochemical and Genomic Characterization of Two New Strains of Lacticaseibacillus paracasei Isolated from the Traditional Corn-Based Beverage of South Africa, Mahewu, and Their Comparison with Strains Isolated from Kefir Grains. Foods 2023, 12, 223. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.H.; Yang, R.S.; Lin, Y.C.; Xin, W.G.; Zhou, H.Y.; Wang, F.; Zhang, Q.L.; Lin, L.B. Assessment of the Safety and Probiotic Characteristics of Lactobacillus salivarius CGMCC20700 Based on Whole-Genome Sequencing and Phenotypic Analysis. Front. Microbiol. 2023, 14, 1120263. [Google Scholar] [CrossRef] [PubMed]
- Yalçin, S.K.; Ozbas, Z.Y. Yeasts from Traditional Cheeses for Potential Applications. Glob. Food Secur. Wellness 2017, 3, 277–293. [Google Scholar]
- Mazziotta, C.; Tognon, M.; Martini, F.; Torreggiani, E.; Rotondo, J.C. Probiotics Mechanism of Action on Immune Cells and Beneficial Effects on Human Health. Cells 2023, 12, 184. [Google Scholar] [CrossRef] [PubMed]
- National Institutes of Health. Probiotics: Fact Sheet for Health Professionals; National Institutes of Health: Bethesda, MD, USA, 2020.
- Dempsey, E.; Corr, S.C. Lactobacillus spp. for Gastrointestinal Health: Current and Future Perspectives. Front. Immunol. 2022, 13, 840245. [Google Scholar] [CrossRef]
- Sharma, R.; Diwan, B.; Singh, B.P.; Kulshrestha, S. Probiotic Fermentation of Polyphenols: Potential Sources of Novel Functional Foods. Food Prod. Process. Nutr. 2022, 4, 21. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Rodrigues, C.F.; Stojanović-Radić, Z.; Dimitrijević, M.; Aleksić, A.; Neffe-Skocińska, K.; Zielińska, D.; Kołożyn-Krajewska, D.; Salehi, B.; Milton Prabu, S.; et al. Probiotics: Versatile Bioactive Components in Promoting Human Health. Medicina 2020, 56, 433. [Google Scholar] [CrossRef]
- Mohammadi, M.; Shadnoush, M.; Sohrabvandi, S.; Yousefi, M.; Khorshidian, N.; Mortazavian, A.M. Probiotics as Potential Detoxification Tools for Mitigation of Pesticides: A Mini Review. Int. J. Food Sci. Technol. 2021, 56, 2078–2087. [Google Scholar] [CrossRef]
- Maske, B.L.; de Melo Pereira, G.V.; Vale, A.D.S.; De Carvalho Neto, D.P.; Karp, S.G.; Viesser, J.A.; De Dea Lindner, J.; Pagnoncelli, M.G.; Soccol, V.T.; Soccol, C.R. A Review on Enzyme-Producing Lactobacilli Associated with the Human Digestive Process: From Metabolism to Application. Enzyme Microb. Technol. 2021, 149, 109836. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, P.; Zhang, X. Probiotics Regulate Gut Microbiota: An Effective Method to Improve Immunity. Molecules 2021, 26, 6076. [Google Scholar] [CrossRef]
- Xie, N.; Zhang, L.; Gao, W.; Huang, C.; Huber, P.E.; Zhou, X.; Li, C.; Shen, G.; Zou, B. NAD+ Metabolism: Pathophysiologic Mechanisms and Therapeutic Potential. Signal Transduct. Target. Ther. 2020, 5, 227. [Google Scholar] [CrossRef]
- Grujović, M.Ž.; Mladenović, K.G.; Semedo-Lemsaddek, T.; Laranjo, M.; Stefanović, O.D.; Kocić-Tanackov, S.D. Advantages and Disadvantages of Non-starter Lactic Acid Bacteria from Traditional Fermented Foods: Potential Use as Starters or Probiotics. Compr. Rev. Food Sci. Food Saf. 2022, 21, 1537–1567. [Google Scholar] [CrossRef] [PubMed]
- Rolfe, C.; Daryaei, H. Intrinsic and Extrinsic Factors Affecting Microbial Growth in Food Systems. In Food Safety Engineering; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Śliżewska, K.; Chlebicz-Wójcik, A. Growth Kinetics of Probiotic Lactobacillus Strains in the Alternative, Cost-Efficient Semi-solid Fermentation Medium. Biology 2020, 9, 423. [Google Scholar] [CrossRef] [PubMed]
- Abedi, E.; Hashemi, S.M.B. Lactic Acid Production–Producing Microorganisms and Substrates Sources-State of Art. Heliyon 2020, 6, e04974. [Google Scholar] [CrossRef] [PubMed]
- Nkhata, S.G.; Ayua, E.; Kamau, E.H.; Shingiro, J.B. Fermentation and Germination Improve Nutritional Value of Cereals and Legumes Through Activation of Endogenous Enzymes. Food Sci. Nutr. 2018, 6, 2446–2458. [Google Scholar] [CrossRef]
- Abdul Hakim, B.N.; Xuan, N.J.; Oslan, S.N.H. A Comprehensive Review of Bioactive Compounds from Lactic Acid Bacteria: Potential Functions as Functional Food in Dietetics and the Food Industry. Foods 2023, 12, 2850. [Google Scholar] [CrossRef]
- Szutowska, J. Functional Properties of Lactic Acid Bacteria in Fermented Fruit and Vegetable Juices: A Systematic Literature Review. Eur. Food Res. Technol. 2020, 246, 357–372. [Google Scholar] [CrossRef]
- Plaza-Vinuesa, L.; Sánchez-Arroyo, A.; López De Felipe, F.; de Las Rivas, B.; Muñoz, R. Non-redundant Functionality of Lactiplantibacillus plantarum Phospho-β-Glucosidases Revealed by Carbohydrate Utilization Signatures Associated to pbg2 and pbg4 Gene Mutants. J. Appl. Microbiol. 2023, 134, lxad077. [Google Scholar] [CrossRef]
- Li, J.; Jia, S.; Ma, D.; Deng, X.; Tian, J.; Wang, R.; Li, J.; Shan, A. Effects of Citric Acid and Heterofermentative Inoculants on Anaerobic Co-fermentation of Chinese Cabbage Waste and Wheat Bran. Bioresour. Technol. 2023, 377, 128942. [Google Scholar] [CrossRef]
- Lacroux, J.; Llamas, M.; Dauptain, K.; Avila, R.; Steyer, J.P.; van Lis, R.; Trably, E. Dark Fermentation and Microalgae Cultivation Coupled Systems: Outlook and Challenges. Sci. Total Environ. 2023, 865, 161136. [Google Scholar] [CrossRef] [PubMed]
- Jabłońska-Ryś, E.; Sławińska, A.; Skrzypczak, K.; Goral, K. Dynamics of Changes in pH and the Contents of Free Sugars, Organic Acids and LAB in Button Mushrooms During Controlled Lactic Fermentation. Foods 2022, 11, 1553. [Google Scholar] [CrossRef] [PubMed]
- Upadhyaya, B.P.; DeVeaux, L.C.; Christopher, L.P. Metabolic Engineering as a Tool for Enhanced Lactic Acid Production. Trends Biotechnol. 2014, 32, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Sivamaruthi, B.S.; Prasanth, M.I.; Kesika, P.; Chaiyasut, C. Probiotics in Human Mental Health and Diseases-A Minireview. Trop. J. Pharm. Res. 2019, 18, 889–895. [Google Scholar] [CrossRef]
- Eastwood, J.; Walton, G.; Van Hemert, S.; Williams, C.; Lamport, D. The Effect of Probiotics on Cognitive Function Across the Human Lifespan: A Systematic Review. Neurosci. Biobehav. Rev. 2021, 128, 311–327. [Google Scholar] [CrossRef]
- Socała, K.; Doboszewska, U.; Szopa, A.; Serefko, A.; Włodarczyk, M.; Zielińska, A.; Poleszak, E.; Fichna, J.; Wlaź, P. The Role of Microbiota-Gut-Brain Axis in Neuropsychiatric and Neurological Disorders. Pharmacol. Res. 2021, 172, 105840. [Google Scholar] [CrossRef]
- Papalini, S.; Michels, F.; Kohn, N.; Wegman, J.; van Hemert, S.; Roelofs, K.; Arias-Vasquez, A.; Aarts, E. Stress Matters: Randomized Controlled Trial on the Effect of Probiotics on Neurocognition. Neurobiol. Stress 2019, 10, 100141. [Google Scholar] [CrossRef]
- Roussin, L.; Prince, N.; Perez-Pardo, P.; Kraneveld, A.D.; Rabot, S.; Naudon, L. Role of the Gut Microbiota in the Pathophysiology of Autism Spectrum Disorder: Clinical and Preclinical Evidence. Microorganisms 2020, 8, 1369. [Google Scholar] [CrossRef]
- Li, Q.; Han, Y.; Dy, A.B.C.; Hagerman, R.J. The Gut Microbiota and Autism Spectrum Disorders. Front. Cell. Neurosci. 2017, 11, 120. [Google Scholar] [CrossRef]
- Vellingiri, B.; Aishwarya, S.Y.; Benita Jancy, S.; Sriram Abhishek, G.; Winster Suresh Babu, H.; Vijayakumar, P.; Narayanasamy, A.; Mariappan, S.; Sangeetha, R.; Valsala Gopalakrishnan, A.; et al. An Anxious Relationship Between Autism Spectrum Disorder and Gut Microbiota: A Tangled Chemistry? J. Clin. Neurosci. 2022, 99, 169–189. [Google Scholar] [CrossRef]
- Savignac, H.M.; Kiely, B.; Dinan, T.G.; Cryan, J.F. Bifidobacteriaexert Exert Strain-Specific Effects on Stress-Related Behavior and Physiology in BALB/C Mice. Neurogastroenterol. Motil. 2014, 26, 1615–1627. [Google Scholar] [CrossRef]
- Srivastav, S.; Neupane, S.; Bhurtel, S.; Katila, N.; Maharjan, S.; Choi, H.; Hong, J.T.; Choi, D.Y. Probiotics Mixture Increases Butyrate, and Subsequently Rescues the Nigral Dopaminergic Neurons from MPTP and Rotenone-Induced Neurotoxicity. J. Nutr. Biochem. 2019, 69, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.H.; Hor, J.W.; Chong, C.W.; Lim, S.Y. Probiotics for Parkinson’s Disease: Current Evidence and Future Directions. JGH Open 2021, 5, 414–419. [Google Scholar] [CrossRef]
- Allen, A.P.; Hutch, W.; Borre, Y.E.; Kennedy, P.J.; Temko, A.; Boylan, G.; Murphy, E.; Cryan, J.F.; Dinan, T.G.; Clarke, G. Bifidobacterium longum 1714 as a Translational Psychobiotic: Modulation of Stress, Electrophysiology and Neurocognition in Healthy Volunteers. Transl. Psychiatry 2016, 6, e939. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Braun, C.; Murphy, E.F.; Enck, P. Bifidobacterium longum 1714™ Strain Modulates Brain Activity of Healthy Volunteers During Social Stress. Am. J. Gastroenterol. 2019, 114, 1152–1162. [Google Scholar] [CrossRef]
- Kumar, A.; Sidhu, J.; Goyal, A.; Tsao, J.W.; Doerr, C. Alzheimer Disease (Nursing). In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- DeTure, M.A.; Dickson, D.W. The Neuropathological Diagnosis of Alzheimer’s Disease. Mol. Neurodegener. 2019, 14, 32. [Google Scholar] [CrossRef] [PubMed]
- Simoes, S.; Guo, J.; Buitrago, L.; Qureshi, Y.H.; Feng, X.; Kothiya, M.; Cortes, E.; Patel, V.; Kannan, S.; Kim, Y.H.; et al. Alzheimer’s Vulnerable Brain Region Relies on a Distinct Retromer Core Dedicated to Endosomal Recycling. Cell Rep. 2021, 37, 110182. [Google Scholar] [CrossRef]
- Tan, J.Z.A.; Gleeson, P.A. The Role of Membrane Trafficking in the Processing of Amyloid Precursor Protein and Production of Amyloid Peptides in Alzheimer’s Disease. Biochim. Biophys. Acta Biomembr. 2019, 1861, 697–712. [Google Scholar] [CrossRef]
- Evrard, C.; Gilet, A.L.; Colombel, F.; Dufermont, E.; Corson, Y. Now You Make False Memories; Now You Do Not: The Order of Presentation of Words in DRM Lists Influences the Production of the Critical Lure in Alzheimer’s Disease. Psychol. Res. 2018, 82, 429–438. [Google Scholar] [CrossRef]
- Naomi, R.; Embong, H.; Othman, F.; Ghazi, H.F.; Maruthey, N.; Bahari, H. Probiotics for Alzheimer’s Disease: A Systematic Review. Nutrients 2021, 14, 20. [Google Scholar] [CrossRef]
- Steiner, H.; Fukumori, A.; Tagami, S.; Okochi, M. Making the Final Cut: Pathogenic Amyloid-β Peptide Generation by γ-Secretase. Cell Stress 2018, 2, 292–310. [Google Scholar] [CrossRef]
- Kinney, J.W.; Bemiller, S.M.; Murtishaw, A.S.; Leisgang, A.M.; Salazar, A.M.; Lamb, B.T. Inflammation as a Central Mechanism in Alzheimer’s Disease. Alzheimers. Dement. 2018, 4, 575–590. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Sisodia, S.S.; Vassar, R.J. The Gut Microbiome in Alzheimer’s Disease: What We Know and What Remains to Be Explored. Mol. Neurodegener. 2023, 18, 9. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Yang, D.; Sun, J.; Li, Y. Probiotic Supplements Are Effective in People with Cognitive Impairment: A Meta-analysis of Randomized Controlled Trials. Nutr. Rev. 2023, 81, 1091–1104. [Google Scholar] [CrossRef]
- Drljača, J.; Milošević, N.; Milanović, M.; Abenavoli, L.; Milić, N. When the Microbiome Helps the Brain-Current Evidence. CNS Neurosci. Ther. 2023, 29 (Suppl. S1), 43–58. [Google Scholar] [CrossRef]
- Zhu, G.; Zhao, J.; Wang, G.; Chen, W. Bifidobacterium breve HNXY26M4 Attenuates Cognitive Deficits and Neuroinflammation by Regulating the gut-brain Axis in APP/PS1 Mice. J. Agric. Food Chem. 2023, 71, 4646–4655. [Google Scholar] [CrossRef]
- Song, X.; Zhao, Z.; Zhao, Y.; Wang, Z.; Wang, C.; Yang, G.; Li, S. Lactobacillus plantarum DP189 Prevents Cognitive Dysfunction in D-Galactose/AlCl3 Induced Mouse Model of Alzheimer’s Disease via Modulating Gut Microbiota and PI3K/Akt/GSK-3β Signaling Pathway. Nutr. Neurosci. 2022, 25, 2588–2600. [Google Scholar] [CrossRef]
- Ohsawa, K.; Uchida, N.; Ohki, K.; Nakamura, Y.; Yokogoshi, H. Lactobacillus helveticus–Fermented Milk Improves Learning and Memory in Mice. Nutr. Neurosci. 2015, 18, 232–240. [Google Scholar] [CrossRef]
- Bairamian, D.; Sha, S.; Rolhion, N.; Sokol, H.; Dorothée, G.; Lemere, C.A.; Krantic, S. Microbiota in Neuroinflammation and Synaptic Dysfunction: A Focus on Alzheimer’s Disease. Mol. Neurodegener. 2022, 17, 19. [Google Scholar] [CrossRef]
- Bernier, F.; Kuhara, T.; Xiao, J. Probiotic Bifidobacterium breve MCC1274 Protects Against Oxidative Stress and Neuronal Lipid Droplet Formation via PLIN4 Gene Regulation. Microorganisms 2023, 11, 791. [Google Scholar] [CrossRef]
- Verbeke, K.A.; Boobis, A.R.; Chiodini, A.; Edwards, C.A.; Franck, A.; Kleerebezem, M.; Nauta, A.; Raes, J.; van Tol, E.A.; Tuohy, K.M. Towards Microbial Fermentation Metabolites as Markers for Health Benefits of Prebiotics. Nutr. Res. Rev. 2015, 28, 42–66. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Psichas, A.; Sleeth, M.L.; Murphy, K.G.; Brooks, L.; Bewick, G.A.; Hanyaloglu, A.C.; Ghatei, M.A.; Bloom, S.R.; Frost, G. The Short Chain Fatty Acid Propionate Stimulates GLP-1 and PYY Secretion via Free Fatty Acid Receptor 2 in Rodents. In Int. J. Obes. 2015, 39, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Y.; Du, Y.F.; Chen, L. Neuropeptides Exert Neuroprotective Effects in Alzheimer’s Disease. Front. Mol. Neurosci. 2018, 11, 493. [Google Scholar] [CrossRef]
- Nimgampalle, M.; Kuna, Y. Anti-Alzheimer Properties of Probiotic, Lactobacillus plantarum MTCC 1325 in Alzheimer’s Disease Induced Albino Rats. J. Clin. Diagn. Res. 2017, 11, KC01–KC05. [Google Scholar] [CrossRef] [PubMed]
- Moya-Pérez, A.; Perez-Villalba, A.; Benítez-Páez, A.; Campillo, I.; Sanz, Y. Bifidobacterium CECT 7765 Modulates Early Stress-Induced Immune, Neuroendocrine, and Behavioral Alterations in Mice. Brain Behav. Immun. 2017, 65, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.S.; Begeny, C.T.; Boyle, P.A.; Schneider, J.A.; Bennett, D.A. Vulnerability to Stress, Anxiety, and Development of Dementia in Old Age. Am. J. Geriatr. Psychiatry 2011, 19, 327–334. [Google Scholar] [CrossRef]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous Bacteria from the Gut Microbiota Regulate Host Serotonin Biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef]
- Mindus, C.; Ellis, J.; Van Staaveren, N.; Harlander-Matauschek, A. Lactobacillus-Based Probiotics Reduce the Adverse Effects of Stress in Rodents: A Meta-analysis. Front. Behav. Neurosci. 2021, 15, 642757. [Google Scholar] [CrossRef]
- Bonfili, L.; Cecarini, V.; Gogoi, O.; Berardi, S.; Scarpona, S.; Angeletti, M.; Rossi, G.; Eleuteri, A.M. Gut Microbiota Manipulation Through Probiotics Oral Administration Restores Glucose Homeostasis in a Mouse Model of Alzheimer’s Disease. Neurobiol. Aging 2020, 87, 35–43. [Google Scholar] [CrossRef]
- Ji, H.F.; Shen, L. Probiotics as Potential Therapeutic Options for Alzheimer’s Disease. Appl. Microbiol. Biotechnol. 2021, 105, 7721–7730. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.H.; Park, S.; Paik, J.W.; Chae, S.W.; Kim, D.H.; Jeong, D.G.; Ha, E.; Kim, M.; Hong, G.; Park, S.H.; et al. Efficacy and Safety of Lactobacillus plantarum C29-Fermented Soybean (DW2009) in Individuals with Mild Cognitive Impairment: A 12-Week, Multi-center, Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Nutrients 2019, 11, 305. [Google Scholar] [CrossRef] [PubMed]
- Dudek-Wicher, R.; Junka, A.; Paleczny, J.; Bartoszewicz, M. Clinical Trials of Probiotic Strains in Selected Disease Entities. Int. J. Microbiol. 2020, 2020, 8854119. [Google Scholar] [CrossRef] [PubMed]
- Merenstein, D.; Pot, B.; Leyer, G.; Ouwehand, A.C.; Preidis, G.A.; Elkins, C.A.; Hill, C.; Lewis, Z.T.; Shane, A.L.; Zmora, N.; et al. Emerging Issues in Probiotic Safety: 2023 Perspectives. Gut Microbes 2023, 15, 2185034. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | Dairy-Based Probiotics | Non-Dairy-Based Probiotics | Mechanisms | Effects on Mental Health | References |
|---|---|---|---|---|---|
| Lactobacillaceae acidophilus | Commonly found in yogurt and fermented dairy products | Found in non-dairy sources like fermented non-dairy beverages | Regulation of neurotransmitters, modulation of the gut-brain axis, and reduction in inflammation | May reduce symptoms of anxiety and depression; improves mood and cognitive function | [28,33,34,35] |
| Lactobacillaceae casei | Frequently present in dairy-based probiotics like yogurt | Also found in non-dairy probiotics | Modulation of neurotransmitters and improvement of intestinal barrier function | Potential to alleviate symptoms of stress, anxiety, and depression | [31,36,37]. |
| Lactobacillaceae rhamnosus | Found in dairy-based probiotics such as yogurt | Also present in non-dairy probiotics | Modulation of neurotransmitters and enhancement of GABA receptor function | May reduce symptoms of anxiety and stress and improve mood | [35,38,39] |
| Bifidobacterium infantis | Often included in dairy-based probiotics like yogurt | Also used in non-dairy probiotic formulations | Regulation of neurotransmitters and reduction in inflammation | Potential to improve mood and reduce symptoms of depression and anxiety | [40,41] |
| Bifidobacterium longum | Commonly found in dairy-based probiotics like yogurt | Also used in non-dairy probiotic formulations | Regulation of neurotransmitters and modulation of the gut-brain axis | May have positive effects on mood, stress reduction, and cognitive function | [42,43] |
| Saccharomyces boulardii | Non-dairy probiotic commonly used in supplements | Frequently used in non-dairy probiotic formulations | Modulation of neurotransmitters and enhancement of intestinal barrier function | May have a positive impact on mental health and gastrointestinal disorders | [44,45] |
| Bacillus coagulans | Found in dairy-based probiotic products like yogurt | Also used in non-dairy probiotic formulations | Production of neurotransmitters and modulation of the immune system | Potential to support mental well-being and reduce anxiety | [46,47] |
| Lactococcus lactis | Frequently used in dairy-based probiotic formulations | Not commonly used in non-dairy probiotics | Production of neurotransmitters and modulation of the immune system | Limited studies on mental health effects | [48,49] |
| Streptococcus thermophilus | Commonly found in yogurt and other dairy products | Not typically used in non-dairy probiotics | Not extensively studied for mental health effects | Limited studies on mental health effects | [50,51] |
| Enterococcus faecium | Often present in dairy-based probiotics like fermented milk products | Also found in non-dairy probiotics | Not extensively studied for mental health effects | Limited studies on mental health effects | [52,53] |
| Lactobacillus-Originated Probiotics | Probiotics Originated from Other Bacterial Strains | References |
|---|---|---|
| Widely accepted and high consumer demand | Limited consumer demand and niche markets | [67,68] |
| Extensively studied and approved for use | Varies depending on the strain and region | [69,70] |
| Compatible with various processing technologies | Compatible with specific processes | [71] |
| Can have a relatively long shelf life | Varies depending on the strain and formulation | [72,73] |
| Well-documented and broad range of benefits | Specific benefits with less research support | [74,75] |
| Dairy-based products, fermented foods, and supplements | Niche products and specific dietary preferences | [76] |
| Established manufacturing processes | May require optimization for commercial production | [67] |
| Neurological Disorder | Probiotics Strain | Mechanism | Reference/Author |
|---|---|---|---|
| Autism spectrum disorder | Lactobacillaceae reuteri | It improves the neurotransmission and decreases the level of corticosterone. | [102] |
| Bifidobacterium fragilis | It restores the gut microbiota and improves gut permeability and physiology. | [103,104] | |
| Stress and depression | B. infantis | It has been seen that in the rat the treatment of B. infantis normalized the immune response and norepinephrine level in the brainstem. | [105] |
| Parkinson’s disease- | L. rhamnosus GG, L. acidophilus, and Bifidobacterium animalis subsp. lactis | It increases the level of butyrate and the levels of BDNF and GDNF, which induces the survival of the neuronal cell and the synthesis of dopamine. | [106,107] |
| Reducing stress and enhancing memory | Bifidobacterium longum (B. longum) 1714 | It induces an anti-stress response that helps in the reduction in stress. | [108,109] |
| Probiotic Strain | Effects on AD | Mechanisms | Reference |
|---|---|---|---|
| Lactobacillaceae acidophilus | Improved cognitive function, reduced neuroinflammation, and enhanced amyloid clearance | Modulation of gut-brain axis, anti-inflammatory effects, and interaction with amyloid proteins | [118,119] |
| Bifidobacterium longum | Enhanced memory, reduced amyloid plaques, and attenuated neuroinflammation | Modulation of immune response, reduction in oxidative stress, and interaction with amyloid proteins. | [118,120] |
| Lactobacillaceae plantarium | Protection against cognitive decline, reduced neuroinflammation, and improved synaptic function | Regulation of neurotransmitters and anti-inflammatory effects and maintenance of blood–brain barrier integrity | [121,122] |
| Lactobacillaceae helveticus | Ameliorated memory deficits, reduced neuroinflammation, and improved neurogenesis | Stress reduction, anti-inflammatory effects, and promotion of neuronal growth and connectivity | [120,123] |
| Bifidobacterium breve | Reduced cognitive decline, decreased amyloid accumulation, and attenuated neuroinflammation. | Regulation of gut permeability, anti-inflammatory effects, and modulation of immune response | [124,125] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tripathi, A.; Pandey, V.K.; Tiwari, V.; Mishra, R.; Dash, K.K.; Harsányi, E.; Kovács, B.; Shaikh, A.M. Exploring the Fermentation-Driven Functionalities of Lactobacillaceae-Originated Probiotics in Preventive Measures of Alzheimer’s Disease: A Review. Fermentation 2023, 9, 762. https://doi.org/10.3390/fermentation9080762
Tripathi A, Pandey VK, Tiwari V, Mishra R, Dash KK, Harsányi E, Kovács B, Shaikh AM. Exploring the Fermentation-Driven Functionalities of Lactobacillaceae-Originated Probiotics in Preventive Measures of Alzheimer’s Disease: A Review. Fermentation. 2023; 9(8):762. https://doi.org/10.3390/fermentation9080762
Chicago/Turabian StyleTripathi, Anjali, Vinay Kumar Pandey, Vivek Tiwari, Rashi Mishra, Kshirod Kumar Dash, Endre Harsányi, Béla Kovács, and Ayaz Mukarram Shaikh. 2023. "Exploring the Fermentation-Driven Functionalities of Lactobacillaceae-Originated Probiotics in Preventive Measures of Alzheimer’s Disease: A Review" Fermentation 9, no. 8: 762. https://doi.org/10.3390/fermentation9080762
APA StyleTripathi, A., Pandey, V. K., Tiwari, V., Mishra, R., Dash, K. K., Harsányi, E., Kovács, B., & Shaikh, A. M. (2023). Exploring the Fermentation-Driven Functionalities of Lactobacillaceae-Originated Probiotics in Preventive Measures of Alzheimer’s Disease: A Review. Fermentation, 9(8), 762. https://doi.org/10.3390/fermentation9080762