
Metabolic Engineering of Microorganisms to Produce L-Aspartate and Its Derivatives



Abstract
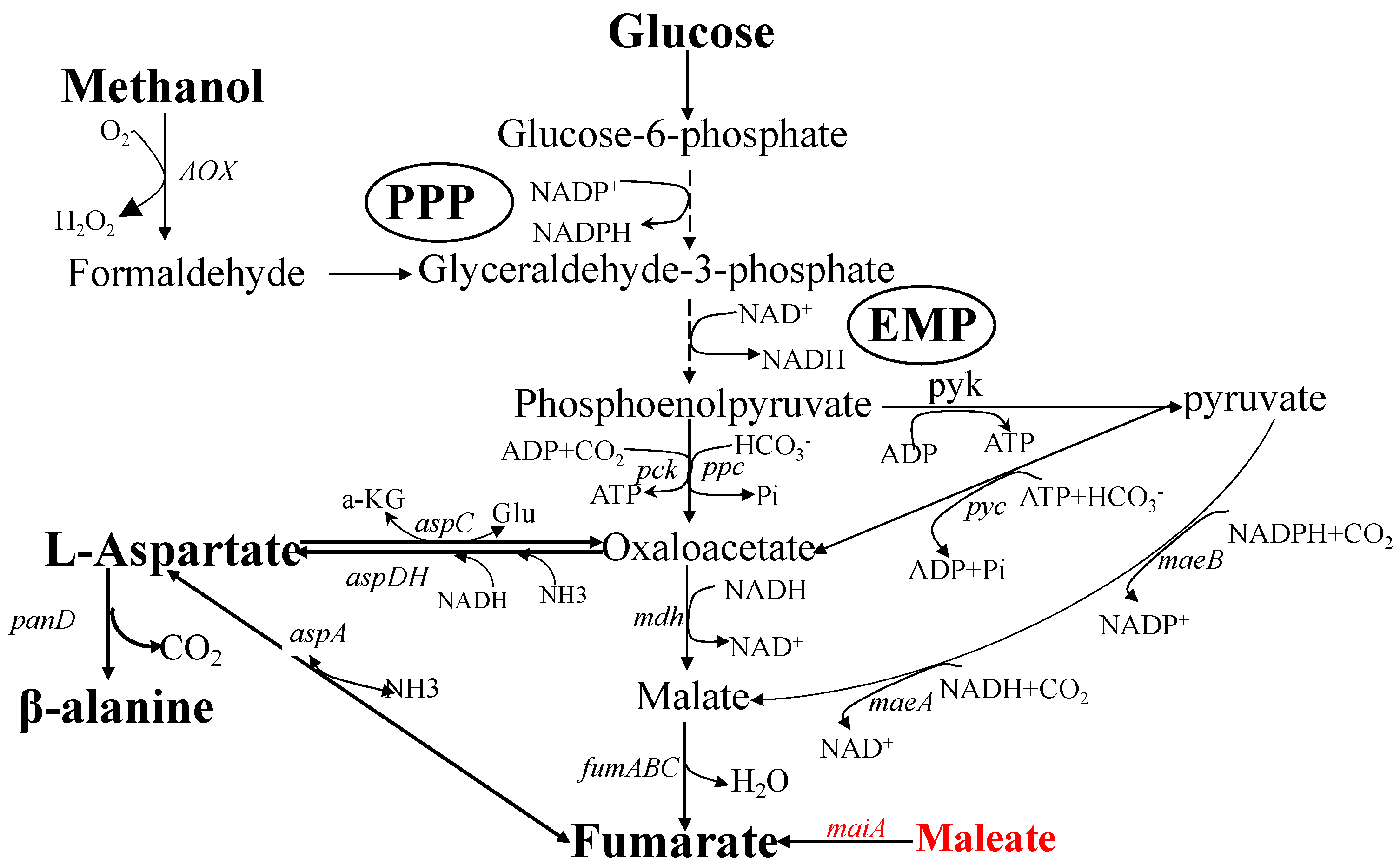
1. Introduction
2. Metabolic Engineering of Microbials to Produce L-Aspartate and Its Derivatives
2.1. Developing Cell Factories to Produce L-Aspartate

{kind=link}
{kind=link}
{kind=link}
| Organism | Metabolic Engineering Strategies | Substrate | Titer (g/L) | Yield a (g/g) | Fermentation Strategy | Reference |
|---|---|---|---|---|---|---|
| Engineered cell factories | ||||||
| E. coli XAR31 | Introducing and overexpressing CgaspC, Cgppc, Mspck, glk, bt-ca, acs, Cgasp, BsrocG and CR, deleting genes involved in byproduct biosynthesis, developing a cofactor self-sufficient system, optimizing the fermentation conditions | glucose | 33.1 | 0.39 | fed-batch | [15] |
| C. glutamicum SLV. pEKEx3-aspB | Deleting genes involved in byproduct biosynthesis (sdhCAB, ldhA and avtA) | glucose | ~5.72 a | 0.75 | flask | [18] |
| B. flavum 70 | Developing several mutations: a citrate synthase-defective glutamate auxotroph, S-(2-aminoethyl)-L-cysteine-resistant mutant, a methionine-insensitive revertant and hosphoenolpyruvate carboxylase, a supplement of biotin | glucose | 22.6 | 0.22 | flask | [16] |
| Enzyme catalysis | ||||||
| E. coli pMA-RBS4-G27A/G171A | Co-overexpressing maleate cis-trans isomerase (MaiA) mutant and aspartase (AspA) on the plasmid and optimizing their activity ratio by ribosome binding site (RBS) regulation | maleate | 419.8 | 0.72 | 5-L fermenter | [13] |
| E. coli JCL1258/pBAW2/pASP400 | Overexpressing aspC and tyrB on plasmid pASP400 and overexpressing parB and aspA on plasmid pBAW2 | fumarate | 77.60 a | 0.83 | [14] | |

2.2. Developing Cell Factories to Produce β-Alanine
2.3. Developing Cell Factories to Produce Ectoine
2.4. Developing Cell Factories to Produce 3-Hydroxypropionate
2.5. Current Process for Developing Cell Factories to Produce D-Pantothenic Acid
2.6. Developing Cell Factories to Produce L-Homoserine
3. Perspective
- (1)
- metabolic engineering of the bacteria to increase the titer, yield and productivity of the target products
- (2)
- fermentation condition optimization
- (3)
- downstream processing
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Choi, K.R.; Jang, W.D.; Yang, D.; Cho, J.S.; Park, D.; Lee, S.Y. Systems metabolic engineering strategies: Integrating systems and synthetic biology with metabolic engineering. Trends Biotechnol. 2019, 37, 817–837. [Google Scholar] [PubMed]
- Choi, K.Y.; Wernick, D.G.; Tat, C.A.; Liao, J.C. Consolidated conversion of protein waste into biofuels and ammonia using Bacillus subtilis. Metab. Eng. 2014, 23, 53–61. [Google Scholar] [PubMed]
- Angermayr, S.A.; Gorchs Rovira, A.; Hellingwerf, K.J. Metabolic engineering of cyanobacteria for the synthesis of commodity products. Trends Biotechnol. 2015, 33, 352–361. [Google Scholar] [PubMed]
- Antoniewicz, M.R. A guide to metabolic flux analysis in metabolic engineering: Methods, tools and applications. Metab. Eng. 2021, 63, 2–12. [Google Scholar] [PubMed]
- Huang, L.; Ho, C.T.; Wang, Y. Biosynthetic pathways and metabolic engineering of spice flavors. Crit. Rev. Food Sci. Nutr. 2021, 61, 2047–2060. [Google Scholar]
- Zhu, K.; Kong, J.; Zhao, B.; Rong, L.; Liu, S.; Lu, Z.; Zhang, C.; Xiao, D.; Pushpanathan, K.; Foo, J.L.; et al. Metabolic engineering of microbes for monoterpenoid production. Biotechnol. Adv. 2021, 53, 107837. [Google Scholar]
- Yang, D.; Park, S.Y.; Park, Y.S.; Eun, H.; Lee, S.Y. Metabolic engineering of Escherichia coli for natural product biosynthesis. Trends Biotechnol. 2020, 38, 745–765. [Google Scholar]
- Sinha, R.; Shukla, P. Current trends in protein engineering: Updates and progress. Curr. Protein Pept. Sci. 2019, 20, 398–407. [Google Scholar]
- Yarmush, M.L.; Banta, S. Metabolic engineering: Advances in modeling and intervention in health and disease. Annu. Rev. Biomed. Eng. 2003, 5, 349–381. [Google Scholar]
- Jarboe, L.R.; Zhang, X.; Wang, X.; Moore, J.C.; Shanmugam, K.T.; Ingram, L.O. Metabolic engineering for production of biorenewable fuels and chemicals: Contributions of synthetic biology. J. Biomed. Biotechnol. 2010, 2010, 761042. [Google Scholar]
- Han, M.; Zhang, C.; Suglo, P.; Sun, S.; Wang, M.; Su, T. L-aspartate: An essential metabolite for plant growth and stress acclimation. Molecules 2021, 26, 1887. [Google Scholar]
- Czarnecka, K.; Pilarz, A.; Rogut, A.; Maj, P.; Szymańska, J.; Olejnik, Ł.; Szymański, P. Aspartame-true or false? Narrative review of safety analysis of general use in products. Nutrients 2021, 13, 1957. [Google Scholar] [PubMed]
- Liu, Z.; Yu, L.; Zhou, L.; Zhou, Z. One-pot biosynthesis of L-aspartate from maleate via an engineered strain containing a dual-enzyme system. Appl. Environ. Microbiol. 2019, 85, 19. [Google Scholar]
- Chao, Y.P.; Lo, T.E.; Luo, N.S. Selective production of L-aspartic acid and L-phenylalanine by coupling reactions of aspartase and aminotransferase in Escherichia coli. Enzyme Microb. Technol. 2000, 27, 19–25. [Google Scholar] [CrossRef]
- Piao, X.; Wang, L.; Lin, B.; Chen, H.; Liu, W.; Tao, Y. Metabolic engineering of Escherichia coli for production of L-aspartate and its derivative β-alanine with high stoichiometric yield. Metab. Eng. 2019, 54, 244–254. [Google Scholar]
- Mori, M.; Shiio, I. Production of aspartic acid and enzymatic alteration in pyruvate kinase mutants of Brevibacterium flavum. Agric. Biol. Chem. 1984, 48, 9. [Google Scholar] [CrossRef]
- Tajima, T.; Hamada, M.; Nakashimada, Y.; Kato, J. Efficient aspartic acid production by a psychrophile-based simple biocatalyst. J. Ind. Microbiol. Biotechnol. 2015, 42, 1319–1324. [Google Scholar] [PubMed]
- Ziert, C. Metabolic Engineering of Corynebacterium glutamicum for the Production of L-Aspartate and Its Derivatives β-Alanine and Ectoine; Universitätsbibliothek Bielefeld: Bielefeld, Germany, 2014. [Google Scholar]
- Padhi, S.; Dash, M.; Sahu, R.; Panda, P. Urinary tract infection due to Paenibacillus alvei in a chronic kidney disease: A rare case report. J. Lab. Physicians 2013, 5, 133–135. [Google Scholar] [PubMed]
- Hoffman, J.R.; Varanoske, A.; Stout, J.R. Effects of β-Alanine supplementation on carnosine elevation and physiological performance. Adv. Food Nutr. Res. 2018, 84, 183–206. [Google Scholar]
- Kopec, W.; Jamroz, D.; Wiliczkiewicz, A.; Biazik, E.; Pudlo, A.; Korzeniowska, M.; Hikawczuk, T.; Skiba, T. Antioxidative characteristics of chicken breast meat and blood after diet supplementation with carnosine, L-histidine, and β-alanine. Antioxidants 2020, 9, 1093. [Google Scholar]
- Yuan, S.F.; Nair, P.H.; Borbon, D.; Coleman, S.M.; Fan, P.H.; Lin, W.L.; Alper, H.S. Metabolic engineering of E. coli for β-alanine production using a multi-biosensor enabled approach. Metab. Eng. 2022, 74, 24–35. [Google Scholar] [PubMed]
- Wang, P.; Zhou, H.Y.; Li, B.; Ding, W.Q.; Liu, Z.Q.; Zheng, Y.G. Multiplex modification of Escherichia coli for enhanced β-alanine biosynthesis through metabolic engineering. Bioresour. Technol. 2021, 342, 126050. [Google Scholar] [CrossRef] [PubMed]
- Ghiffary, M.R.; Prabowo, C.P.S.; Adidjaja, J.J.; Lee, S.Y.; Kim, H.U. Systems metabolic engineering of Corynebacterium glutamicum for the efficient production of β-alanine. Metab. Eng. 2022, 74, 121–129. [Google Scholar] [PubMed]
- Hu, S.; Fei, M.; Fu, B.; Yu, M.; Yuan, P.; Tang, B.; Yang, H.; Sun, D. Development of probiotic E. coli Nissle 1917 for β-alanine production by using protein and metabolic engineering. Appl. Microbiol. Biotechnol. 2023, 107, 2277–2288. [Google Scholar] [PubMed]
- Li, B.; Zhang, B.; Wang, P.; Cai, X.; Chen, Y.Y.; Yang, Y.F.; Liu, Z.Q.; Zheng, Y.G. Rerouting fluxes of the central carbon metabolism and relieving mechanism-based inactivation of l-Aspartate-α-decarboxylase for fermentative production of β-Alanine in Escherichia coli. ACS Synth. Biol. 2022, 11, 1908–1918. [Google Scholar]
- Zou, X.; Guo, L.; Huang, L.; Li, M.; Zhang, S.; Yang, A.; Zhang, Y.; Zhu, L.; Zhang, H.; Zhang, J.; et al. Pathway construction and metabolic engineering for fermentative production of β-alanine in Escherichia coli. Appl. Microbiol. Biotechnol. 2020, 104, 2545–2559. [Google Scholar]
- Wang, J.Y.; Rao, Z.M.; Xu, J.Z.; Zhang, W.G. Enhancing β-alanine production from glucose in genetically modified Corynebacterium glutamicum by metabolic pathway engineering. Appl. Microbiol. Biotechnol. 2021, 105, 9153–9166. [Google Scholar]
- Tadi SR, R.; Nehru, G.; Sivaprakasam, S. Metabolic Engineering of Bacillus megaterium for the Production of β-alanine. Biotechnol. Bioprocess Eng. 2022, 27, 909–920. [Google Scholar]
- Miao, L.; Li, Y.; Zhu, T. Metabolic engineering of methylotrophic Pichia pastoris for the production of β-alanine. Bioresour. Bioprocess. 2021, 8, 89. [Google Scholar] [CrossRef]
- Shen, Y.; Zhao, L.; Li, Y.; Zhang, L.; Shi, G. Synthesis of β-alanine from L-aspartate using L-aspartate-α-decarboxylase from Corynebacterium glutamicum. Biotechnol. Lett. 2014, 36, 1681–1686. [Google Scholar]
- Wang, L.; Piao, X.; Cui, S.; Hu, M.; Tao, Y. Enhanced production of β-alanine through co-expressing two different subtypes of L-aspartate-α-decarboxylase. J. Ind. Microbiol. Biotechnol. 2020, 47, 465–474. [Google Scholar]
- Tadi SR, R.; Nehru, G.; Sivaprakasam, S. One-Pot biosynthesis of 3-aminopropionic acid from fumaric acid using recombinant Bacillus megaterium Containing a linear dual-enzyme cascade. Appl. Biochem. Biotechnol. 2022, 194, 1740–1754. [Google Scholar]
- Bownik, A.; Stępniewska, Z. Ectoine as a promising protective agent in humans and animals. Arh Hig. Rada Toksikol. 2016, 67, 260–265. [Google Scholar] [PubMed]
- Guzmán, H.; Van-Thuoc, D.; Martín, J.; Hatti-Kaul, R.; Quillaguamán, J. A process for the production of ectoine and poly(3-hydroxybutyrate) by Halomonas boliviensis. Appl. Microbiol. Biotechnol. 2009, 84, 1069–1077. [Google Scholar] [CrossRef]
- Sattar, O.I.A.; Abuseada, H.H.M.; Emara, M.S.; Rabee, M. Green electrochemical and chromatographic quantifications of the extremolyte ectoine in halophilic bacterial cultures and related pharmaceutical preparations. J. Pharm. Biomed. Anal. 2022, 213, 114680. [Google Scholar]
- Peters, P.; Galinski, E.A.; Trüper, H.G. The biosynthesis of ectoine. FEMS Microbiol. Lett. 1990, 71, 157–162. [Google Scholar]
- Czech, L.; Höppner, A.; Kobus, S.; Seubert, A.; Riclea, R.; Dickschat, J.S.; Heider, J.; Smits SH, J.; Bremer, E. Illuminating the catalytic core of ectoine synthase through structural and biochemical analysis. Sci. Rep. 2019, 9, 364. [Google Scholar] [PubMed]
- Mantas MJ, Q.; Nunn, P.B.; Ke, Z.; Codd, G.A.; Barker, D. Genomic insights into the biosynthesis and physiology of the cyanobacterial neurotoxin 2,4-diaminobutanoic acid (2,4-DAB). Phytochemistry 2021, 192, 112953. [Google Scholar]
- Fallet, C.; Rohe, P.; Franco-Lara, E. Process optimization of the integrated synthesis and secretion of ectoine and hydroxyectoine under hyper/hypo-osmotic stress. Biotechnol. Bioeng. 2010, 107, 124–133. [Google Scholar] [PubMed]
- Zhang, H.; Liang, Z.; Zhao, M.; Ma, Y.; Luo, Z.; Li, S.; Xu, H. Metabolic engineering of Escherichia coli for ectoine production with a fermentation strategy of supplementing the amino donor. Front. Bioeng. Biotechnol. 2022, 10, 824859. [Google Scholar] [CrossRef]
- Ning, Y.; Wu, X.; Zhang, C.; Xu, Q.; Chen, N.; Xie, X. Pathway construction and metabolic engineering for fermentative production of ectoine in Escherichia coli. Metab. Eng. 2016, 36, 10–18. [Google Scholar] [PubMed]
- Kang, J.Y.; Lee, B.; Kim, J.A.; Kim, M.S.; Kim, C.H. Identification and characterization of an ectoine biosynthesis gene cluster from Aestuariispira ectoiniformans sp. nov., isolated from seawater. Microbiol. Res. 2022, 254, 126898. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Zhang, H.; Wang, X.; Ma, J.; Lei, P.; Xu, H.; Li, S. Enhancing ectoine production by recombinant Escherichia coli through step-wise fermentation optimization strategy based on kinetic analysis. Bioprocess Biosyst. Eng. 2021, 44, 1557–1566. [Google Scholar] [PubMed]
- He, Y.Z.; Gong, J.; Yu, H.Y.; Tao, Y.; Zhang, S.; Dong, Z.Y. High production of ectoine from aspartate and glycerol by use of whole-cell biocatalysis in recombinant Escherichia coli. Microb. Cell Fact. 2015, 14, 55. [Google Scholar]
- Chen, J.; Liu, P.; Chu, X.; Zhang, H.; Rowley, D.C.; Wang, H. Metabolic pathway construction and optimization of Escherichia coli for high-level ectoine production. Curr. Microbiol. 2020, 77, 1412–1418. [Google Scholar] [CrossRef]
- Gießelmann, G.; Dietrich, D.; Jungmann, L.; Kohlstedt, M.; Jeon, E.J.; Yim, S.S.; Sommer, F.; Zimmer, D.; Mühlhaus, T.; Schroda, M.; et al. Metabolic engineering of Corynebacterium glutamicum for high-level ectoine production: Design, combinatorial assembly, and implementation of a transcriptionally balanced heterologous ectoine pathway. Biotechnol. J. 2019, 14, e1800417. [Google Scholar] [CrossRef]
- Becker, J.; Schäfer, R.; Kohlstedt, M.; Harder, B.J.; Borchert, N.S.; Stöveken, N.; Bremer, E.; Wittmann, C. Systems metabolic engineering of Corynebacterium glutamicum for production of the chemical chaperone ectoine. Microb. Cell Fact. 2013, 12, 110. [Google Scholar]
- Jiang, A.; Song, Y.; You, J.; Zhang, X.; Xu, M.; Rao, Z. High-yield ectoine production in engineered Corynebacterium glutamicum by fine metabolic regulation via plug-in repressor library. Bioresour. Technol. 2022, 362, 127802. [Google Scholar]
- Zhao, Q.; Li, S.; Lv, P.; Sun, S.; Ma, C.; Xu, P.; Su, H.; Yang, C. High ectoine production by an engineered Halomonas hydrothermalis Y2 in a reduced salinity medium. Microb. Cell Fact. 2019, 18, 184. [Google Scholar]
- Ma, H.; Zhao, Y.; Huang, W.; Zhang, L.; Wu, F.; Ye, J.; Chen, G.Q. Rational flux-tuning of Halomonas bluephagenesis for co-production of bioplastic PHB and ectoine. Nat. Commun. 2020, 11, 3313. [Google Scholar]
- Salar-García, M.J.; Bernal, V.; Pastor, J.M.; Salvador, M.; Argandoña, M.; Nieto, J.J.; Vargas, C.; Cánovas, M. Understanding the interplay of carbon and nitrogen supply for ectoines production and metabolic overflow in high density cultures of Chromohalobacter salexigens. Microb. Cell Fact. 2017, 16, 23. [Google Scholar] [CrossRef] [PubMed]
- Nagata, S.; Wang, Y.; Oshima, A.; Zhang, L.; Miyake, H.; Sasaki, H.; Ishida, A. Efficient cyclic system to yield ectoine using Brevibacterium sp. JCM 6894 subjected to osmotic downshock. Biotechnol. Bioeng. 2008, 99, 941–948. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Onraedt, A.E.; Walcarius, B.A.; Soetaert, W.K.; Vandamme, E.J. Optimization of ectoine synthesis through fed-batch fermentation of Brevibacterium epidermis. Biotechnol. Prog. 2005, 21, 1206–1212. [Google Scholar] [CrossRef] [PubMed]
- Van-Thuoc, D.; Guzmán, H.; Thi-Hang, M.; Hatti-Kaul, R. Ectoine production by Halomonas boliviensis: Optimization using response surface methodology. Mar. Biotechnol. 2010, 12, 586–593. [Google Scholar] [CrossRef]
- Chen, W.C.; Hsu, C.C.; Lan, J.C.; Chang, Y.K.; Wang, L.F.; Wei, Y.H. Production and characterization of ectoine using a moderately halophilic strain Halomonas salina BCRC17875. J. Biosci. Bioeng. 2018, 125, 578–584. [Google Scholar] [CrossRef]
- Chen, S.Y.; Peng, T.C.; Huang, S.Z.; Chien, C.C. Isolation of an ectoine-producing Sinobaca sp. and identification of genes that are involved in ectoine biosynthesis. FEMS Microbiol. Lett. 2022, 369, fnac046. [Google Scholar] [CrossRef]
- Zhang, L.H.; Lang, Y.J.; Nagata, S. Efficient production of ectoine using ectoine-excreting strain. Extremophiles 2009, 13, 717–724. [Google Scholar] [CrossRef]
- Fatollahi, P.; Ghasemi, M.; Yazdian, F.; Sadeghi, A. Ectoine production in bioreactor by Halomonas elongata DSM2581: Using MWCNT and Fe-nanoparticle. Biotechnol. Prog. 2021, 37, e3073. [Google Scholar] [CrossRef]
- Chen, W.C.; Yuan, F.W.; Wang, L.F.; Chien, C.C.; Wei, Y.H. Ectoine production with indigenous Marinococcussp. MAR2 isolated from the marine environment. Prep. Biochem. Biotechnol. 2020, 50, 74–81. [Google Scholar] [CrossRef]
- Lang, Y.J.; Bai, L.; Ren, Y.N.; Zhang, L.H.; Nagata, S. Production of ectoine through a combined process that uses both growing and resting cells of Halomonas salina DSM 5928T. Extremophiles 2011, 15, 303–310. [Google Scholar] [CrossRef]
- Wei, Y.H.; Yuan, F.W.; Chen, W.C.; Chen, S.Y. Production and characterization of ectoine by Marinococcus sp. ECT1 isolated from a high-salinity environment. J. Biosci. Bioeng. 2011, 111, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Nagata, S.; Wang, Y.B. Accumulation of ectoine in the halotolerant Brevibacterium sp. JCM 6894. J. Biosci. Bioeng. 2001, 91, 288–293. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; Gao, X.; Xing, J.; Wang, R.; Zhu, D.; Shen, G. Comparative genomic analysis of Halomonas campaniensis wild-type and ultraviolet radiation-mutated strains reveal genomic differences associated with increased ectoine production. Int. Microbiol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Wang, Z.; Wang, J.; Mohisn, A.; Liu, H.; Zhang, Y.; Zhuang, Y.; Guo, M. Physiological metabolic topology analysis of Halomonas elongata DSM 2581. Biotechnol. Bioeng. 2022, 119, 3509–3525. [Google Scholar] [CrossRef] [PubMed]
- Van-Thuoc, D.; Guzmán, H.; Quillaguamán, J.; Hatti-Kaul, R. High productivity of ectoines by Halomonas boliviensis using a combined two-step fed-batch culture and milking process. J. Biotechnol. 2010, 147, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Ayadi, H.; Frikha-Dammak, D.; Fakhfakh, J.; Chamkha, M.; Hassairi, I.; Allouche, N.; Sayadi, S.; Maalej, S. The saltern-derived Paludifilum halophilum DSM 102817T is a new high-yield ectoines producer in minimal medium and under salt stress conditions. 3 Biotech 2020, 10, 533. [Google Scholar] [CrossRef]
- Chen, R.; Zhu, L.; Lv, L.; Yao, S.; Li, B.; Qian, J. Optimization of the extraction and purification of the compatible solute ectoine from Halomonas elongate in the laboratory experiment of a commercial production project. World J. Microbiol. Biotechnol. 2017, 33, 116. [Google Scholar] [CrossRef]
- Yu, W.; Cao, X.; Gao, J.; Zhou, Y.J. Overproduction of 3-hydroxypropionate in a super yeast chassis. Bioresour. Technol. 2022, 361, 127690. [Google Scholar] [CrossRef]
- Choi, S.; Song, C.W.; Shin, J.H.; Lee, S.Y. Biorefineries for the production of top building block chemicals and their derivatives. Metab. Eng. 2015, 28, 223–239. [Google Scholar] [CrossRef]
- Della Pina, C.; Falletta, E.; Rossi, M. A green approach to chemical building blocks. The case of 3-hydroxypropanoic acid. Green Chem. 2011, 13, 1624–1632. [Google Scholar] [CrossRef]
- Zhao, P.; Ma, C.; Xu, L.; Tian, P. Exploiting tandem repetitive promoters for high-level production of 3-hydroxypropionic acid. Appl. Microbiol. Biotechnol. 2019, 103, 4017–4031. [Google Scholar] [CrossRef]
- Kim, J.W.; Ko, Y.S.; Chae, T.U.; Lee, S.Y. High-level production of 3-hydroxypropionic acid from glycerol as a sole carbon source using metabolically engineered Escherichia coli. Biotechnol. Bioeng. 2020, 117, 2139–2152. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Su, M.; Chen, Y.; Wang, Z.; Nielsen, J.; Liu, Z. Engineering yeast mitochondrial metabolism for 3-hydroxypropionate production. Biotechnol. Biofuels Bioprod. 2023, 16, 64. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.R.; Yan, X.; Yu, L.P.; Liu, X.Y.; Chen, G.Q. Hyperproduction of 3-hydroxypropionate by Halomonas bluephagenesis. Nat. Commun. 2021, 12, 1513. [Google Scholar] [CrossRef]
- Liu, B.; Xiang, S.; Zhao, G.; Wang, B.; Ma, Y.; Liu, W.; Tao, Y. Efficient production of 3-hydroxypropionate from fatty acids feedstock in Escherichia coli. Metab. Eng. 2019, 51, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Niu, K.; Xiong, T.; Qin, H.B.; Wu, H.; Liu, Z.Q.; Zheng, Y.G. 3-Hydroxypropionic acid production by recombinant Escherichia coli ZJU-3HP01 using glycerol-glucose dual-substrate fermentative strategy. Biotechnol. Appl. Biochem. 2017, 64, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Sankaranarayanan, M.; Ashok, S.; Park, S. Production of 3-hydroxypropionic acid from glycerol by acid tolerant Escherichia coli. J. Ind. Microbiol. Biotechnol. 2014, 41, 1039–1050. [Google Scholar] [CrossRef] [PubMed]
- Chaves, G.L.; Batista, R.S.; Cunha, J.S.; Oliveira, D.B.; da Silva, M.R.; Pisani GF, D.; Selistre-de-Araújo, H.S.; Zangirolami, T.C.; da Silva, A.J. Improving 3-hydroxypropionic acid production in E. coli by in silico prediction of new metabolic targets. New Biotechnol. 2022, 72, 80–88. [Google Scholar] [CrossRef]
- Heo, W.; Kim, J.H.; Kim, S.; Kim, K.H.; Kim, H.J.; Seo, J.H. Enhanced production of 3-hydroxypropionic acid from glucose and xylose by alleviation of metabolic congestion due to glycerol flux in engineered Escherichia coli. Bioresour. Technol. 2019, 285, 121320. [Google Scholar] [CrossRef]
- Dai, Z.; Pomraning, K.R.; Deng, S.; Kim, J.; Campbell, K.B.; Robles, A.L.; Hofstad, B.A.; Munoz, N.; Gao, Y.; Lemmon, T.; et al. Metabolic engineering to improve production of 3-hydroxypropionic acid from corn-stover hydrolysate in Aspergillus species. Biotechnol. Biofuels Bioprod. 2023, 16, 53. [Google Scholar] [CrossRef]
- Liang, B.; Zhang, X.; Meng, C.; Wang, L.; Yang, J. Directed evolution of tripartite ATP-independent periplasmic transporter for 3-Hydroxypropionate biosynthesis. Appl. Microbiol. Biotechnol. 2023, 107, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Ding, Y.; Zhang, R.; Liu, H.; Xian, M.; Zhao, G. Functional balance between enzymes in malonyl-CoA pathway for 3-hydroxypropionate biosynthesis. Metab. Eng. 2016, 34, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Rathnasingh, C.; Raj, S.M.; Jo, J.E.; Park, S. Development and evaluation of efficient recombinant Escherichia coli strains for the production of 3-hydroxypropionic acid from glycerol. Biotechnol. Bioeng. 2009, 104, 729–739. [Google Scholar] [PubMed]
- Kwak, S.; Park, Y.C.; Seo, J.H. Biosynthesis of 3-hydroxypropionic acid from glycerol in recombinant Escherichia coli expressing Lactobacillus brevis dhaB and dhaR gene clusters and E. coli K-12 aldH. Bioresour. Technol. 2013, 135, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.S.; Kim, Y.S.; Lee, C.M.; Lee, J.H.; Jung, W.S.; Ahn, J.H.; Song, S.H.; Choi, I.S.; Cho, K.M. Metabolic engineering of 3-hydroxypropionic acid biosynthesis in Escherichia coli. Biotechnol. Bioeng. 2015, 112, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Jung, I.Y.; Lee, J.W.; Min, W.K.; Park, Y.C.; Seo, J.H. Simultaneous conversion of glucose and xylose to 3-hydroxypropionic acid in engineered Escherichia coli by modulation of sugar transport and glycerol synthesis. Bioresour. Technol. 2015, 198, 709–716. [Google Scholar] [CrossRef]
- Lee, T.Y.; Min, W.K.; Kim, H.J.; Seo, J.H. Improved production of 3-hydroxypropionic acid in engineered Escherichia coli by rebalancing heterologous and endogenous synthetic pathways. Bioresour. Technol. 2020, 299, 122600. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Wang, Y.; Xu, M.; Fei, Q.; Gu, Y.; Luo, Y.; Wu, H. Efficient biosynthesis of 3-hydroxypropionic acid from ethanol in metabolically engineered Escherichia coli. Bioresour. Technol. 2022, 363, 127907. [Google Scholar] [CrossRef]
- Kildegaard, K.R.; Jensen, N.B.; Schneider, K.; Czarnotta, E.; Özdemir, E.; Klein, T.; Maury, J.; Ebert, B.E.; Christensen, H.B.; Chen, Y.; et al. Engineering and systems-level analysis of Saccharomyces cerevisiae for production of 3-hydroxypropionic acid via malonyl-CoA reductase-dependent pathway. Microb. Cell Fact. 2016, 15, 53. [Google Scholar] [CrossRef]
- Fina, A.; Brêda, G.C.; Pérez-Trujillo, M.; Freire DM, G.; Almeida, R.V.; Albiol, J.; Ferrer, P. Benchmarking recombinant Pichia pastoris for 3-hydroxypropionic acid production from glycerol. Microb. Biotechnol. 2021, 14, 1671–1682. [Google Scholar] [CrossRef]
- Luo, L.H.; Seo, J.W.; Heo, S.Y.; Oh, B.R.; Kim, D.H.; Kim, C.H. Identification and characterization of Klebsiella pneumoniae aldehyde dehydrogenases increasing production of 3-hydroxypropionic acid from glycerol. Bioprocess Biosyst. Eng. 2013, 36, 1319–1326. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Li, Y.; Ge, X.; Tian, P. 3-Hydroxypropionaldehyde-specific aldehyde dehydrogenase from Bacillus subtilis catalyzes 3-hydroxypropionic acid production in Klebsiella pneumoniae. Biotechnol. Lett. 2015, 37, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Xian, M.; Liu, W.; Xu, C.; Zhang, H.; Zhao, G. Biosynthesis of poly(3-hydroxypropionate) from glycerol using engineered Klebsiella pneumoniae strain without vitamin B12. Bioengineered 2015, 6, 77–81. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, X.; Chen, L.; Wang, X.; Tian, P. Physiological investigations of the influences of byproduct pathways on 3-hydroxypropionic acid production in Klebsiella pneumoniae. J. Basic Microbiol. 2019, 59, 1195–1207. [Google Scholar] [CrossRef]
- Zhao, P.; Ren, M.; Ge, X.; Tian, P.; Tan, T. Development of orthogonal T7 expression system in Klebsiella pneumoniae. Biotechnol. Bioeng. 2020, 117, 2446–2459. [Google Scholar] [CrossRef]
- Wang, X.; Sa, N.; Wang, F.H.; Tian, P.F. Engineered constitutive pathway in Klebsiella pneumoniae for 3-hydroxypropionic acid production and implications for decoupling glycerol dissimilation pathways. Curr. Microbiol. 2013, 66, 293–299. [Google Scholar] [CrossRef]
- Luo, L.H.; Seo, J.W.; Baek, J.O.; Oh, B.R.; Heo, S.Y.; Hong, W.K.; Kim, D.H.; Kim, C.H. Identification and characterization of the propanediol utilization protein PduP of Lactobacillus reuteri for 3-hydroxypropionic acid production from glycerol. Appl. Microbiol. Biotechnol. 2011, 89, 697–703. [Google Scholar] [CrossRef]
- Ashok, S.; Sankaranarayanan, M.; Ko, Y.; Jae, K.E.; Ainala, S.K.; Kumar, V.; Park, S. Production of 3-hydroxypropionic acid from glycerol by recombinant Klebsiella pneumoniae ΔdhaTΔyqhD which can produce vitamin B12 naturally. Biotechnol. Bioeng. 2013, 110, 511–524. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Ge, X.; Tian, P. High Production of 3-Hydroxypropionic acid in Klebsiella pneumoniae by systematic optimization of glycerol metabolism. Sci. Rep. 2016, 6, 26932. [Google Scholar] [CrossRef]
- Suyama, A.; Higuchi, Y.; Urushihara, M.; Maeda, Y.; Takegawa, K. Production of 3-hydroxypropionic acid via the malonyl-CoA pathway using recombinant fission yeast strains. J. Biosci. Bioeng. 2017, 124, 392–399. [Google Scholar] [CrossRef]
- Takayama, S.; Ozaki, A.; Konishi, R.; Otomo, C.; Kishida, M.; Hirata, Y.; Matsumoto, T.; Tanaka, T.; Kondo, A. Enhancing 3-hydroxypropionic acid production in combination with sugar supply engineering by cell surface-display and metabolic engineering of Schizosaccharomyces pombe. Microb. Cell Fact. 2018, 17, 176. [Google Scholar] [CrossRef] [PubMed]
- Suppuram, P.; Ramakrishnan, G.G.; Subramanian, R. An integrated process for the production of 1,3-propanediol, lactate and 3-hydroxypropionic acid by an engineered Lactobacillus reuteri. Biosci. Biotechnol. Biochem. 2019, 83, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Burgé, G.; Saulou-Bérion, C.; Moussa, M.; Pollet, B.; Flourat, A.; Allais, F.; Athès, V.; Spinnler, H.E. Diversity of Lactobacillus reuteri strains in converting glycerol into 3-hydroxypropionic acid. Appl. Biochem. Biotechnol. 2015, 177, 923–939. [Google Scholar] [CrossRef]
- Li, W.; Wang, T.; Dong, Y.; Li, T. Screening, identification, and low-energy ion modified breeding of a yeast strain producing high level of 3-hydroxypropionic acid. Microbiologyopen 2020, 9, e00956. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Park, S.J.; Park, O.J.; Cho, J.; Rhee, J.W. Production of 3-hydroxypropionic acid from acrylic acid by newly isolated rhodococcus erythropolis LG12. J. Microbiol. Biotechnol. 2009, 19, 474–481. [Google Scholar] [CrossRef]
- Russmayer, H.; Ergoth, S.; Marx, H.; Sauer, M. Process engineering towards an oxidative cellular state improves 3-hydroxypropionic acid production with Lentilactobacillus diolivorans. Bioresour. Technol. 2023, 382, 129160. [Google Scholar] [CrossRef]
- Sun, L.; Yu, F.; Zheng, Y. [Biosynthesis of 3-hydroxypropionic acid from 1,3-propanediol by Gluconobacter oxydans ZJB09112]. Sheng Wu Gong Cheng Xue Bao 2012, 28, 498–507. [Google Scholar]
- Zhao, L.; Lin, J.; Wang, H.; Xie, J.; Wei, D. Development of a two-step process for production of 3-hydroxypropionic acid from glycerol using Klebsiella pneumoniae and Gluconobacter oxydans. Bioprocess Biosyst. Eng. 2015, 38, 2487–2495. [Google Scholar] [CrossRef]
- Tahiliani, A.G.; Beinlich, C.J. Pantothenic acid in health and disease. Vitam. Horm. 1991, 46, 165–228. [Google Scholar]
- Zhao, K.; Tang, H.; Zhang, B.; Zou, S.; Liu, Z.; Zheng, Y. Microbial production of vitamin B5: Current status and prospects. Crit. Rev. Biotechnol. 2022, 9, 1–21. [Google Scholar] [CrossRef]
- Liu, Z.; Sun, Z. Cloning and expression of D-lactonohydrolase cDNA from Fusarium moniliforme in Saccharomyces cerevisiae. Biotechnol. Lett. 2004, 26, 1861–1865. [Google Scholar] [CrossRef] [PubMed]
- Tigu, F.; Zhang, J.; Liu, G.; Cai, Z.; Li, Y. A highly active pantothenate synthetase from Corynebacterium glutamicum enables the production of D-pantothenic acid with high productivity. Appl. Microbiol. Biotechnol. 2018, 102, 6039–6046. [Google Scholar] [CrossRef] [PubMed]
- Tadi SR, R.; Nehru, G.; Allampalli SS, P.; Sivaprakasam, S. Engineering precursor and co-factor supply to enhance D-pantothenic acid production in Bacillus megaterium. Bioprocess Biosyst. Eng. 2022, 45, 843–854. [Google Scholar] [CrossRef]
- Tadi SR, R.; Nehru, G.; Limaye, A.M.; Sivaprakasam, S. High-level expression and optimization of pantoate-β-alanine ligase in Bacillus megaterium for the enhanced biocatalytic production of D-pantothenic acid. J. Food Sci. Technol. 2022, 59, 917–926. [Google Scholar] [CrossRef]
- Li, B.; Zhang, B.; Wang, P.; Cai, X.; Tang, Y.Q.; Jin, J.Y.; Liang, J.X.; Liu, Z.Q.; Zheng, Y.G. Targeting metabolic driving and minimization of by-products synthesis for high-yield production of D-pantothenate in Escherichia coli. Biotechnol. J. 2022, 17, e2100431. [Google Scholar] [PubMed]
- Zou, S.P.; Wang, Z.J.; Zhao, K.; Zhang, B.; Niu, K.; Liu, Z.Q.; Zheng, Y.G. High-level production of d-pantothenic acid from glucose by fed-batch cultivation of Escherichia coli. Biotechnol. Appl. Biochem. 2021, 68, 1227–1235. [Google Scholar] [CrossRef]
- Zou, S.; Zhao, K.; Tang, H.; Zhang, Z.; Zhang, B.; Liu, Z.; Zheng, Y. Improved production of D-pantothenic acid in Escherichia coli by integrated strain engineering and fermentation strategies. J. Biotechnol. 2021, 339, 65–72. [Google Scholar] [CrossRef]
- Cai, X.; Shi, X.; Liu, S.Q.; Qiang, Y.; Shen, J.D.; Zhang, B.; Liu, Z.Q.; Zheng, Y.G. Hot spot-based engineering of ketopantoate hydroxymethyltransferase for the improvement of D-pantothenic acid production in Escherichia coli. J. Biotechnol. 2023, 364, 40–49. [Google Scholar] [CrossRef]
- Zou, S.P.; Zhao, K.; Wang, Z.J.; Zhang, B.; Liu, Z.Q.; Zheng, Y.G. Overproduction of D-pantothenic acid via fermentation conditions optimization and isoleucine feeding from recombinant Escherichia coli W3110. 3 Biotech 2021, 11, 295. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, X.M.; Wang, W.; Liu, Z.Q.; Zheng, Y.G. Metabolic engineering of Escherichia coli for d-pantothenic acid production. Food Chem. 2019, 294, 267–275. [Google Scholar]
- Wang, P.; Zhou, H.Y.; Zhou, J.P.; Li, B.; Liu, Z.Q.; Zheng, Y.G. Module engineering coupled with omics strategies for enhancing D-pantothenate production in Escherichia coli. Bioresour. Technol. 2022, 352, 127024. [Google Scholar] [CrossRef] [PubMed]
- Su, R.; Wang, T.; Bo, T.; Cai, N.; Yuan, M.; Wu, C.; Jiang, H.; Peng, H.; Chen, N.; Li, Y. Enhanced production of D-pantothenic acid in Corynebacterium glutamicum using an efficient CRISPR-Cpf1 genome editing method. Microb. Cell Fact. 2023, 22, 3. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Sun, X.; Yuan, Y.; Chen, Q.; Ou, Z.; Deng, Z.; Ma, T.; Liu, T. Metabolic Engineering of Saccharomyces cerevisiae for Vitamin B5 Production. J. Agric. Food Chem. 2023, 71, 7408–7417. [Google Scholar] [CrossRef]
- Li, H.; Wang, B.S.; Li, Y.R.; Zhang, L.; Ding, Z.Y.; Gu, Z.H.; Shi, G.Y. Metabolic engineering of Escherichia coli W3110 for the production of L-methionine. J. Ind. Microbiol. Biotechnol. 2017, 44, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Vo, T.M.; Park, S. Metabolic engineering of Escherichia coli W3110 for efficient production of homoserine from glucose. Metab. Eng. 2022, 73, 104–113. [Google Scholar] [CrossRef]
- Muller, M.; Lee, C.M.; Gasiunas, G.; Davis, T.H.; Cradick, T.J.; Siksnys, V.; Bao, G.; Cathomen, T.; Mussolino, C. Streptococcus thermophilus CRISPR-Cas9 systems enable specific editing of the human genome. Mol. Ther. 2016, 24, 636–644. [Google Scholar] [CrossRef]
- Mu, Q.; Zhang, S.; Mao, X.; Tao, Y.; Yu, B. Highly efficient production of L-homoserine in Escherichia coli by engineering a redox balance route. Metab. Eng. 2021, 67, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Li, L.; Yu, S.; Zhou, J. Dual-channel glycolysis balances cofactor supply for L-homoserine biosynthesis in Corynebacterium glutamicum. Bioresour. Technol. 2023, 369, 128473. [Google Scholar] [CrossRef]
- Zhang, Y.; Wei, M.; Zhao, G.; Zhang, W.; Li, Y.; Lin, B.; Xu, Q.; Chen, N.; Zhang, C. High-level production of L-homoserine using a non-induced, non-auxotrophic Escherichia coli chassis through metabolic engineering. Bioresour. Technol. 2021, 327, 124814. [Google Scholar] [CrossRef]
- Cai, M.; Zhao, Z.; Li, X.; Xu, Y.; Xu, M.; Rao, Z. Development of a nonauxotrophic L-homoserine hyperproducer in Escherichia coli by systems metabolic engineering. Metab. Eng. 2022, 73, 270–279. [Google Scholar] [CrossRef]
- Liu, M.; Lou, J.; Gu, J.; Lyu, X.M.; Wang, F.Q.; Wei, D.Z. Increasing L-homoserine production in Escherichia coli by engineering the central metabolic pathways. J. Biotechnol. 2020, 314–315, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.Y.; Wang, F.Q.; Zhao, J.; Tao, X.Y.; Liu, M.; Wei, D.Z. Engineering Escherichia coli for L-homoserine production. J. Basic Microbiol. 2023, 63, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Xu, S.; Du, G.; Chen, J.; Zhou, J. Efficient production of L-homoserine in Corynebacterium glutamicum ATCC 13032 by redistribution of metabolic flux. Biochem. Eng. J. 2020, 161, 107665. [Google Scholar] [CrossRef]
- Plachý, J.; Ulbert, S.; Pelechová, J.; Krumphanzl, V. Fermentation production of L-homoserine by Corynebacterium sp. and its possible use in the preparation of threonine and lysine. Folia Microbiol. 1985, 30, 485–492. [Google Scholar] [CrossRef]
- Shi, A.; Zhu, X.; Lu, J.; Zhang, X.; Ma, Y. Activating transhydrogenase and NAD kinase in combination for improving isobutanol production. Metab. Eng. 2013, 16, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fei, M.; Mao, X.; Chen, Y.; Lu, Y.; Wang, L.; Yang, J.; Qiu, J.; Sun, D. Development of a dual-fluorescence reporter system for high-throughput screening of L-aspartate-α-decarboxylase. Acta. Biochim. Biophys. Sin. 2020, 52, 1420–1426. [Google Scholar] [CrossRef] [PubMed]
- Strecker, A.; Schubert, C.; Zedler, S.; Steinmetz, P.; Unden, G. DcuA of aerobically grown Escherichia coli serves as a nitrogen shuttle (L-aspartate/fumarate) for nitrogen uptake. Mol. Microbiol. 2018, 109, 801–811. [Google Scholar] [CrossRef]
- Zhang, C.; Sharma, S.; Wang, W.; Zeng, A.P. A novel downstream process for highly pure 1,3-propanediol from an efficient fed-batch fermentation of raw glycerol by Clostridium pasteurianum. Eng. Life Sci. 2021, 21, 351–363. [Google Scholar] [CrossRef]


| Organism | Metabolic Engineering Strategies | Substrate | Titer (g/L) | Yield (g/g) | Productivity (g/(L.h)) | Fermentation Strategy | Reference |
|---|---|---|---|---|---|---|---|
| Engineered cell factories | |||||||
| E. coli XBR41 | Introducing BspanD, deleting genes involved in byproduct biosynthesis, developing a cofactor self-sufficient system, optimizing the fermentation conditions | glucose | 37.7 | 0.75 a | Fed-batch | [15] | |
| E. coli NL-A13 | High-throughput method to screen L-aspartate-α-decarboxylase variant ADCK43Y, evaluation and elevation cells’ tolerance to β-alanine, improving fumarate supply and strengthening the pathway of fumarate and OAA to L-aspartate, optimizing culture medium | glucose glycerol | 11.9 | Fed-batch | [25] | ||
| E. coli W3110 | Introducing panD from Bacillus subtilis, rerouting fluxes of the central carbon metabolism, relieving the inactivation of L-aspartate-α-decarboxylase, optimizing the fed-batch bioprocess | glucose | 85.18 | 0.24 | 1.05 | Fed-batch | [26] |
| E. coli ALA17/pTrc99a-panDBS-aspBCG | Introducing panD from B.subtilis and aspB from C. glutamicum, inactivating the β-alanine uptake system, the aspartate kinase I and III, iclR, ptsG, aroG, galR, overexpressing ppc, aspC, aceB, aceA, glk, and gltBD operon | glucose | 43.94 | 0.20 | Fed-batch | [23] | |
| E. coli W FZβA-10 | Introducing an L-aspartate a-decarboxylase gene from Bacillus tequilensis, a L-aspartate dehydrogenase gene from Pseudomonas aeruginosa and a pyruvate decarboxylase from Corynebacterium glutamicum, overexpressing aspA, deleting three native L-aspartate kinase genes and genes for byproduct biosynthesis (ldhA, pflB, pta and adhE) | glucose | 43.12 | 0.89 | Fed-batch | [27] | |
| E. coli eBA32 | Biosensor-enabled high-throughput screening, cofactor balancing and pathway modification | glucose | 34.8 | Fed-batch | |||
| C. glutamicum XQ-5 | Deleting byproduct biosynthesis pathway (lactate dehydrogenase and alanine/valine aminotransferases), replacing L-aspartate kinase (AK) with wild-type AK, introducing and overexpressing a mutated L-aspartate-α-decarboxylase (BsADCE56S/I88M) from B. subtilis | glucose | 56.5 | 39.5% b | 0.79 | Fed-batch | [28] |
| C. glutamicum BAL10 (pBA2_tr18) | Introducing panD from B. subtilis, overexpressing PTS-independent glucose uptake system, ppc, pyc, aspB, rocG from B. subtilis, aspA from E. coli and β-alanine exporter (NCgl0580), replacing the native pck with that from Mannheimia succiniciproducens (encoded by Mspck), deleting odx and mdh | glucose | 166.6 | 0.28 | 1.74 | Fed-batch | [24] |
| B. megaterium BMDBPG | Introducing a codon-optimized panD from B. subtilis, overexpressing aspB, ppc and NADH-dependent glutamate dehydrogenase (gdh) | glucose | 17.60 | 0.23 | 0.78 | [29] | |
| methylotrophic Pichia pastoris 2ADC-Spe | Overexpressing panD from B. subtilis and aspDH from S. proteamaculans | methanol | 5.6 | Fed-batch | [30] | ||
| Enzyme catalysis | |||||||
| L-aspartate-α-decarboxylase (ADC) from C. glutamicum | Overexpressed in Escherichia coli BL21(DE3), optimal at 55 °C and pH 6 with excellent stability at 16–37 °C and pH 4–7 | L-aspartate | 12.85 | 97.2% b | Purified enzyme | [31] | |
| E.coli BTW | Co-expressing two different types of L-aspartate-α-decarboxylase: one was from B. subtilis and the other was from Tribolium castaneum | L-aspartate | 271.5 | 92.4% b | Whole cell | [32] | |
| E.coli BTEW | Co-expressing three enzymes: two types of L-aspartate-α-decarboxylase (one was from B. subtilis and the other was from Tribolium castaneum) and one type of L-aspartase (AspA) from E. coli | fumarate | 200.3 | 90.0% b | Whole cell | [32] | |
| B. megaterium BMDA-6 | Balancing the expression of L-aspartate-1-decarboxylases (ADC) from B. subtilis and aspartate ammonia-lyase (AspA) from B. megaterium, optimizing the cultivation conditions and biocatalysis process parameters | fumarate | 11.68 | 0.78 | Whole cell | [33] | |
| Organism | Metabolic Engineering Strategies | Substrate | Titer (g/L) | Yield (g/g) | Productivity (g/(L.h)) | Fermentation Strategy | Reference |
|---|---|---|---|---|---|---|---|
| Engineered cell factories | |||||||
| E. coli ET11 (ectA:ectB:ectC = 1:2:1) | Introducing the ectABC gene cluster from Halomonas venusta ZH, regulating the copy number of ectA, ectB and ectC, eliminating byproduct metabolic pathways, optimizing the culture medium | glucose | 53.2 | 0.33 | 1.11 | fed-batch | [41] |
| E. coli Ect05 | Introducing ectABC gene cluster from Halomonas elongata and a feedback-resistant L-aspartate kinase (lysC) from Corynebacterium glutamicum, deleting thrA and iclR, improving ppc expression by promoter replacement | glucose | 25.1 | 0.11 | 0.84 | fed-batch | [42] |
| E. coli S16-ectBAC | Introducing ectABC gene cluster from Aestuariispira SWCN16T into E. coli BL21 | sodium aspartate and glycerol | 2.26 | cell suspension bioconversion reactions in the optimum buffer | [43] | ||
| E. coli ET01 | Introducing the ectABC operon from Halomonas venusta ZH, optimizing the fermentation process | glucose | 47.8 | fed-batch | [44] | ||
| E. coli BW25113 | Introducing ectABC from Halomonas elongata and overexpressing these three genes with an arabinose-inducible promoter, optimizing the fermentation process | aspartate and glycerol | 25.1 | 4.1 b | 1.04 | whole-cell catalysis | [45] |
| E. coli ECT2 | Introducing the ectABC genes from Halomonas elongata, deleting lysA | glycerol and sodium aspartate | 12.7 | 1.27 | 0.53 | whole-cell catalysis | [46] |
| C. glutamicum ectABCopt | Introducing the ectABC genes from Pseudomonas stutzeri and regulating their expression with different promoters and three linker elements | glucose, sucrose and fructose | 65 | 0.19 | 2.3 f | fed-batch | [47] |
| C. glutamicum ECT-2 | Introducing a codon-optimized synthetic ectABCD gene cluster from Pseudomonas stutzeri, inactivating the L-lysine exporter, optimizing the fed-batch process | glucose | 4.5 | 0.24 d | 0.28 e | fed-batch | [48] |
| C.glutamicum CB5L6 | Introducing the ectBAC cluster from Pseudomonas stutzeri, deleting pck, ldh and sugR, improving the precursor supply (overexpression of Ecasd and CglysCS301Y), constructing repressor libraries (BetI from E. coli and LmrA from B. subtilis) | glucose | 45.52 | 0.25 | fed-batch | [49] | |
| H. hydrothermalis Y2/ΔectD/ΔdoeA | Identifying the pathways for ectoine synthesis and catabolism, deleting genes involved in ectoine catabolism (EctD and DoeA) and Na+/H+ antiporter (Mrp), optimizing the culture medium | monosodium glutamate | 10.5 | 0.21 | fed-batch | [50] | |
| H. bluephagenesis TD-ADEL-58 | Overexpressing three clusters related to ectoine biosynthesis, including ectABC, lysC and asd, deleting byproduct biosynthetic pathways | glucose | 28 | 0.21 | 1.0 | fed-batch | [51] |
| Natural producers | |||||||
| Chromohalobacter salexigens | Optimizing the medium composition, especially the C/N ratio, to regulate the metabolic pattern | glucose | 4.21 | fed-batch | [52] | ||
| Chromohalobacter salexigens | Producing ectoine with two continuously operated bioreactors, regulating the hyperosmotic conditions and thermal stress | glucose | 8.2 | 2.1 | fed-batch | [40] | |
| Brevibacterium sp. JCM 6894 | Inducing ectoine biosynthesis with 2 M NaCl, fermentation with non-sterilized medium | polypepton and dried yeast extract | 2.4 | flask | [53] | ||
| Brevibacterium epidermis DSM20659 | Optimizing the fermentation conditions and the extraction technology | monosodium glutamate | 8 | 0.05 | 0.08 e | fed-batch | [54] |
| H. boliviensis LC1T | Optimizing NaCl concentrations and the medium for fed-batch cultivations | glucose, monosodium glutamate | 4.3 | 0.07 c | 0.12 e | fed-batch | [35] |
| H. boliviensis LC1T | Optimizing the nutrient parameters in the fed-batch fermenter | glucose, monosodium glutamate | 9.2 | 0.26 e | fed-batch | [55] | |
| H. salina BCRC17875 | Optimizing the agitation speed and medium composition | yeast extract | 13.94 | fed-batch | [56] | ||
| Sinobaca sp. H24 | Isolating an ectoine producer from soil, optimizing culture medium, identifying the genes involved in ectoine biosynthesis | yeast extract, glycerol | 0.01 | flask | [57] | ||
| H. salina DSM 5928 | Optimizing the culture medium and NaCl concentration | monosodium glutamate | 6.9 | 0.33 e | batch | [58] | |
| H. elongata DSM2581 | Two nanostructures, multiwalled carbon nanotube (MWCNT) and iron oxide nanoparticle (Fe2 O3 NPs), to increase the availability of the substrate | glucose | 14.25 | batch | [59] | ||
| Marinococcus sp. MAR2 | Optimizing the culture condition with response surface methodology (RSM) and a fed-batch strategy | yeast extract | 5.6 | 0.16 e | fed-batch | [60] | |
| H. salina DSM 5928T | Optimizing the two-step fermentation conditions: growing of cells and production of ectoine by resting cells | monosodium glutamate | 14.86 | 0.14 | 0.32 e | batch | [61] |
| Marinococcus sp. ECT1 | Developing semi-synthesized medium (YAMS medium), optimizing the yeast extract concentrations | yeast extract | 2.5 | batch | [62] | ||
| H. brevibacterium sp. JCM 6894 | Optimizing the conditions for ectoine biosynthesis | polypepton, glucose, yeast extract | 2.5 | [63] | |||
| H. campaniensis G8-52 (CCTCCM2019777) | Developing a higher ectoine producer by multiple rounds of UV mutation, identifying the key mutations (orf00723 and orf02403 (lipA)) related to ectoine biosynthesis | sodium L-glutamate | 1.51 | flask | [64] | ||
| H. elongata DSM 2581T | Testing NaCl influence on ectoine biosynthesis, revealing higher NaCl concentration activating genes involved in the pentose phosphate pathway, Entner–Doudoroff pathway, flagellar assembly pathway, ectoine metabolism, repressing genes involved in the tricarboxylic acid cycle and fatty acid metabolism | 12.91 | [65] | ||||
| H. boliviensis DSM 15516(T) | Optimizing the conditions for two-step fermentation and producing ectoine with milking process | glucose | 8.9 | 0.38 e | fed-batch | [66] | |
| P. halophilum DSM 102817T | Optimizing the culture medium, developing strategies for ectoine isolation | glucose | 0.41 a | flask | [67] | ||
| H. elongate 1A01717 | Optimizing the ectoine extraction and purification process | glucose | 15.9 | fed-batch | [68] | ||
| Organism | Metabolic Engineering Strategies | Substrate | Titer (g/L) | Yield (g/g) | Productivity (g/(L.h)) | Fermentation Strategy | Reference |
|---|---|---|---|---|---|---|---|
| Engineered cell factories | |||||||
| E. coli FA08 | Optimizing the FA utilization pathway and fermentation conditions, introducing 3-HP biosynthesis module and balancing the carbon flux to maximize 3-HP production | fatty acids (FAs) | 52 | 1.56 | fed-batch | [76] | |
| E. coli ZJU-3HP01 | Developing a dual-substrate fermentative strategy, balancing the activity between glycerol dehydratase and aldehyde dehydrogenase with glucose added | glycerol and glucose | 17.20 | fed-batch | [77] | ||
| E. coli WL (pTac15kBAB, p100Rkyd) | Introducing a glycerol-dependent 3-HP biosynthetic pathway (dhaB1234, gdrAB and ydcW) from Klebsiella pneumoniae, regulating the expression of ydcW, optimizing the fed-batch fermentation conditions | glycerol | 76.2 | 0.457 | 1.89 | fed-batch | [73] |
| E. coli W DUBGK | Identifying the 3-HP-tolerant Escherichia coli strain among nine strains according to their growth in the presence of 25 g/L of 3-HP, introducing the 3-HP biosynthetic pathway into E. coli W, overexpressing them on plasmids | glycerol | 41.5 | 0.61 a | 0.86 | fed-batch | [78] |
| E. coli PSO119 | Overexpressing pyruvate aminotransferase, 3-hydroxyacid dehydrogenase, L-aspartate-1-decarboxylase, L-alanine aminotransferase, phosphoenolpyruvate carboxylase and alanine racemase, adaptive evolution, deleting L-valine transaminase, developing a dual-substrate fermentative strategy | glucose and xylose | 29.1 | 0.22 | fed-batch | [79] | |
| E. coli JHS01304 | Overexpressing galP and gpsA, analyzing the metabolome, deleting exogenous GPD1 | glucose and xylose | 37.6 | 0.17 | 0.63 | fed-batch | [80] |
| Aspergillus niger An3HP9/pyc2/ald6a∆/3HP-6 | Introducing the β-alanine biosynthetic pathway, identifying and modifying the genetic targets according to proteomic and metabolomic analysis, optimizing the fermentation conditions | corn stover hydrolysate | 36.0 | 0.48 b | [81] | ||
| E. coli C43 (DE3) ZXP05 | Developing malonic acid transporter mutants via directed evolution and enzyme-inhibition-based high throughput screening approach | malonate | 20.08 | 1.55 a | [82] | ||
| E. coli Q2186 | Directed evolution of rate-limiting enzyme MCR-C and fine tuning of MCR-N expression level, optimizing the fermentation conditions | glucose | 40.6 | 0.19 | fed-batch | [83] | |
| E. coli SH-BGK1 | Modulating the expression level of glycerol dehydratase (DhaB), alpha-ketoglutaric semialdehyde dehydrogenase (KGSADH) and glycerol dehydratase reactivase (GDR) | glycerol | 38.7 | fed-batch | [84] | ||
| E. coli JHS00947 expressing L. brevis dhaB and dhaR and E. coli aldH | Overexpression of dhaB and dhaR from Lactobacillus brevis KCTC33069 and aldH from E. coli, two-step feeding strategy | glycerol | 14.3 | 0.26 | fed-batch | [85] | |
| E. coli SH501_E209Q/E269Q | Developing variants of an aldehyde dehydrogenase (GabD4) from Cupriavidus necator | glycerol | 71.9 | 1.8 | fed-batch | [86] | |
| E. coli JHS01300/pELDRR + pCPaGGRm | Deleting ptsG, overexpressing xylR, GPD1 and GPP2 genes from S.cerevisiae, dhaB1B2B3 and dhaR1R2 from Lactobacillus brevis and aldhH from Pseudomonas aeruginosa | glucose and xylose | 29.4 | 0.36 | 0.54 | fed-batch | [87] |
| E. coli JHS_Δgypr-PT7 | Overexpressing puuC with a strong promoter, deleting puu operon repressor gene, puuR | co-fermentation of glucose and xylose | 53.7 | fed-batch | [88] | ||
| E. coli BEP113 | Overexpressing AdhEMut, mcr from Chloroflexus aurantiacus and dtsR1, accBC from Corynebacterium glutamicum, modulating pntAB expression | ethanol | 13.17 | 0.57 | [89] | ||
| S. cerevisiae ST687 | Integrating multiple copies of malonyl-CoA reductase (MCR) from Chloroflexus aurantiacus and phosphorylation- and acetyl-CoA carboxylase ACC1 genes into the chromosome, overexpressing native pyruvate decarboxylase PDC1, aldehyde dehydrogenase ALD6 and acetyl-CoA synthase from Salmonella enterica SEacs (L641P), engineering glyceraldehyde-3-phosphate dehydrogenase to increase NADPH supply, 13C metabolic flux analysis | glucose | 9.8 | 0.13 a | fed-batch | [90] | |
| S. cerevisiae N3IP_2 | Producing 3-HP in the mitochondria by overexpressing malonyl-CoA reductase (MCR) in the mitochondria, overexpressing POS5 and IDP1 to improve NADPH supply, overexpressing of an ACC1 mutant to improve 3-HP production | glucose | 71.09 | 0.23 | 0.71 | fed-batch | [74] |
| S. cerevisiae SH18 | Genome integration of MCR-C encoding C-terminal of MCR, improving supply of malonyl-CoA and NADPH by overexpressing MPCox, RtCIT1, YHM2, MmACL/AnACL, ACC1, MDH3, RtME, PYC1, IDP2, ZWF1, GND1, TKL1 and TAL1, modulating the expression of a fatty acid synthase gene FAS1 with a glucose concentration-sensitive promoter PHXT1 | glucose | 56.5 | 0.31 | 0.53 | fed-batch | [69] |
| Pichia pastoris PpHP6 | Introducing and engineering the mcr gene from Chloroflexus aurantiacus, improving NADPH and malonyl-CoA supply by overexpressing the ACCYl and cPOS5Sc, optimizing the fermentation conditions | glycerol | 24.75 | 0.13 | 0.54 | fed-batch | [91] |
| K. pneumoniae with YneI overexpression | Overexpressing aldehyde dehydrogenase, YneI and YdcW | glycerol | 2.4 | shake-flask culture | [92] | ||
| K. pneumoniae with aldehyde dehydrogenases (ALDH) from Bacillus subtilis | Introducing aldehyde dehydrogenases (ALDH), DhaS from B.subtilis | glycerol | 18 | [93] | |||
| K. pneumoniae Q1643 | Overexpressing glycerol dehydratase, its reactivation factor (dhaB123, gdrA and gdrB from K. pneumoniae), aldehyde dehydrogenase (aldH from E. coli), deleting dhaT and yqhD | glycerol | 2.03 | flask culture | [94] | ||
| K. pneumoniae ΔadhPΔpflB (pTAC-puuC) | Deleting adhP and pflB, overexpressing puuC | glycerol | 66.91 | 1.40 | fed-batch | [95] | |
| K. pneumoniae -T7 (pET28a-puuC) | Developing the T7 expression system and overexpressing puuC | glycerol | 67.59 | 0.5632 | fed-batch | [96] | |
| K. pneumoniae | Overexpressing ald4 and dhaB, optimizing the fermentation conditions | glycerol and glucose | 3.77 | flask | [97] | ||
| K. pneumoniae with L. reuteri pduP overexpression | Overexpressing a pduP gene from Lactobacillus reuteri | glycerol | 1.38 | batch fermentation | [98] | ||
| K. pneumoniae ΔdhaTΔyqhD overexpressing both PuuC and DhaB | Deleting dhaT and yqhD, overexpressing puuC and dhaB | glycerol | >28 | >0.4 | fed-batch | [99] | |
| K. pneumoniae (p3tac-PuuC) | Overexpressing puuC, optimizing fermentation conditions, mathematical model analysis | glycerol | 102.61 | fed-batch | [72] | ||
| K. pneumoniae Δldh1Δldh2Δpta (pTAC-puuC) | Overexpressing puuC, deleting the pathways for lactate and acetate biosynthesis according to metabolix flux analysis, optimizing fermentation conditions, describing a flux distribution model of glycerol metabolism | glycerol | 83.8 | 0.54 | fed-batch | [100] | |
| Schizosaccharomyces pombe overexpressing Cut6p and CaMCR | Overexpressing the S. pombe acetyl-CoA carboxylase (Cut6p) and the malonyl-CoA reductase from Chloroflexus aurantiacus (CaMCR) with the S. pombe hsp9 promoter, optimizing the fermentation conditions | glycerol and acetate | 7.6 | [101] | |||
| Schizosaccharomyces pombe | Dissecting the mcr gene from Chloroflexus aurantiacus into two functionally distinct fragments and balancing the activity of them, overexpressing aldehyde dehydrogenase, acetyl-CoA synthase and pantothenate kinase, introducing beta-glucosidase | glucose and cellobiose | 11.4 | 0.11 b | fed-batch | [102] | |
| engineered Halomonas bluephagenesis TD27 | Deleting the 3-HP degradation pathway, overexpressing alcohol dehydrogenases (AdhP) | 1,3-propanediol | 154 | 0.93 | 2.4 | fed-batch | [75] |
| Natural producers | |||||||
| Lactobacillus reuteri | Optimizing fermentation conditions | glycerol | 5.2 | 1.3 | fed-batch | [103] | |
| Lactobacillus reuteri DSM17938 | Comparing the ability of three Lactobacillus reuteri strains to produce 3-HP, analyzing the influence of glycerol and metabolites on strains’ physiological states and survival | glycerol | 2 | [104] | |||
| Debaryomyces hansenii WT39 | Selecting strains with propionic acid as the substrate, making mutations with the low-energy ion N+ | glucose | 62.42 | 1.30 | flask | [105] | |
| Rhodococcus erythropolis LG12 | Isolating strains with acrylic acid as the substrate, optimizing the fermentation conditions | acrylic acid | 17.5 | 1.11 a | 0.22 | [106] | |
| Lentilactobacillus diolivorans | Optimizing the fermentation conditions | 0.025 mol/mol glucose/glycerol | 67.7 | fed-batch | [107] | ||
| Gluconobacter oxydans ZJB09112 | Optimizing the fermentation conditions | 1,3-propanediol | 76.3 | 1.5 | fed-batch | [108] | |
| K. pneumoniae and Gluconobacter oxydans | Developing a two-step process to produce 3-HP with glycerol | glycerol | 60.5 | 0.50 | [109] | ||
| Organism | Metabolic Engineering Strategies | Substrate | Titer (g/L) | Yield (g/g) | Productivity (g/(L.h)) | Fermentation Strategy | Reference |
|---|---|---|---|---|---|---|---|
| Engineered cell factories | |||||||
| E. coli DPAL 8 | Overexpressing pck, maeB, ilvD, ilvBN and cycA, decreasing the expression of gdhA, deleting pta, optimizing the fermentation conditions | glucose β-alanine | 66.39 | 0.27 b | fed-batch | [116] | |
| E. coli DPA02/pT-ppnk | Overexpressing ppnk, deleting aceF and mdh, optimizing the fermentation conditions | glucose | 68.3 | 0.36 | 0.794 | fed-batch | [118] |
| E. coli BL21(DE3) strain expressing pantothenate synthetase from C.glutamicum | Overexpressing pantothenate synthetases from C. glutamicum | pantoate and β-alanine | 97.1 | 3.0 | substrate added at the beginning | [113] | |
| E. coli W3110 DPA-11/pTrc99A-panB-K25A/E189S-panC | Protein engineering of ketopantoate hydroxymethyltransferase from C. glutamicum, overexpressing panB, CgKPHMT-K25A/E189S and panC | glucose β-alanine L-isoleucine | 41.17 | 0.65 | fed-batch | [119] | |
| E. coli W3110/pTrc99A-panB-panC | Optimizing the fermentation conditions, overexpressing panB and panC | L-isoleucine glucose | 31.6 | 0.17 a | 0.55 a | fed-batch | [120] |
| E. coli DPA-9/pTrc99a-panBC(C.G) | Overexpressing panB, panC, panE and ilvC, making mutations of ilvG and coaA, deleting avtA and ilvA, deregulating ilvE | β-alanine glucose, | 28.45 | 0.40 | fed-batch | [121] | |
| Escherichia DPA21 | Decreasing ilvE expression, overexpressing ilvBN, glyA, pntAB, cyo, cyoA and serAfbr, optimizing the fermentation conditions according to comparative transcriptome and metabolomics analysis | citric acid, glucose, β-alanine | 45.35 | 0.31 | 0.50 b | fed-batch | [122] |
| Bacillus megaterium BM-4 | Overexpressing panBC, panE, ilvBNC, ilvD, serA and glyA | glucose, β-alanine | 19.70 | 0.26 | 0.78 b | fed-batch | [114] |
| B. megaterium BM-1 (pantoate-β-alanine ligase (PBL)) | Overexpressing panC from B. subtilis | pantoate and β-alanine | 45.56 | fed-batch | [115] | ||
| C. glutamicum Pan-4/pXtuf-panBCDBsu | Overexpressing panBCD from B. subtilis, ilvBNC, aspB and aspA, deleting avtA, ilvE and ilvA | glucose | 18.62 | 5 L bioreactor | [123] | ||
| Saccharomyces cerevisiae DPA171 | Enhancing the D-pantothenic acid biosynthetic pathway by adjusting the copy numbers of key genes, deleting bypass genes, balancing cofactor utilization, optimizing GAL-inducible system | glucose | 4.1 | [124] | |||
| Natural producers | |||||||
| E. coli ECPA | Optimizing the fermentation conditions | L-isoleucine glucose | 39.1 | 0.175 | 0.58 a | fed-batch | [117] |
| Organism | Metabolic Engineering Strategies | Substrate | Titer (g/L) | Yield (g/g) | Productivity (g/(L.h)) | Fermentation Strategy | Reference |
|---|---|---|---|---|---|---|---|
| E. coli W-18/pM2/pR1 | Overexpressing glf, ppc, aspA, glk, asd, metL and rhtA, deleting lysA, thrB, metA, lacI, ldhA, adhE, pflB, ptsG, iclR and arcA, optimizing the fermentation conditions | glucose | 110.8 | 0.64 | 1.82 | fed-batch | [126] |
| E. coli HOM-14 | Overexpressing thrB, thrAfbr, ppc, aspA8, pntAB and rhtA | glucose | 60.1 | 0.42 | 1.25 | fed-batch | [130] |
| E. coli H28 | Overexpressing thrAfbr, thrABC, ppc, aspC, aspA lysCcglfbr, rhtA pntAB, asdtmo and adhpae, deleting lacI | glucose | 85.29 | 0.43 | 1.78 | fed-batch | [131] |
| E. coli LJL12 | Overexpressing thrA, deleting lysA, metA, thrBC, iclR, gltA, pykA and pykF | glucose | 35.8 | 0.35 | 0.82 | fed-batch | [132] |
| E. coli SHL17 | Overexpressing pntAB, rhtA, ppc, thrA and asd, introducing a hok/sok toxin/antitoxin system | glucose | 44.4 | 0.21 | 0.93 | fed-batch | [133] |
| E. coli HS15 | Overexpressing pntAB, rhtB, glk, zglf, ppc, aspC, gdhA, thrA, asd and aspA, deleting lysA, metA, thrB, lacI, ldhA, poxB, pflB and iclR | glucose | 84.1 | 0.50 | 1.96 | fed-batch | [128] |
| C. glutamicum Cg18-1 | Overexpressing thrAS345F, aspC, pycP458S, lysCT311I, asd, homV59A, brnFE, icdM1V, dapAM1V and gapN, deleting mcbR, metD, thrB, NCgl2688 and metY | glucose | 63.5 | 0.25 | fed-batch | [129] | |
| C. glutamicum Cg09−1 | Overexpressing lysC, asd, hom, pyc, brnFE, lysCT311I and asd from C. glutamicum and aspC and thrAS345F from E. coli, deleting thrB, mcbR and metD, decreasing the expression of dapA and icd | glucose | 8.8 | batch | [134] | ||
| Corynebacterium sp. 9366-EMS/329 | Developing a mutant Corynebacterium sp. requiring threonine | sucrose | 14.5 | batch | [135] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, A.; Liu, Y.; Jia, B.; Zheng, G.; Yao, Y. Metabolic Engineering of Microorganisms to Produce L-Aspartate and Its Derivatives. Fermentation 2023, 9, 737. https://doi.org/10.3390/fermentation9080737
Shi A, Liu Y, Jia B, Zheng G, Yao Y. Metabolic Engineering of Microorganisms to Produce L-Aspartate and Its Derivatives. Fermentation. 2023; 9(8):737. https://doi.org/10.3390/fermentation9080737
Chicago/Turabian StyleShi, Aiqin, Yan Liu, Baolei Jia, Gang Zheng, and Yanlai Yao. 2023. "Metabolic Engineering of Microorganisms to Produce L-Aspartate and Its Derivatives" Fermentation 9, no. 8: 737. https://doi.org/10.3390/fermentation9080737
APA StyleShi, A., Liu, Y., Jia, B., Zheng, G., & Yao, Y. (2023). Metabolic Engineering of Microorganisms to Produce L-Aspartate and Its Derivatives. Fermentation, 9(8), 737. https://doi.org/10.3390/fermentation9080737